
藜麦愈伤组织诱导和分化条件初探
2021-11-24 10:13王燕芳宋时奎
亚热带农业研究 2021年2期
王燕芳, 宋时奎
(1.福建农林大学园艺学院;2.福建农林大学园艺植物生物学及代谢组学研究中心,福建 福州 350002)
藜麦(ChenopodiumquinoaWilld.) 又称藜谷、南美藜和奎奴亚藜,原产于南美洲中高海拔山区,为藜科藜属1年生双子叶植物[1-2],是一种全营养型谷物。藜麦耐盐碱、干旱、低温和贫瘠等[3-5],能够在盐碱地上种植,某些基因型甚至可以在pH>8或盐度高于海水(500 mmol·L-1NaCl)的土地上生存[6-7]。
由于藜麦花小,杂交操作困难,传统育种和基因功能研究受到了限制。以转基因技术为代表的分子育种手段可以打破种间限制而缩短育种年限,同时便于研究基因功能[8]。根癌农杆菌介导的遗传转化是转基因技术中应用最为普遍的一种,且植物遗传转化大多需要经历再生过程[9]。有关藜麦再生成功的报道较少。2005年,Eisa et al[10]以藜麦下胚轴为外植体诱导愈伤组织,首次实现了体细胞胚再生;2016年,Hesami et al[11]以藜麦下胚轴为外植体诱导愈伤组织,并分化出丛生芽;2018年,曹宁等[12]用台湾红藜茎段为外植体诱导愈伤组织进而建立了再生体系;2020年,段鹏慧等[13]用带腋芽的茎段为外植体,经过腋芽诱导、不定芽增殖和生根培养实现了藜麦的再生。藜麦再生的后续报道较少且都未涉及遗传转化,至今尚未见藜麦转基因成功的报道,限制了藜麦分子育种的发展。因此,本研究对藜麦愈伤组织诱导条件进行优化,并尝试建立再生体系,以期为其遗传转化提供参考。
1 材料与方法
1.1 供试材料
1.1.1 植物材料 选用福建农林大学园艺植物生物学及代谢组学研究中心保存的藜麦品种为材料。藜麦种子用氯气灭菌6 h,再将灭菌的种子点种于含1/2MS培养基的组织培养瓶中,培养5 d后获得藜麦无菌苗。
1.1.2 菌株和载体 双元载体PTF101S分别携带目的基因Wusa、CLV3、BBM1和SERK;PTF102携带目的基因GUS和BAR。菌株为根癌农杆菌EHA105。
1.2 愈伤组织诱导
参考文献[10-12],选用藜麦无菌苗下胚轴为外植体进行农杆菌侵染并诱导愈伤组织。将无菌苗的下胚轴切成0.5~1.0 cm的小段,放入根癌农杆菌菌液中侵染20 min。根癌农杆菌菌液制备如下:将菌株EHA105涂布于相应抗性YEP培养基上进行初次活化,在28 ℃根癌农杆菌恒温培养箱中倒置培养48 h,之后进行二次活化;1 d后收集菌体,悬浮于MS侵染液中,菌液浓度为△D600 nm=0.3。侵染结束后,将下胚轴移至共培养培养基中暗培养3 d,再移至诱导培养基培养4周。诱导培养基为MSB培养基(购于Phyto Tech公司),激素为0.5 mg 2,4-D和0.1 mg KT;凝固剂为2.5 g结冷胶。在诱导过程中,分别比较藜麦品种、光照、碳源、外源基因对愈伤组织诱导的影响。
1.2.1 品种 为了比较不同藜麦品种愈伤组织诱导的难易程度,分别对18个品种的下胚轴进行诱导,其他条件均一致。参试品种分别为:带壳红藜5号、高雄红藜9号、山西白藜(购于山西亿隆)、SKU18339、SKU176771、MS509/N、QN108B、1ESP、141R、144R、11P、COPACABANA、Villa Juira、Rosa Junin、CQ119、CQ135、CJAPANESE STRAIN和NL-6。
