
红条毛肤石鳖和日本花棘石鳖的转录组比较分析
2021-11-19 04:44:14
闽江学院学报 2021年5期
(闽江学院海洋研究院,福建 福州 350108)
0 引言
多板纲(Polyplacophora)属于软体动物门,本纲动物统称石鳖,是广泛存在于潮间带的海洋动物,从热带到极地的世界各大洋中均有分布,通常在岩石上营底栖附着生活,对潮间带的生态平衡和能量流动起着重要作用[1]。石鳖背部通常有八片连续的壳板,壳板被文石鳞片或针状体环带包围,俗称“八节毛”“石铁板”“铁角”等。我国东南沿海石鳖种类较多,例如红条毛肤石鳖(Acanthochitonarubrolineata)、日本花棘石鳖(Liolophuurajaponica)、日本宽板石鳖(Placiphorellajaponica)和平濑锦石鳖(Onithochitonhirasei)等[2],其中红条毛肤石鳖属于毛肤石鳖科,属于广温广布性种类,数量较多,也有研究表明该物种对医治淋巴结核和麻风病有一定的疗效;日本花棘石鳖属于石鳖科,是亚热带、热带暖水性种类,在福建分布较为广泛,而且日本花棘石鳖个体相对更大,常被沿海居民采捕食用。此外,石鳖的齿舌因结构和磁性的特殊性受到广泛关注,其磁性物质为纳米颗粒结构,是有潜力的未来磁性纳米材料来源之一[3]。
在软体动物门中,多板纲独特的壳板使其具有特殊的形态结构。研究表明Pax(Paired-box)基因家族与动物发育形态密切有关,编码的蛋白是一组极为重要的转录调控因子,在胚胎发育的器官形成中扮演重要角色。Pax基因家族的主要特征是编码含有128个氨基酸组成的成对域(paired domain,PD)[4],Pax蛋白通过成对域与DNA或其他蛋白相互作用,其主要功能包括:调控细胞增殖、促进细胞自我更新、诱导前体细胞定向转移以及改变特异细胞系的分化方向等[5]。研究表明Pax因子的非正常表达会导致多种器官组织发育畸形[6]。根据结构域组成的差异将其分为不同的亚族:Pax1/9、Pax2/5/8、Pax3/7、Pax4/6、Poxneuro、Paxα/β和Paxeyg[7]。多板纲形态独特,但目前软体动物中关于Pax基因家族的研究较少,在多板纲动物中的研究几乎为空白。
