
侧柏水势对干旱胁迫与复水的响应特征
2021-11-18 17:12冯树林周婷黄优刘潇阳吕国利张玉玉王进鑫
江苏农业科学 2021年19期
冯树林 周婷 黄优 刘潇阳 吕国利 张玉玉 王进鑫

摘要:分析干旱和复水环境中植物水势特征,对探讨植物水分状况对土壤水分环境的响应机制具有重要意义。为探索侧柏水势对干旱与复水环境的响应特征,通过温室盆栽试验,研究干旱胁迫与复水环境对侧柏苗木不同生长阶段水势的影响。结果表明,土壤含水量、干旱持续时间、苗木生长时期以及各因素之间的相互作用对侧柏苗木水势均产生了极显著的影响(P<0.01)。随着干旱程度加剧,侧柏苗木水势呈下降趋势,水分胁迫指数呈上升趋势。40.00% SWC处理的苗木水势下降幅度达到最大,其中干旱15 d的生长初期、生长中期和生长后期的苗木水势分别比对照下降了1.46、1.10、1.84 MPa。复水后,侧柏苗木3个生长时期水势逐渐恢复,恢复速率与干旱程度、干旱持续时间和生长时期相关。随着供水时间的延长,复水激发了侧柏苗木水势的补偿作用,苗木生长中期阶段水势恢复快于生长初期和生长后期阶段,在复水72 h后逐渐接近对照水平。
关键词:侧柏;干旱胁迫;复水;水势;恢复
中图分类号:S791.380.1 文献标志码: A文章编号:1002-1302(2021)19-0149-07
土壤干旱是影响植物生长发育的主要环境因素,如果植物生活的土壤环境不能提供正常的水分需求,植物的生长发育、扩散以及相关生理代谢活动都会受到影响,最终导致植物的生长发育受挫[1]。在干旱环境条件下,苗木的生理机能通常会受到伤害,但也因植物品种、干旱程度和干旱持续时间不同呈现一定的差异,而干旱胁迫解除后复水能够一定程度地修复因干旱环境对植物产生的伤害。植物水势能够体现植物抗旱性的强弱,是植物体内水分状况监测的主要生理指标之一,常应用在植物水分关系研究中[2-4]。
前人研究揭示,相同土壤水分环境情况下,如果苗木的水势表现越低,则说明苗木的抗旱性能越好[5]。在干旱环境下,植物可通过改变自身水势调节从土壤中吸收水分的能力,以保障植物正常生长发育的水分需求[6]。研究人员关于灌木植物的研究发现,醉鱼草(Buddleja lindleyana)、花棒(Hedysarum scoparium)、毛条(Caragana korshinskii)、沙木蓼(Atraphaxis bracteata)、杨柴(Hedysarum mongolicum)、四翅滨藜(Atriplex canescens)等苗木水势随土壤干旱程度的加剧呈现下降的变化趋势[7]。杨建伟等对杨树(Populus)水势的研究表明,杨树在轻度水分胁迫和中度水分胁迫下水势缓慢下降,在重度水分胁迫下水势显著下降[8]。Guo等研究发现,沙棘(Hippophae rhamnoides)水势随土壤含水量下降而明显下降,揭示沙棘具有低水势耐旱特性[9]。关于植物在干旱环境条件下的水分生理响应特征,在先前的一些研究报道中已得到证实[10-12]。
侧柏是我国西北地区植被恢复、生态治理和林草融合常用的造林常绿树种和先锋树种[13-14],具有悠久的栽培种植历史,但由于种植地域土壤干旱缺水,其生理功能常常会受到干旱环境的干扰,而基于干旱侧柏苗木复水后不同生长时期的生理响应信息尚不十分完善。基于此,本研究以1龄侧柏苗木为试验对象,通过人工控制土壤含水量的方法模拟干旱胁迫与复水试验,开展土壤水分亏缺对侧柏苗木不同生长时期水势的影响,以及干旱侧柏苗木复水后水势的恢复特征,以揭示侧柏不同生长阶段苗木水势与土壤含水量之间的相关关系,为侧柏在植被恢复、生态治理和林草融合中的高效开发利用提供理论基础和实践支撑。
