
环境因子和颖花构造对水稻颖花关闭的效应
2021-11-18 16:19黄友明曾晓春
江苏农业科学 2021年19期
黄友明 曾晓春
摘要:以可育水稻兴安早占、金优桂99、两优培九和不育系水稻启元S为材料,对离体稻穗进行单色光照射以及不同温度和浸水处理,研究光质、温度和浸水对水稻已开颖花关闭的效应。采用剪取小穗轴长度不等的颖花、去除副护颖和捣碎浆片处理,研究小穗轴长度、副护颖和浆片对颖花关闭的影响。研究表明,红黄绿蓝4种波长的单色光对水稻已开颖花的关闭既无促进也无抑制效应;水稻已开颖花的关闭存在温度“三基点”效应,35~40 ℃最适于颖花关闭,高于45 ℃或低于20 ℃对颖花关闭均不利;水稻成熟颖花浸没在中性介质水中也能开闭,但浸水能一定程度阻止颖花的关闭。小穗轴长度极显著影响颖花关闭,小穗轴越长,颖花关闭越快;副护颖也极显著影响颖花的关闭,去除副护颖,颖花关闭变慢;捣碎浆片则极显著加快颖花的关闭。了解环境因子和颖花构造对水稻颖花关闭的效应对研究水稻颖花关闭机理和颖花关闭调控具有重要意义。
关键词:水稻;颖花;构造;环境因子;关闭;效应
中图分类号: S511.01文献标志码: A文章编号:1002-1302(2021)19-0094-07
水稻穗为复总状花序,由穗轴、一次枝梗、二次枝梗、小穗轴和小穗组成。每个小穗分化3朵颖花,其中2朵在发育过程中退化,退化后各留下外颖,即一般所称的护颖(颖片),小穗基部的2个小突起即退化的颖片,称为副护颖。水稻小花俗称颖花,由外稃、内稃(或外颖和内颖)、1 对浆片、1 枚雌蕊和 6 枚雄蕊组成。内外稃(颖)互相嵌合把颖花内的器官包裹起来,起保护作用。浆片位于颖花基部,在子房与外稃之间[1]。
水稻颖花开闭过程属植物性膨压运动[2],受诸多环境刺激产生的信号影响[3]。许多学者都曾观察过环境因素如温度,湿度、光照等对水稻开闭颖的影响,多数人认为温度是影响水稻开闭颖快慢的主要因素,在一定温度范围内,高温促进开闭颖,低温抑制开闭颖[4-5]。但也有人认為,影响开颖的主要因素不是温度,而是光照。Caldwell等发现大麦颖花从黑暗移至光下会迅速开颖[6-8]。湿度不影响水稻的开闭颖,将颖花放在水中能像空气中那样开闭,将颖花放在干燥器中,浆片也并不提早萎缩。
目前认为,水稻开颖由浆片吸水膨大所启动,浆片的膨大将外稃向外推开,同时将内稃向内挤压,从而使外稃和内稃的钩合点松开,外稃和内稃相互分开,同时,雄蕊花丝急剧伸长,使花药伸出颖壳并裂开,花粉散落,进行授粉;一段时间后,浆片失水萎缩,内外稃又重新合拢,颖花关闭。由于浆片的膨大,推压外颖,外颖进而又压迫小穗轴和副护颖,使小穗轴和副护颖变形而出现伤痕,同时,小穗轴和副护颖对外颖产生了反向弹性力。当浆片失水萎缩时,外颖在小穗轴和副护颖反向弹性力的作用下会向内颖靠拢而闭合[9-11]。