石鳖除了具有特殊的形态结构之外,在研究软体动物的起源与进化中也有着重要的地位,是软体动物门中的原始类群[8]。目前,关于多板纲动物的分子数据较少,仅有少量线粒体基因组及线粒体细胞色素c氧化酶亚基Ⅰ基因(COⅠ)数据[9-11],对其分类仍然主要依赖形态观察,包括壳板、齿舌、外套膜的形态差异[12]。目前,通过分子生物学技术对多板纲进行进化分析是发展趋势。丰富多板纲的分子数据有利于明确多板纲分类,加深对其进化地位的了解。因此,本文采用二代Illumina Hi-seq测序技术对红条毛肤石鳖和日本花棘石鳖两种石鳖种类进行了转录组测序,构建转录组数据库,并对Unigene进行功能注释,丰富基因数据库,为以后明确多板纲动物分类积累数据,为解析多板纲进化地位奠定基础。
1 材料与方法
1.1 样品收集和总RNA提取
在福建省厦门市浅滩采集两种石鳖,依据形态特征和18S RNA鉴定物种分别是红条毛肤石鳖和日本花棘石鳖[12],将整个个体组织迅速放入液氮中速冻,随后放在-80℃冰箱中保存。每个物种选取3个个体,将其混合在一起组成一个转录组测序样品,通过TRIzol法提取RNA,琼脂糖凝胶电泳和Nanodrop分析样品RNA浓度和完整性,用于转录组测序。
1.2 cDNA文库建库和转录组测序
石鳖的转录组建库、测序和拼接组装委托北京诺禾致源生物信息科技有限公司完成。建库使用的试剂盒为Illumina的NEBNext© UltraTM RNA Library Prep Kit,以mRNA为模板,构建cDNA双链,库检合格后,通过Illumina Hi-seq进行测序。为了保证数据分析的质量及可靠性,对原始数据进行过滤,去除带接头、含N、低质量reads,同时对Clean data进行Q20、Q30和GC含量计算,后续所有分析均是基于Clean data。
1.3 基因注释和基因富集分析
鉴于石鳖无参考基因组,采用Trinity对Clean reads进行拼接,将拼接得到的转录本序列作为后续分析的参考序列,取每条基因中最长的转录本作为Unigene,进行后续的分析。对转录本及Unigene的长度分别进行统计。为获得全面的基因功能信息,进行了7大数据库的基因功能注释,包括:Nr、Nt、Pfam、KOG/COG、Swiss-prot、KEGG和GO。对基因进行GO注释之后,将注释成功的基因按照GO的3个大类(生物学过程,细胞组分,分子功能)的下一层级进行分类。
1.4 SSR分析