1.2.2 光照 光作为一种环境信号而影响植物组织的生长发育。根据1.2.1品种比较的结果,从易诱导品种中随机挑选带壳红藜5号、高雄红藜9号和山西白藜,将其下胚轴分别用EHA105/PTF102、EHA105/PTF101S进行侵染。 侵染结束后分别在光强为2 000 lx、光周期为16 h光照/8 h黑暗条件下进行培养。4周后观察愈伤组织形态。
1.2.3 培养基碳源 将MS509/N下胚轴分别用携带GUS、SERK、Wusa、BBM1、CLV3基因的EHA105菌株及含有SERK、Wusa、BBM1和CLV3的混合菌液进行侵染。所用菌液制备方法同1.2。共培养结束后分别移至碳源为葡萄糖、麦芽糖、蔗糖的诱导培养基中培养,3~4周后比较3种碳源的效果。选出诱导效果较好的葡萄糖和麦芽糖,进一步比较二者的诱导效果。将MS509/N下胚轴分别移入碳源为30 g葡萄糖、30 g麦芽糖、20 g葡萄糖+10 g麦芽糖、20 g麦芽糖+10 g葡萄糖的诱导培养基中,3周后观察效果。
1.2.4 外源基因SERK、CLV3、Wusa与植物茎顶端干细胞的发育相关[14-16]。带壳红藜5号和山西白藜下胚轴分别用携带不同外源基因(SERK、CLV3、Wusa)的农杆菌EHA105侵染,以不携带外源基因的EHA105作为对照,4周后比较愈伤组织的分化情况。
1.3 愈伤组织分化
挑选形态大小、质地一致的愈伤组织进行分化培养,探索KNO3和6-BA浓度对愈伤组织分化的影响。分化培养基的大量元素、微量元素和铁盐用量与MS培养基相同;有机元素种类与B5培养基相同,但肌醇用量增加1倍、VB1用量增加10倍;激素为6-BA;凝固剂为2.5 g植物凝胶。分化培养条件为光照3 000 lx,光周期为16 h光照/8 h黑暗,温度(25±1) ℃。
1.3.1 KNO3质量浓度 硝态氮是植物吸收利用的主要形式。愈伤组织分别在950、1 900和3 800 mg·L-1KNO3的培养基中分化4周,比较其形态。
1.3.2 6-BA质量浓度 6-BA是最常用的细胞分裂素。愈伤组织在不含6-BA和添加0.35、1.00 mg·L-16-BA的培养基中分化4、8周,观察分化结果。
2 结果与分析
2.1 品种、光照等对藜麦愈伤组织诱导的影响
2.1.1 品种 如图1所示, SKU176771、SKU18339、MS509/N、QN108B、CJAPANESE STRAIN、NL-6、带壳红藜5号、高雄红藜9号和山西白藜共9个品种诱导出形态较好的愈伤组织;Villa Juira、1ESP、141R和144R的下胚轴前期能够分裂,后期停止;COPACABANA、Rosa Junin和11P诱导的愈伤组织体积小易褐化,且后期不再增大;CQ119和CQ135的下胚轴逐渐干枯死亡,不能被诱导。由上可见,不同品种自身敏感性有很大区别。

A.SKU176771;B.SKU18339;C.MS509/N;D.QN108B;E.CJAPANESE STRAIN;F.NL-6;G.带壳红藜5号;H.高雄红藜9号;I.山西白藜;J.Villa Juira(同1ESP、141R、144R);K.Rosa Junin(同COPACABANA、11P);L.CQ119(同CQ135)。图1 不同藜麦品种愈伤组织诱导难易的比较Figure 1 Comparison of callus induced from different quinoa cultivars