之前的研究主要集中在颖花的开放与调控上,其实,开放后的颖花及其关闭过程也很重要,特别是在水稻育种和杂交制种过程中。正常情况下,水稻的传粉受精是在开颖后进行。杂交制种的重要条件是父母本同时开颖,但与可育水稻相比,不育系水稻颖花开放迟且不整齐,一天中没有明显的开颖高峰,常存在父母本花时不遇的问题。花时不遇严重影响授粉率和异交结实率,从而影响制种产量。开颖授粉后,闭颖的好坏则直接影响子房的发育。不育系水稻的开颖过程,普遍存在开颖至闭颖历时过长、闭颖慢、颖花关闭不严、甚至不闭颖的现象。这一现象会导致不育系颖花的子房长时间暴露在空气中,很容易感染病害;此外,暴露的子房还要忍受风吹、日晒、雨淋、有害化学物质、机械损伤、虫害等因素的影响,往往导致子房发育异常,形成裂颖或子粒畸形,最终影响种子活力和发芽率,降低杂交种子的使用价值。杂交水稻种子存在严重的、普遍的裂颖现象,种子质量不高[12-13]。在杂交水稻制种过程中,种子的产量和质量是制种单位非常关心的问题,而影响种子产量和质量的主要因素就是不育系水稻的开闭颖习性。
研究影响水稻颖花关闭的因子及调控措施,以期解决水稻杂交制种过程中母本颖花开闭习性不良等问题,能为提高杂交种子的产量和质量提供技术支持。研究发现,外施生长素类物质对水稻已开颖花的关闭起抑制效应,而外施脱落酸则起促进效应[14]。本研究从颖花自身构造和其所处的环境因素切入,研究对颖花关闭的效应,以期进一步丰富和发展颖花关闭机理和调控措施。
1材料与方法
1.1供试材料
以可育水稻兴安早占、金优桂99、两优培九和不育系水稻启元S为材料,均常规栽培管理于宜春学院校内试验基地。2018年7—10月研究环境因子对水稻颖花关闭的效应;2019年7—10月研究颖花构造对其关闭的影响。由于常规水稻颖花开闭较集中且历时较短,而不育系水稻颖花开闭不集中且历时较长,为了保证试验的顺利完成、样本颖花生理状态的一致性以及样本颖花所受环境因子的影响相对一致,研究过程中实行多个试验者对同一个水稻品种先进行预备试验后连续重复试验多天的方案。
1.2处理方法
各品种的试验地肥力状况、气象因素和栽培管理一致。在水稻抽穗开花盛期,于晴好天气选择长势和稻穗大小基本一致且顶部颖花已在前一天开放少许、中部颖花将在当天大量开放的穗子。在颖花开放前15 min,随机选择并剪取长势基本一致的成熟稻穗,插入盛清水的桶中,迅速带回实验室。
第1步:把稻穗分别插入盛清水的三角瓶中,每瓶3穗。再把三角瓶置于RDN型人工气候箱内,5个气候箱分别设红光、黄光、绿光、蓝光、白光处理,事先调节好温度为35 ℃、湿度为70%、光照度为 50 000 lx。对处理后的稻穗颖花关闭率和开闭历时进行统计,每处理3瓶,并重复试验3次。
第2步:把稻穗分别插入盛清水的三角瓶中,每瓶3穗。再把三角瓶分别置于6个OBY-H160-SE1恒温恒湿培养箱(160 L),培养箱事先设置温度分别为20、25、30、35、40、45 ℃,湿度为70%,光照度为50 000 lx。对处理后的稻穗进行颖花关闭率和开闭历时统计,每处理3瓶并重复3次。
第3步:把稻穗分成2个部分,其中一部分穗杆基部系重物,让整个稻穗浸没在盛满35 ℃清水的 1 000 mL 量筒中,再把量筒置于35 ℃的恒温水浴锅中。为便于观察,每量筒1穗,3个量筒即3穗为1个处理,重复试验3次。把另一部分稻穗基部插入盛清水的三角瓶中,颖花露在空气中,再把三角瓶置于35 ℃恒温恒湿培养箱中,设置培养箱的湿度为100%,光照度与实验室的光照度相同。每3穗为1个处理,重复试验3次。对处理后的稻穗进行颖花关闭率和开闭历时统计。