1材料与方法
1.1试验材料
以1年生侧柏(Platycladus orientalis)苗木为研究对象,选取株高、地径基本一致的健壮营养杯苗作为试验苗木。试验栽培基质为取自陕西杨凌农业高新技术产业示范区崔西沟农田10~20 cm耕作层的土壤,田间持水量为22.3%。苗木栽植前,将栽培土壤充分混匀、风干和过筛,之后存放在科研温室作为苗木栽植土壤。
1.2试验方法
试验区域位于西北农林科技大学温室科学试验区(108°4′E、34°16′N),根据随机区组试验设计,试验设置3个因素,即土壤干旱程度、干旱持续时间和生长时期。以土壤含水量为100%[土壤相对含水量(soil relative water content,SWC)]作为本试验的对照(CK),土壤含水量和干旱历时分别设4个水平,即土壤含水量为 87.84% SWC、70.00% SWC、5216% SWC和40.00% SWC,干旱歷时为15、30、45、60 d。试验采用人工称质量控制土壤含水量的方法进行干旱胁迫与旱后复水试验,研究干旱环境对侧柏生长初期阶段、生长中期阶段和生长后期阶段苗木水势的影响,以及干旱侧柏复水后水势的恢复特征。2017年3月上旬将侧柏幼苗移入深 27 cm、顶部直径29 cm、底部直径27 cm的塑料栽培桶中,每桶装栽培基质10 kg,每个处理3次重复。苗木栽植后,立即给盆栽苗木浇水。根据侧柏苗木的年生长期,分别在苗木的生长初期阶段(4—6月)、生长中期阶段(6—8月)和生长后期阶段(8—10月)实施控水处理,开展土壤干旱胁迫与旱后复水试验。此外,在侧柏其中的一个生长阶段进行干旱胁迫试验时,另外2个时期试验组的苗木盆栽土壤保持100% SWC的土壤含水量。每天17:00通过电子秤(精度1 g)称质量计算苗木栽植土壤减少的水分,将每个处理的土壤含水量补充到试验设定的土壤含水量。试验时间为4—10 月,试验期间采用人工称质量的方法控制侧柏幼苗盆栽土壤含水量,使土壤含水量符合试验设计要求。
1.3指标测定
1.3.1水势用植物水分压力室(Model-1000)分别测定干旱胁迫及旱后复水处理的侧柏苗木生长初期阶段、生长中期阶段和生长后期阶段的水势。根据李继文等的测定方法[15],分别测定侧柏每个生长阶段干旱胁迫处理的水势,以及复水后2、24、48、72 h的水势。
1.3.2水分胁迫指数苗木水分胁迫指数是水分胁迫环境下各指标偏离对照情况的评价指标,可以一定程度反映苗木受水分胁迫的影响情况。水分胁迫指数的值在0~1,其值越大,说明苗木受到水分胁迫的影响越大。苗木水分胁迫指数的测定参照王丁的方法[16]进行。
1.4数据处理
试验数据为每个处理3个重复的平均值,侧柏生长初期阶段、生长中期阶段和生长后期阶段干旱胁迫和复水条件下的苗木水势数据分别用Excel 2021和SPSS 17.0进行统计处理和方差分析,用制图软件Origin 2018进行绘图。
2结果与分析
2.1干旱对侧柏水势的影响
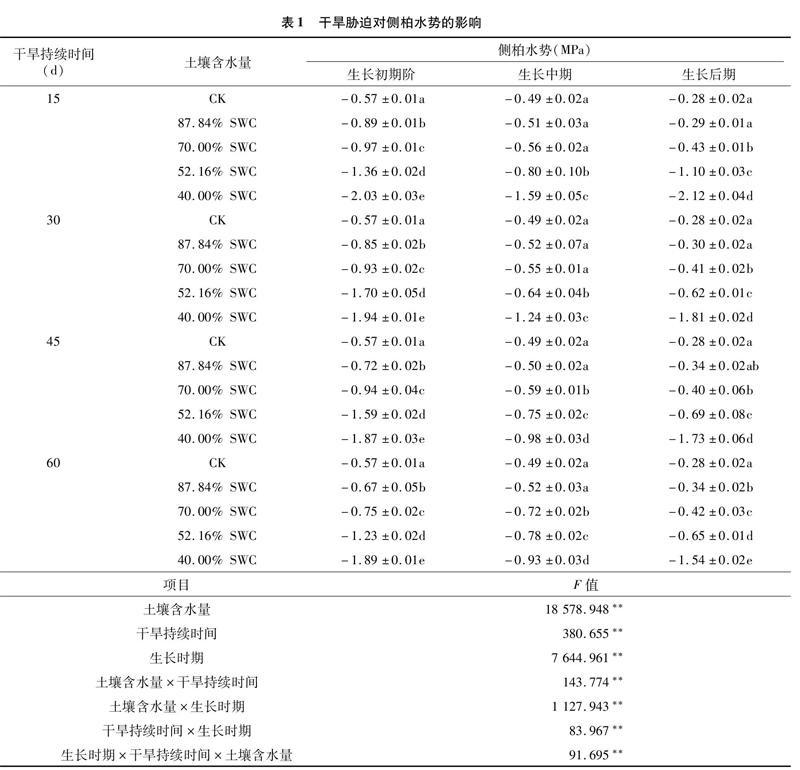