简单重复序列标记(simple sequence repeats,SSR)的搜索与筛选采用MISA软件,对应的各个unit size(1-6)的最少重复次数分别为10、6、5、5、5、5,例如以二碱基重复单位计,最少重复数为6。对Unigene进行SSR检测。根据SSR两端序列,采用Primer 3进行SSR引物设计。
1.5 Pax基因家族分析
通过对石鳖转录组进行序列分析,得到部分或完整的Pax基因序列。通过预测蛋白结构域(http://smart.embl.de/),选择具有Pax结构域的基因,同时下载已公布的软体动物Pax序列(https://www.ncbi.nlm.nih.gov/gene),用MEGA 7.0自带的ClustalW程序进行氨基酸的多序列比对,将两端进行人工修齐后,采用NJ邻接法构建系统发生树,同时采用1 000次bootstrap进行检验。将系统发生树导入iTOL网站(http://itol.embl.de/)进行图片处理。
2 实验结果
2.1 石鳖转录组数据质量和基因统计
经过原始数据过滤、测序错误率检查、GC含量分布检查等,获得后续分析使用的Clean reads,数据汇总如表1所示。红条毛肤石鳖样品(AP)测序后得到60 666 664条原始reads数,数据过滤后得到50 109 686条reads数,Q20值和Q30值为96.20%和90.35%,GC含量为47.41%。日本花棘石鳖样品(LJ)测序后得到48 856 746条原始reads数,过滤后得到48 052 466条reads数,Q20值和Q30值为97.08%和92.25%,GC含量为45.13%。

表1 石鳖的转录组测序基本信息
组装获得Unigene后,将所有Unigene在NR、NT、KO、SwissProt、PFAM、GO和KOG 7大数据库进行蛋白功能基本注释,注释结果见表2。红条毛肤石鳖和日本花棘石鳖转录组共有121 892和83 776个Unigene。NR注释到的Unigene所占比例最高,分别有30 299和22 827个Unigene被注释;在NT数据库中注释的Unigene所占比例最低,分别是5.03%和5.96%;综合7个数据库的结果,分别有3 111个和2 682个Unigene在7个数据库中均被注释;39 908和29 528个Unigene至少在其中一个数据库中有被注释。

表2 在数据库中基因注释数目与注释成功率
2.2 基因注释和数据基本分析