2.1.2 光照 植物组织形态的建成、器官分化和生长调控都需要光的参与,尤其外植体细胞最初的分裂对光的依赖更大[17]。如图2所示,带壳红藜5号、高雄红藜9号和山西白藜的下胚轴在光照下诱导的愈伤组织体积较大。同时,红藜的愈伤组织呈现红色,白藜呈现绿色或红色,这可能是由于甜菜红素积累引起的。 暗处下,带壳红藜5号和高雄红藜9号的愈伤组织形成速度较慢且为浅白色,易生成根毛,而山西白藜的下胚轴几乎不能被诱导成愈伤组织。由上可见,光照有利于藜麦愈伤组织的诱导。
2.1.3 碳源 碳源种类不仅会影响细胞的生长速度,还会影响代谢水平。本研究中,单独以蔗糖为碳源的愈伤较硬、容易破碎且玻璃化严重,以葡萄糖和麦芽糖为碳源的愈伤没有明显的区别,说明蔗糖不适合作为碳源诱导藜麦愈伤组织(图3)。进一步比较葡萄糖和麦芽糖对愈伤组织诱导的影响表明,以30 g葡萄糖为碳源,愈伤组织易水化;以30 g麦芽糖为碳源,诱导速度较慢;以(20 g麦芽糖+10 g葡萄糖)为碳源较以(20 g葡萄糖+10 g麦芽糖)为碳源的诱导效果更好,愈伤组织形态质地最佳(图4)。

第1排为光照下诱导,第2排为暗处诱导;A、G:带壳红藜5号转化PTF102;B、H:带壳红藜5号转化PTF101S;C、 I:高雄红藜9号转化PTF102;D、J:高雄红藜9号转化PTF101S; E、K:山西白藜转化PTF102;F、L:山西白藜转化PTF101S。图2 光照和暗处理对藜麦愈伤组织诱导的影响Figure 2 Effects of lighting and darkness on quinoa callus redifferentiation

A.葡萄糖,B.蔗糖,C.麦芽糖;1~6表示转化的外源基因分别为:CUS、SERK、Wusa、BBM1、CLV3、SERK+Wusa+BBM1+CLV3。图3 3种碳源对藜麦愈伤组织诱导效果的比较Figure 3 Comparisons of quinoa callus induced under 3 carbon sources

A.30 g葡萄糖;B.20 g葡萄糖+10 g麦芽糖;C.20 g麦芽糖+10 g葡萄糖;D.30 g麦芽糖。图4 麦芽糖和葡萄糖比例对藜麦愈伤组织诱导的影响Figure 4 Effects of different combinations of maltose and glucose on quinoa callus redifferentiation
2.1.4 外源基因SERK是编码体细胞胚发生相关类受体蛋白激酶的一个重要基因[18]。如图5所示,转化SERK的下胚轴形成愈伤组织的速度较快,愈伤组织体积比对照、过表达CLV3和Wusa的愈伤组织大。其中,红藜形成的愈伤组织比山西白藜愈伤组织整体更容易褐化。
2.2 KNO3和6-BA对藜麦愈伤组织分化的影响

A.带壳红藜5号,B.山西白藜;1~4愈伤组织过表达外源基因: CK、SERK、CLV3和Wusa。图5 外源基因对藜麦愈伤组织诱导的影响Figure 5 Effect of exogenous gene on quinoa callus redifferentiation
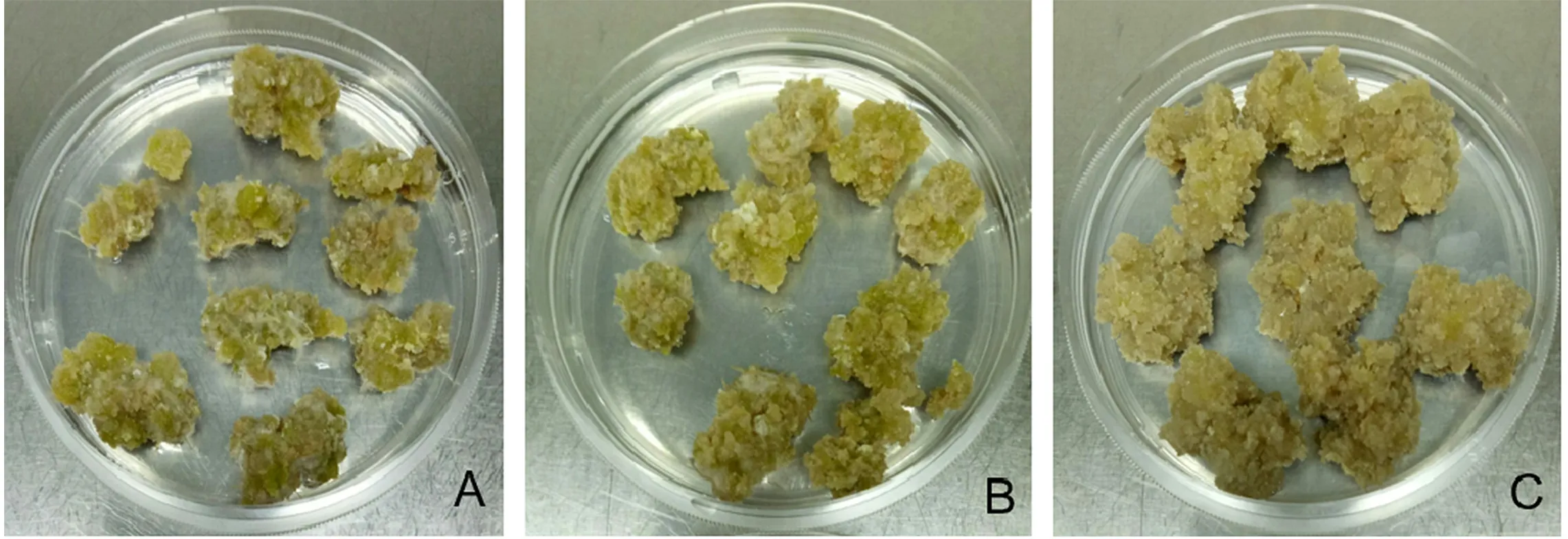
A.950 mg·L-1;B.1 900 mg·L-1;C.3 800 mg·L-1。图6 KNO3浓度对藜麦愈伤组织分化的影响Figure 6 Effect of KNO3 concentration on quinoa callus redifferentiation