由表1可知,在干旱环境下,侧柏每个生长时期的苗木水势对干旱胁迫的响应特征总体相同,均随着土壤水分亏缺程度的加剧呈现下降趋势,但4个干旱历时下各个水分处理组的水势与对照的差异性水平各有不同。在侧柏生长初期阶段,4个干旱历时下40.00% SWC处理的苗木水势分别比对照减少了1.46、1.37、1.30、1.32 MPa;在侧柏生长中期阶段,4个干旱历时下40.00% SWC处理的苗木水势分别比对照减少了1.10、0.75、0.49、 0.44 MPa;在侧柏生长后期阶段,4个干旱历时下40.00% SWC处理的苗木水势比对照分别减少了1.84、1.53、1.45、1.26 MPa。
显著性检验结果揭示,土壤含水量、干旱持续时间和生长时期对侧柏苗木水势的影响均达到极显著水平(P<0.01),并且,每个因素之间的交互作用也对苗木水势有极显著影响(P<0.01)。试验结果表明,干旱胁迫降低了侧柏苗木3个生长时期的水势,苗木通过调节水势有效增强了对土壤水分的吸收能力。
2.2侧柏水分胁迫指数变化特征
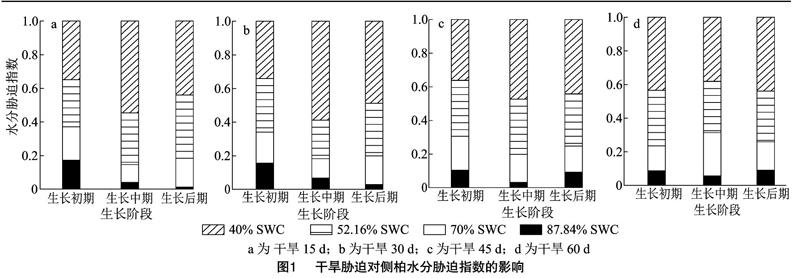
如图1所示,随着土壤含水量的下降,侧柏苗木水分胁迫指数呈上升的变化趋势,但各干旱环境下侧柏苗木生长初期阶段、生长中期阶段和生长后期阶段苗木水分胁迫指数的变化存在差异。
在侧柏苗木生长初期阶段,52.16% SWC和40.00% SWC处理4个干旱历时下侧柏水分胁迫指数的平均值分别达到0.60和0.70,分别是4个干旱历时下87.84% SWC处理侧柏水分胁迫指数平均值的2.34倍和2.73倍,70.00% SWC处理侧柏水分胁迫指数平均值的1.70倍和1.98倍。在侧柏苗木生长中期阶段,52.16% SWC和40.00% SWC处理4个干旱历时下侧柏水分胁迫指数的平均值分别为0.34和0.57,分别是4个干旱历时下87.84% SWC处理侧柏水分胁迫指数平均值的6.18倍和10.32倍,70.00% SWC处理侧柏水分胁迫指数平均值的1.81倍和3.03倍。在侧柏苗木生长后期阶段,52.16% SWC和40.00% SWC处理4个干旱历时下侧柏水分胁迫指数的平均值分别达到0.61和0.84,分别是4个干旱历时下87.84% SWC处理侧柏水分胁迫指数平均值的5.93倍和8.22倍,70.00% SWC处理侧柏水分胁迫指数平均值的1.93倍和2.67倍。
2.3侧柏水势与土壤水分含量的相关性
由图2可知,侧柏的生长初期、生长中期和生长后期3个生长时期,苗木水势与土壤含水量的相关性一致,均表现为正相关关系,但土壤水分对侧柏每个生长阶段苗木水势的影响稍有不同。在侧柏生长初期阶段,干旱历时45 d苗木的相关系数(r2=0.9556)>30d(r2=0.9348)>15d(r2=0.891 4)>60 d(r2=0.841 9);在侧柏生长中期阶段,干旱历时60 d苗木的相关系数(r2=0.904 6)>45 d(r2=0.886 9)>15 d(r2=0.694 3)>30 d(r2=0.633 3);在侧柏生长后期阶段,干旱历时 15 d 苗木的相关系数(r2=0.799 6)>60 d(r2=0.714 9)>45 d(r2=0.697 4)>30 d(r2=0.659 4)。研究结果表明,4个干旱历时下,侧柏苗木水势与土壤含水量均呈线性关系,苗木生长初期阶段、生长中期阶段和生长后期阶段的水势均随着土壤含水量的下降而下降。
2.4復水后干旱侧柏水势恢复特征