GO和KEGG富集分析发现,两个石鳖转录组表达的基因富集的结果无显著差异。富集最高的5个GO条目均是“生物学过程”的“Cellular process”“Metabolic process”“Single-organism process”和“分子功能”的“Binding”和“Catalytic activity”(图1)。在KEGG分类中,富集到基因数目最多的5个类群依次是“Signal transduction”“Transport and catabolism”“Endocrine system”“Folding, sorting and degradation”和“Translation”(图2)。
对红条毛肤石鳖和日本花棘石鳖转录组的Unigene进行SSR检测,分别筛选到14 602个和15 531个SSR,单碱基重复单位SSR的重复次数主要集中在9~12次,其次是13~16次,2~5碱基重复单位SSR的重复次数主要集中5~8次(图3)。
2.3 Pax分析
在红条毛肤石鳖转录本中鉴定到5条Pax基因,包含3条Pax2/5/8,1条Pax1/9和1条Pax6,在日本花棘石鳖转录本中鉴定到3条Pax基因,分别是Pax1、Pax2/5/8和Pax2(表3)。根据已发表的软体动物Pax序列,整理归纳的Pax2/5/8基因及其剪切体有38条,Pax3/7基因及其剪切体有12条,Pax1/9基因及其剪切体有8条,Pax-α/β基因及其剪切体有2条,Pax6基因及其剪切体有20条(表4)。将以上公布的80条Pax基因和来自两个石鳖转录组的8条Pax序列一起构建系统发生树,结果显示软体动物Pax基因家族明显聚为5类(图4):Pax1/9、Pax2/5/8、Pax3/7、Pax-α/β和Pax6,石鳖的Pax基因与相同的基因亚族聚为一支,具有较高的同源性。
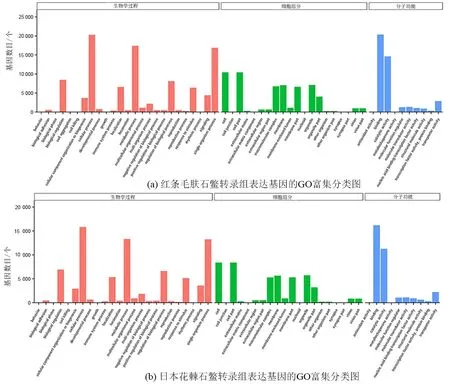
注:横坐标为GO3个大类的下一层级的GO term,纵坐标为注释term下的基因个数。 Biological Pathway:生物学过程;Molecular Function:分子功能;Cellular Component:细胞组分。图1 红条毛肤石鳖和日本花棘石鳖转录组表达基因的GO富集分类图Fig.1 The enriched GO terms of expression genes in the transcriptome of A. rubrolineata and L. japonica