第4步:待内外颖夹角最大时,迅速剪取小穗轴长度分别为0、1、2、3 cm的颖花各50朵,分别置于盛35 ℃清水的培养皿中,颖花浮在水面上,再把培养皿悬浮于35 ℃的恒温水浴锅中。对处理后的颖花进行开闭历时统计,重复3次。
第5步:待内外颖夹角最大时,迅速将颖花副护颖精准剪除,同时选择同穗且位置邻近的开放颖花作对照,保留副护颖。对处理后的颖花进行开闭历时统计,每处理50朵颖花,重复3次。
第6步:待内外颖夹角最大时,迅速将颖花浆片精准捣碎,同时选择同穗且位置邻近的开放颖花作对照,不捣碎浆片。对处理后的颖花进行开闭历时统计,每处理50朵颖花,重复3次。
1.3颖花关闭率动态及开闭历时的测定
1.3.1颖花关闭率各时间点累计关闭的颖花总数占试验当天开放颖花总数的百分数(%)即为颖花关闭率。从当天开放的颖花关闭第1朵时开始(0 min),每30 min累计当天开放颖花的关闭总数,直到当天开放的颖花不再关闭时为止。计算出各处理每时间点的颖花关闭率以及各时间点的平均颖花关闭率和标准差,重复3次。
1.3.2颖花开闭历时颖花内外稃夹角大于0 ℃所经历的时间即为颖花开闭历时。观察记录内外颖始裂开与完全合拢的时刻,计算单朵颖花开闭历时和每处理50朵颖花开闭历时的平均值和标准差,重复3次。对颖花开闭状态判断采用相同的标准。
1.4数据分析
颖花关闭率和颖花开闭历时等数据均采用Microsoft Excel软件作图,并对重复试验的平均数作标准误差分析。对颖花开闭历时经F检验后用最短显著极差法(shortest significant ranges,SSR)对各平均值进行多重比较,作出统计推断,比较结果用标记字母法表示。
2结果与分析
2.1环境因子对水稻颖花关闭的效应
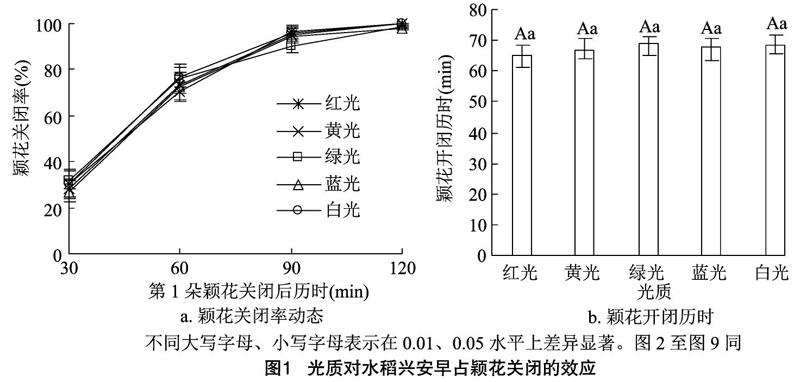
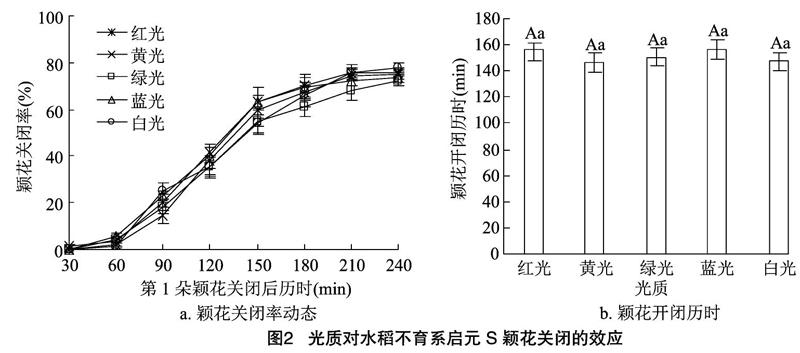
2.1.1光质对水稻颖花关闭的效应由图1、图2可知,兴安早占和启元S类似,不同光质处理各时间点已开颖花的关闭率,与对照白光处理无明显差别;不同光质处理颖花开闭历时差异不显著。不同波长的光处理对水稻颖花关闭率既无提高也无降低的明显变化,对颖花开闭历时既无延长也无缩短的显著差异,故对水稻颖花关闭既无促进也无抑制效应。
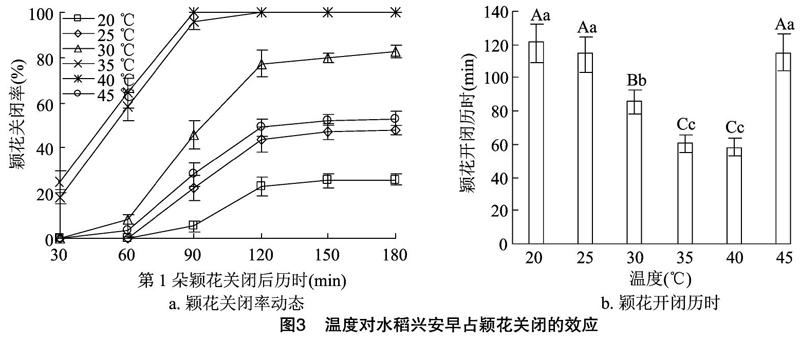
2.1.2温度对水稻颖花关闭的效应由图3、图4可知,兴安早占和启元S一样,在不同温度处理下,各时间点已开颖花的关闭率存在明显差异,20 ℃处理最低(启元S已开颖花20 ℃处理不关闭),25 ℃和45 ℃处理相当,但较前者明显升高,30 ℃处理又较前者明显升高,35、40 ℃处理相当且最高,较前者也存在非常明显的差异。各处理之间的颖花平均开闭历时存在极显著差异,25、45 ℃处理之间无显著性差异,但均极显著长于30 ℃处理,30 ℃处理又极显著长于35、40 ℃处理,35、40 ℃处理之间无显著性差异。表明不同温度处理对水稻颖花关闭影响差异非常明显,其中35~40 ℃处理颖花关闭率最高,颖花开闭历时最短,最适于颖花的关闭,高于 45 ℃ 或低于20 ℃对颖花关闭均是不利的。