由图3可知,旱后复水条件下,侧柏生长初期、生长中期和生长后期3个生长阶段的苗木水势恢复特征总体相近,复水2~72 h,苗木各生长阶段的水势均呈现逐渐升高的变化趋势,但恢复速率存在一定差异。在侧柏的生长初期阶段,复水72 h后,除15、30、45 d干旱历时下52.16% SWC和40.00% SWC处理的水势以及60 d干旱历时下40.00% SWC处理的水势未恢复到对照水平外,其余处理的水势均恢复到接近对照水平。在侧柏的生长中期阶段,苗木水势的恢复与苗木生长初期阶段的恢复稍有不同。复水72 h后,4个干旱历时下各水分处理的苗木水势均恢复到接近对照水平,且与对照差异不显著。在侧柏的生长后期阶段,随着复水时间的增加,4个干旱历时下苗木水势也呈逐渐恢复的变化趋势。复水72 h后,15、30 d干旱历时下苗木水势的恢复速率与生长初期阶段和生长中期阶段稍有不同,45、60 d干旱历时下的苗木水势恢复速率与生长初期阶段不同,与苗木生长中期阶段水势的恢复基本相近。其中,除15、30 d干旱历时下4000% SWC处理的苗木水势未恢复到对照水平外,其余处理的苗木水势恢复到与对照水平相近水平。
研究结果发现,52.16% SWC和40.00% SWC处理的苗木水势在复水72 h后未完全恢复到对照水平,说明干旱侧柏需要更长时间的复水来维持自身水分平衡和生理生化功能的修复。
3讨论与结论
保障植物体内水分平衡是植物正常生长的必备条件,而水势是植物水分生理中的基本参数,可以反映植物水分状况[13,17],能够反映土壤-植被-大气连续系统对植物体内水分的影响程度[18-19]。植株水势越低,吸水能力越强,反之植株水势越高,吸水能力越弱。植物水势的变化能够反映植物从土壤中吸收水分能力的强弱,可用于评价植物的抗旱能力和受干旱胁迫的影响程度,也可作为合理灌溉的生理指标[20]。
相关研究发现,在植物生长发育过程中,均表现出一定的阶段性变化规律,并且在不同生长阶段对环境的响应也存在不同[21-23]。当植物生活在干旱环境时,可通过改变自身水势增强从土壤中吸收水分的能力,同时,植物对干旱环境的响应方式存在差异,水势下降的幅度也各有不同[6]。在本研究中,侧柏3个生长阶段的苗木水势与土壤水分含量的相关性表现一致,均呈现正相关关系。在干旱胁迫条件下,随着土壤水分亏缺程度的加重,侧柏苗木水势呈下降的变化趋势,各生长时期苗木水势的下降幅度有所不同。同时,当土壤水分含量低于70.00% SWC时,侧柏生长初期、生长中期和生长后期阶段的苗木水势均显著下降(P<0.05)。15 d干旱历时下,40.00% SWC 处理的侧柏3个生长阶段的苗木水势在每个生长时期的下降幅度均达到最大,说明干旱侧柏可通过降低自身水势以此来增强
对土壤水分的吸收能力,这与Goto 等对辣椒(Capsicum chinense)的研究结果[24]相似,与吕朝燕等对铁皮石斛(Dendrobium nobile)和金钗石斛(D. officinale)水势的研究结果[1]存在差异。
植物生命活动广泛受到外界各种环境条件的影响,当植物生活的土壤水分环境条件不能满足其正常生理活动时,植物的生长发育就会受到抑制。在土壤水分亏缺环境条件下,干旱环境会对植物的生长产生一定程度的伤害,而抗性强的植物可凭借多种生理代谢活动来调节由干旱胁迫对植物造成的危害,以确保其正常的生长发育。植物水分胁迫指数是苗木受水分胁迫影响程度的重要参数之一,其值越大(變化在0~1之间),揭示植物受干旱环境的影响越大[16]。本试验发现,在土壤水分亏缺条