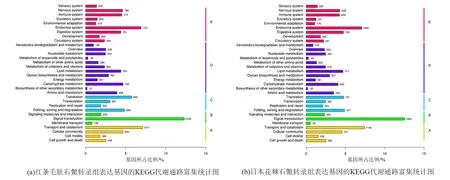
注:图中纵坐标为KEGG代谢通路的名称,横坐标为注释到该通路下的基因个数及其个数占被注释上的基因总数的比例。将基因根据参与的KEGG代谢通路分为5个分支:细胞过程(A,Cellular Processes),环境信息处理(B,Environmental Information Processing),遗传信息处理(C,Genetic Information Processing),代谢(D,Metabolism),有机系统(E,Organismal Systems)。图2 红条毛肤石鳖和日本花棘石鳖转录组表达基因的KEGG代谢通路富集统计图Fig.2 Diagram of KEGG metabolic pathway enrichment of expression genes in the transcriptome of A. rubrolineata and L. japonica
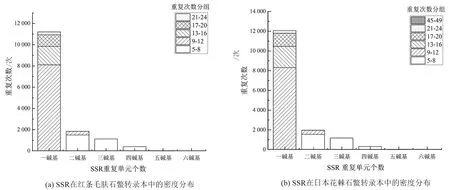
图3 SSR在红条毛肤石鳖(a)和日本花棘石鳖(b)转录本中的密度分布Fig.3 Distribution of SSR in the transcriptome of A. rubrolineata(a) and L. japonica(b)

表3 红条毛肤石鳖和日本花棘石鳖转录本中鉴定的Pax基因
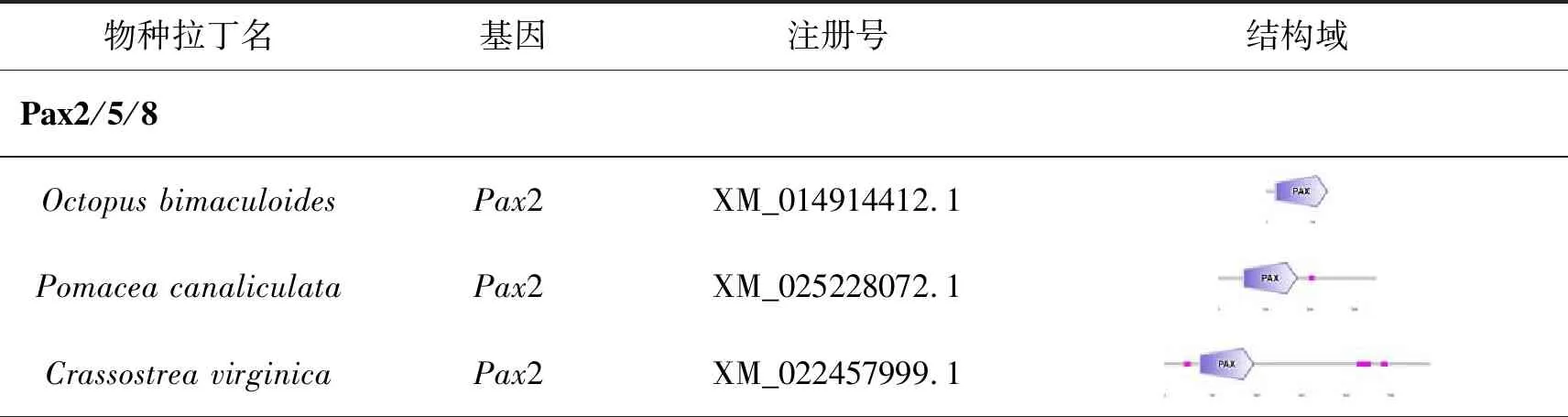
表4 软体动物的Pax蛋白结构域

续表
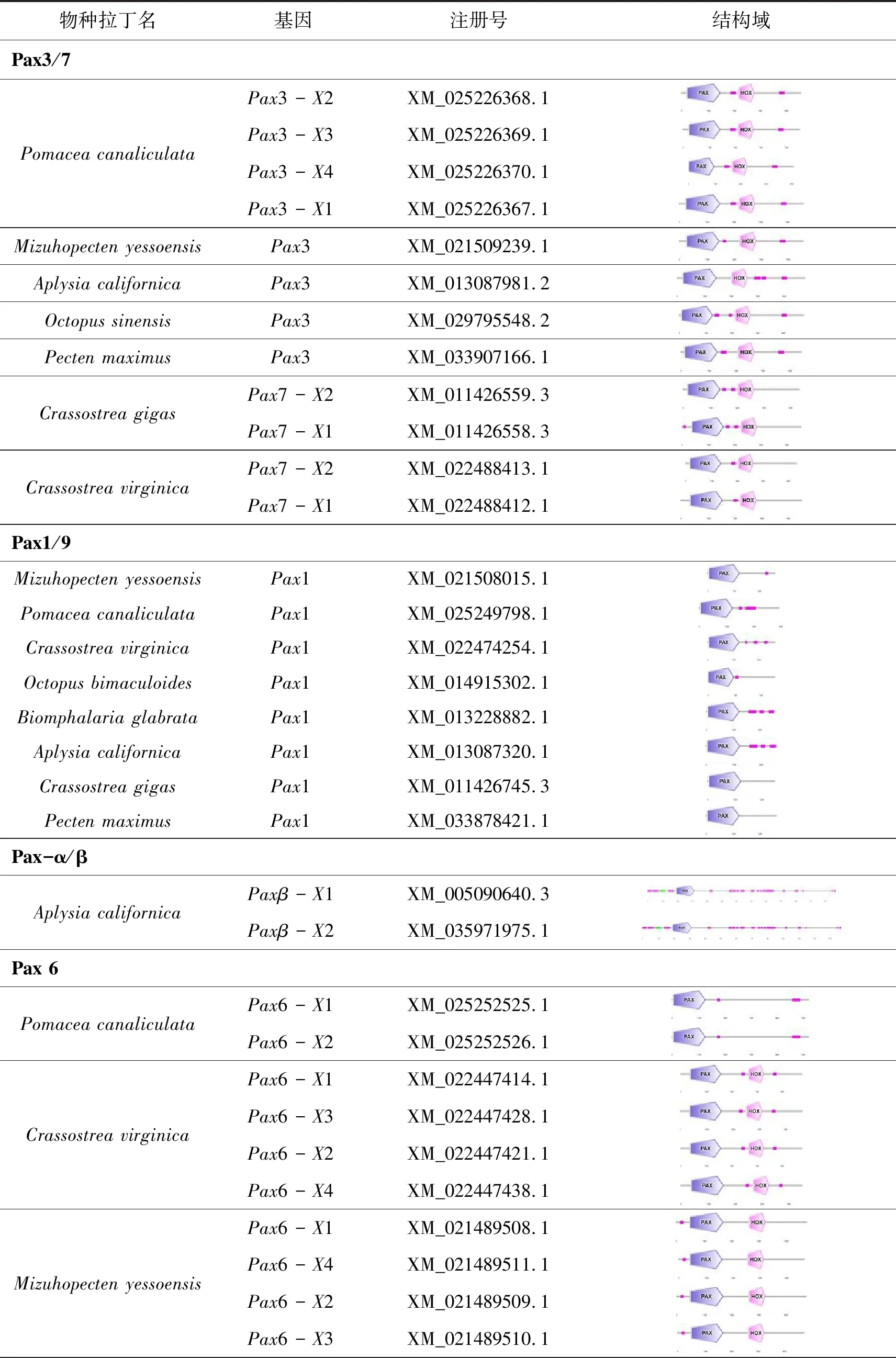
续表
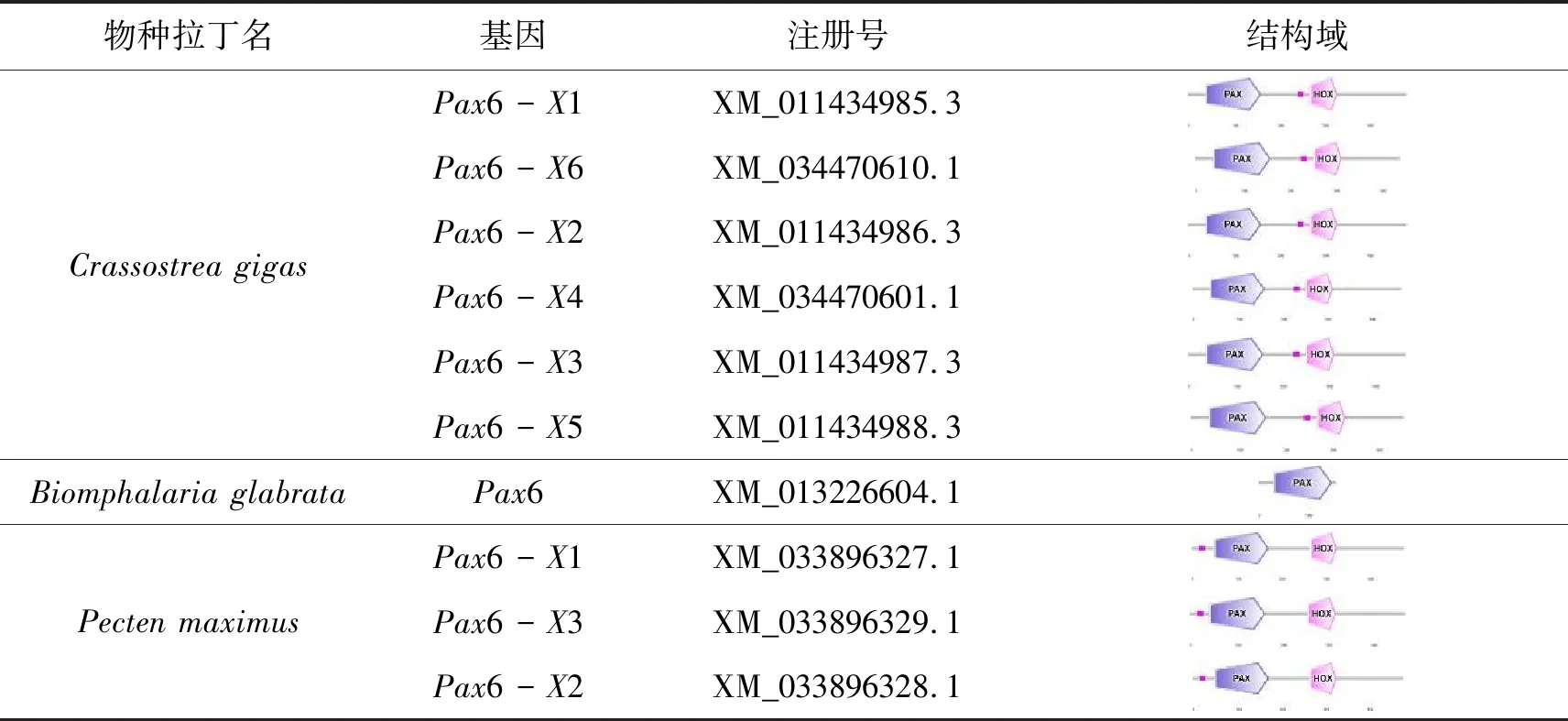
续表

注:箭头表示在红条毛肤石鳖和日本花棘石鳖转录本中鉴定的Pax基因。图4 软体动物的Pax基因系统发生树Fig.4 The neighbour-joining phylogenetic tree of Pax in mollusca
3 讨论
本研究通过RNA-seq技术对红条毛肤石鳖和日本花棘石鳖进行了转录组测序,通过组装分别获得了个121 892和83 776个Unigene,丰富了多板纲分子数据库,为生物学性状的解析奠定了基础;通过对Unigene进行SSR检测,分别筛选到14 602个和15 531个SSR,可以用于开发SSR位点,为多板纲的遗传标记开发提供资源。