2.2.2 6-BA 分化4周时,在未添加6-BA的培养基中藜麦愈伤组织长出大量小而密的根毛,愈伤组织较硬;在0.35 mg·L-16-BA中,愈伤组织表面有轻微玻璃化现象,但较为新鲜,质地疏松;在1.00 mg·L-16-BA中,玻璃化严重且伴随褐化现象,愈伤块易碎。分化8周时,在未添加6-BA的培养基中愈伤组织根毛伸长,1 mg·L-16-BA较0.35 mg·L-16-BA愈伤组织玻璃化严重(图7)。因此,在再生过程中不添加6-BA或浓度过高(1.00 mg·L-16-BA)均不利于愈伤组织分化。

A、B、C分化4周,D、E、F分化8周;A、D为未添加6-BA;B、E为添加0.35 mg·L-1 6-BA;C、F为添加1.00 mg·L-1 6-BA。图7 6-BA浓度对藜麦愈伤组织分化的影响Figure 7 Effect of 6-BA concentration on quinoa callus redifferentiatio
3 讨论与结论
本研究表明,品种、碳源、光照和外源基因对藜麦愈伤组织诱导有不同程度的影响,其中碳源和光照影响较大。葡萄糖单独诱导愈伤组织生长较快,但易水化,可能由于葡萄糖易被细胞吸收引起快速生长从而导致水化现象,且不利于维持培养基渗透势稳定[21]。麦芽糖能够分解为葡萄糖且分解速度较慢, 两种糖组合既保证了愈伤诱导速度也维持了渗透压。因此,以(20 g麦芽糖+10 g葡萄糖)为碳源最适合藜麦愈伤组织诱导。适量的光照较黑暗有利于藜麦愈伤组织诱导,黑暗下愈伤组织易诱发根毛的生长,这一结果与孙阳等[17]和李占歧等[22]对棉花愈伤组织的培养结果相似,但与玉米[23]、水稻[24]和半枝莲[25]等植物的愈伤组织诱导条件不同。
在藜麦愈伤组织分化培养中,KNO3和6-BA浓度对愈伤组织分化的速度和形态有显著影响。参考棉花下胚轴愈伤组织诱导中硝态氮和铵盐的用量[26],在藜麦愈伤组织分化过程中, 950 、1 900 mg·L-1KNO3效果没有较大区别,而3 800 mg·L-1则会导致愈伤组织严重玻璃化。由于藜麦愈伤组织并未成功分化出芽,因此本研究只确定KNO3在分化培养基中浓度不宜过高。分化培养基不含6-BA时,藜麦愈伤组织易长出细小根毛,可能是因为缺少细胞分裂素而使生长素相对含量高;6-BA为0.35 mg·L-1时,愈伤组织形态相对较好,但同样未能再生成功;6-BA浓度过高(1.00 mg·L-1),愈伤组织太过疏松且易出现褐化和玻璃化,这可能由于6-BA能促进多酚氧化酶的活性从而加速褐化[27]。因此,藜麦愈伤组织分化培养时,不含6-BA或其浓度过高均不适合。
藜麦愈伤组织分化成功的报道不多,至今尚未见转基因藜麦成功的报道。因此,本研究选用转基因方法中较为常用的根癌农杆菌介导法,在农杆菌侵染的基础上优化了藜麦愈伤组织诱导条件,摸索影响愈伤组织再生的因素,以期为其遗传转化体系的建立提供参考。
猜你喜欢
油气田地面工程(2022年9期)2022-10-26
作物学报(2022年9期)2022-07-18
舰船科学技术(2022年11期)2022-07-15
保健与生活(2022年13期)2022-07-06
江苏农业科学(2022年6期)2022-04-15
草业科学(2022年3期)2022-03-26
铀矿冶(2022年1期)2022-02-17
阅读(低年级)(2019年11期)2019-12-27
作文与考试·小学高年级版(2017年24期)2017-12-14
老友(2016年3期)2016-05-26