件下,随着土壤水分亏缺程度的加剧,侧柏生长初期阶段、生长中期阶段和生长后期阶段的苗木水分胁迫指数均呈上升趋势,但各生长时期的苗木水分胁迫指数变化幅度存在差异,这与在香樟(Cinnamomum camphora)、杜英(Elaeocarpus sylvestris)、侧柏、滇柏(Fokienia hodginsii)、构树(Broussonetia papyrifera)、紫穗槐(Amorpha fruticosa)和刺槐(Robinia pseudoacacia)苗木上的研究发现[16,25]相似。同时,在侧柏的生长初期阶段、生长中期阶段和生长后期阶段,40.00% SWC处理对苗木水分胁迫指数的影响较87.84% SWC、70.00% SWC和52.16% SWC处理大,其4个干旱历时处理的苗木水分胁迫指数平均值分别是87.84% SWC处理的苗木水分胁迫指数平均值的2.73倍、10.32倍和8.22倍。
植物在干旱环境中的表现如何,能否在干旱解除后的复水过程中迅速恢复,并恢复由干旱胁迫所产生的损伤是植物对逆境适应性的重要表现[26-27]。王丁等对干旱后喀斯特地区6种造林苗木的研究发现,干旱复水后苗木生长初期、生长中期和生长末期阶段的水势恢复速率存在差异[28]。Schimpl等研究表明,干旱胁迫环境下处理组巴西坚果(Bertholletia excelsa)苗木水势与对照组苗木水势差异显著,复水16 d后干旱处理组苗木水势接近对照水平[29]。此外,张玉玉等对侧柏幼苗的研究发现,干旱处理复水后侧柏幼苗生理代谢能够得到相应恢复,旱后复水激发了植物的生理补偿效应[21,30]。本研究结果表明,不同干旱胁迫环境下侧柏3个生长时期的苗木水势均受到不同程度影响,复水后苗木3个生长阶段的水势恢复程度和恢复速率存在差异。随着干旱胁迫解除后复水时间的延长,侧柏水势呈上升的变化趋势,87.84% SWC和70.00% SWC处理的苗木水势较52.16% SWC和40.00% SWC处理的苗木水势率先恢复到接近对照水平,说明适度的水分亏缺后给干旱植物恢复供水能够激发植物生理的补偿效应。此外,侧柏苗木生长中期阶段的水势在复水后72 h后恢复到接近对照水平,表明侧柏能够有效消除干旱环境对其水势的影响,这与前人对紫穗槐和刺槐的研究结果[31]相似,阐明旱后复水能够激发植物的生理补偿效应,表现出一定的补偿作用。
综上,土壤含水量、干旱持续时间、苗木生长时期3个因素,以及各因素间的交互作用均对侧柏水势有极显著影响(P<0.01)。随着土壤干旱程度的加剧,侧柏生长初期、生长中期和生长后期3个生长时期苗木水势均呈现下降的变化趋势,水分胁迫指数呈上升的变化趋势,且水势和水分胁迫指数的变化因土壤干旱程度、干旱持续时间和苗木生长时期的差异而不同。在干旱环境条件下,侧柏生长初期、生长中期和生长后期阶段的苗木水势显著下降(P<0.05),其中40.00% SWC处理的苗木水势下降幅度最大。干旱胁迫解除后复水,干旱侧柏生长初期、生长中期和生长后期阶段的苗木水势逐渐恢复,旱后复水激发了侧柏苗木的生理补偿机制,水势出现补偿效应,但侧柏3个生长时期的苗木水势恢复有所不同,其中复水72 h后干旱侧柏生长中期阶段的水势均恢复到接近对照水平。
参考文献:
[1]吕朝燕,高智席,严羽,等. 干旱-复水对两种石斛属植物叶水势的影响[J]. 广西植物,2021,41(2):177-182.