Pax基因家族编码一组发育调控相关转录因子,在胚胎发育过程中对组织和器官特化起关键的调控作用。Pax基因通过选择性的剪切方式产生多种亚型,这些亚型的蛋白产物单独或形成多聚体或与其他蛋白结合形成复合体参与细胞的调控。在本研究中,在2个转录本中共鉴定到8条Pax基因,包含5条Pax2/5/8,2条Pax1/9和1条Pax6。不同的Pax亚家族各司其职,例如,Pax1/9在动物的骨骼和胸腺发育过程中发挥重要作用[13];Pax2/5/8参与软体动物中枢神经系统和排泄器官的发育过程[14];Pax6在头足类的眼睛、脑、感觉器官和中枢神经系统的形态发育过程中发挥作用[15]。系统进化树结果显示,多板纲的Pax基因是非常保守的转录因子,在系统进化树中均与相对应的软体动物Pax基因亚族聚为一支,同时也验证了基因注释的准确性。本文筛选的Pax基因不仅补充了对多板纲Pax基因家族的研究,同时也有利于将来解析多板纲独特的形态结构特性。
近年来,基于石鳖在生态学、古生物学、材料学、医药和食品等方面的研究和利用价值,多板纲这一重要海洋生物类群日益受到关注。无论是其食用、药用价值,还是其齿舌磁性纳米材料开发等方面,石鳖都是值得进一步深入研究和发展的水产增养殖品种[16]。然而目前,我国对石鳖的研究才刚刚起步,也尚未对其进行增养殖开发。相对于其他软体动物,人们对石鳖知之甚少,今后应提高对该物种的重视,加强对该物种的理论基础研究,丰富其综合利用途径。本研究获得的转录组数据可为将来多板纲动物的进一步研究提供有用的基础和资源。
猜你喜欢
新民周刊(2022年27期)2022-08-01 07:04:49
华人时刊(2022年3期)2022-04-26 14:29:08
中华手工(2021年2期)2021-09-15 02:21:08
传染病信息(2021年6期)2021-02-12 01:52:58
电影(2019年3期)2019-04-04 11:57:16
财经(2017年2期)2017-03-10 14:35:35
财经(2016年15期)2016-06-03 07:38:02
财经(2016年3期)2016-03-07 07:44:46
财经(2016年6期)2016-02-24 07:41:51
生物医学工程学进展(2015年1期)2015-02-28 14:53:42