[2]Bhusal N,Han S G,Yoon T M.Impact of drought stress on photosynthetic response,leaf water potential,and stem sap flow in two cultivars of bi-leader apple trees (Malus×domestica Borkh.)[J]. Scientia Horticulturae,2019,246:535-543.
[3]Tosin R,Pas I,Novo H,et al. Assessing predawn leaf water potential based on hyperspectral data and pigments concentration of Vitis vinifera L. in the Douro Wine Region[J]. Scientia Horticulturae,2021,278:109860.
[4]武燕奇,郭素娟. 5个板栗品种(系)对持续干旱胁迫和复水的生理响应[J]. 中南林业科技大学学报,2017,37(10):67-74.
[5]刘家琼,邱明新,蒲锦春,等. 我国荒漠典型超旱生植物——红砂[J]. 植物学报,1982,24(5):485-488.
[6]曾凡江,张希明,李小明. 柽柳的水分生理特性研究进展[J]. 应用生态学报,2002,13(5):611-614.
[7]王青宁,衣学慧,王晗生,等. 干旱胁迫下6种固沙灌木叶片水分状况研究[J]. 河南农业科学,2014,43(2):63-67.
[8]杨建伟,梁宗锁,韩蕊莲,等. 不同干旱土壤条件下杨树的耗水规律及水分利用效率研究[J]. 植物生态学报,2004,28(5):630-636.
[9]Guo W H,Li B,Huang Y M,et al. Effects of different water stresses on eco-physiological characteristics of Hippophae rhamnoides seedlings[J]. Acta Botanica Sinica,2003,45(10):1238-1244.
[10]Adnan M Y,Hussain T,Asrar H,et al. Desmostachya bipinnata manages photosynthesis and oxidative stress at moderate salinity[J]. Flora-Morphology,Distribution,Functional Ecology of Plants,2016,225:1-9.
[11]Chen D Q,Wang S W,Cao B B,et al. Genotypic variation in growth and physiological response to drought stress and re-watering reveals the critical role of recovery in drought adaptation in maize seedlings[J]. Frontiers in Plant Science,2016,6:1241.
[12]Liu J W,Zhang R H,Zhang G C,et al. Effects of soil drought on photosynthetic traits and antioxidant enzyme activities in Hippophae rhamnoides seedlings[J]. Journal of Forestry Research,2017,28(2):255-263.
[13]劉潇阳,王进鑫. 保水剂与基质水分对侧柏水分利用特征的影响[J]. 水土保持通报,2020,40(2):205-210.
[14]邴塬皓,刘增文,Trung L N,等. 渭北残塬沟壑区牧草枯落物对油松和侧柏纯林土壤极化的修复效应[J]. 草地学报,2014,22(3):511-517.
[15]李继文,王进鑫,张慕黎,等. 干旱及复水对刺槐叶水势的影响[J]. 西北林学院学报,2009,24(3):33-36.
[16]王丁. 喀斯特地区六种苗木水分生理与抗旱性研究[D]. 南京:南京林业大学,2009.
[17]Dghim F,Abdellaoui R,Boukhris M,et al. Physiological and biochemical changes in Periploca angustifolia plants under withholding irrigation and rewatering conditions[J]. South African Journal of Botany,2018,114:241-249.
[18]付爱红,陈亚宁,李卫红,等. 干旱、盐胁迫下的植物水势研究与进展[J]. 中国沙漠,2005,25(5):744-749.
[19]Wang L,Dai Y X,Sun J Z,et al. Differential hydric deficit responses of Robinia pseudoacacia and Platycladus orientalis in pure and mixed stands in northern China and the species interactions under drought[J]. Trees,2017,31(6):2011-2021.
[20]Martinez C,Roux C,Jauneau A,et al. Effect of water potential on the development of an haploid strain of Sporisorium reilianum f. sp. zeae[J]. Plant and Soil,2003,251(1):65-71.
[21]张玉玉,王进鑫,马戌,等. 土壤干旱及复水对侧柏叶绿素荧光参数的影响[J]. 水土保持研究,2021,28(2):242-247,255.
[22]Dambreville A,Lauri P E,Normand F,et al. Analysing growth and development of plants jointly using developmental growth stages[J]. Annals of Botany,2014,115(1):93-105.
[23]Chen J L,Kang S Z,Du T S,et al. Modeling relations of tomato yield and fruit quality with water deficit at different growth stages under greenhouse condition[J]. AgriculturalWaterManagement,
2014,146:131-148.
[24]Goto K,Yabuta S,Ssenyonga P,et al. Response of leaf water potential,stomatal conductance and chlorophyll content under different levels of soil water,air vapor pressure deficit and solar radiation in chili pepper (Capsicum chinense)[J]. Scientia Horticulturae,2021,281:109943.
[25]馮树林,李博渊,吕国利,等. 紫穗槐幼苗不同生长阶段叶水势对干旱胁迫与复水的响应特征[J]. 草地学报,2020,28(5):1363-1371.
[26]王平,王沛,孙万斌,等. 8份披碱草属牧草苗期抗旱性综合评价[J]. 草地学报,2020,28(2):397-404.
[27]Chen X Y,Luo Y P.The influence of fluctuated soil moisture on growth dynamic of winter wheat[J]. Agricultural Sciences in China,2002,1(1):56-63.
[28]王丁,姚健,杨雪,等. 干旱胁迫条件下6种喀斯特主要造林树种苗木叶片水势及吸水潜能变化[J]. 生态学报,2011,31(8):2216-2226.
[29]Schimpl F C,Ferreira M J,Jaquetti R K,et al. Physiological responses of young Brazil nut (Bertholletia excelsa) plants to drought stress and subsequent rewatering[J]. Flora,2019,252:10-17.
[30]张玉玉,王进鑫,马戌,等. 干旱后复水对干旱侧柏幼苗叶绿素含量的影响[J]. 西南林业大学学报(自然科学),2021,41(5):10-17.
[31]Yan W M,Zhong Y,Shangguan Z P. Rapid response of the carbon balance strategy in Robinia pseudoacacia and Amorpha fruticosa to recurrent drought[J]. Environmental and Experimental Botany,2017,138:46-56.
基金项目:咸阳职业技术学院博士科研基金(编号:2021BK05);国家自然科学基金(编号:31670713);陕西省科技统筹计划(编号:2016KTCL03-18)。
作者简介:冯树林(1987—),男,贵州兴仁人,博士,畜牧师,研究方向为林草生态工程。E-mail:shulinfeng06@163.com。
通信作者:王进鑫,博士,教授,博士生导师,主要从事旱区人工林生态系统水分运移调控与生态恢复理论研究。E-mail:jxwang2002@126.com。
猜你喜欢
山东农业科学(2017年4期)2017-05-18
电脑知识与技术(2016年25期)2016-11-16
水利科技与经济(2016年3期)2016-04-09
水利科技与经济(2016年4期)2016-04-09
水利科技与经济(2016年2期)2016-04-09
江苏农业科学(2014年10期)2014-11-22
江苏农业科学(2014年9期)2014-11-15
水土保持研究(2011年1期)2011-02-11