2.1.3浸水对水稻颖花关闭的效应由图5、图6可以看出,兴安早占和启元S一样,已开颖花完全浸没在水中也能关闭,但各时间点的颖花关闭颖率明显较空气中低。可育水稻颖花开闭历时,经F检验,0.01 2.2颖花构造对水稻颖花关闭的效应 2.2.1小穗轴长度对颖花关闭的效应由图7可知,随小穗轴长度增加,颖花开闭历时缩短。兴安 早占小穗轴1 cm颖花开闭历时极显著短于小穗轴0 cm颖花,而极显著长于小穗轴2 cm和3 cm颖花,小穗轴2 cm与3 cm的颖花开闭历时则差异不显著。金优桂99和两优培九小穗轴0~1 cm之间,以及2~3 cm之间颖花开闭历时差异不显著,但0~1 cm 颖花的开闭历时极显著长于2~3 cm的颖花。兴安早占小穗轴长1、2、3 cm时的颖花平均开闭历时分别为57.83、48.91、4521 min,较0 cm长的6508 min分别缩短7.25、16.17、19.87 min,缩短百分比分别达11.14%、2485%、30.53%。金优桂99小穗轴长2、3 cm时的颖花平均开闭历时分别为50.01、47.44 min,较0 cm长的62.14 min分别缩短12.13、14.70 min,缩短百分比分别为19.52%、2366%。两优培九小穗轴长2、3 cm时的颖花平均开闭历时分别为50.00、46.39 min,较0 cm长的7100 min分别缩短21.00、24.61 min,缩短百分比分别为29.58%、34.66%,表明小穗轴的长短显著影响颖花关闭,小穗轴越长,颖花开闭历时越短。 2.2.2副护颖对颖花关闭的效应由图8可知,剪除副护颖处理较保留副護颖延长了颖花开闭历时。兴安早占和两优培九处理的去副护颖颖花开闭历时极显著长于留副护颖;金优桂99是去副护颖处理的颖花开闭历时显著长于留副护颖。表明颖花副护颖有助于颖花关闭。 2.2.3浆片对颖花关闭的效应由图9可知,与完好浆片处理相比,颖花开放后立即捣碎浆片处理明显缩短了颖花开闭历时。兴安早占和两优培九捣碎浆片处理的颖花开闭历时显著短于完整浆片,金优桂99则捣碎浆片处理的颖花开闭历时极显著短于完整浆片。表明颖花开放后捣碎浆片有助于颖花关闭。 3讨论与结论 水稻已开颖花的关闭是一个相当复杂的生理过程,涉及浆片细胞内容物的分解、合成和运输[15];也涉及蔗糖转运蛋白以及相关离子转运蛋白等相关基因的表达与活化[16];还与呼吸有关,呼吸作用促进剂能促进颖花的关闭,而呼吸作用抑制剂能抑制颖花的关闭[4]。本研究发现,35~40 ℃的气温最适于颖花的关闭,高于45 ℃或低于20 ℃的气温对颖花关闭均不利,这完好印证了颖花关闭过程相关酶的温度“三基点”效应。光照处理黑暗中适应的 成熟稻穗并不诱导开颖[5],这可能是因为颖花的开放需要消耗能量与呼吸作用,而黑暗中积累的ATP较少。但在黑麦上的试验不同,光照是控制黑麦开花的关键因素,黑暗或弱光下,成熟麦穗的开花被限制,但暴露在充足的阳光下几分钟即可诱导开花[17]。水稻颖花的关闭与其开放是2个不同的过程,已开颖花置于黑暗中仍能关闭,但较光下有所抑制,这进一步证明水稻颖花的关闭也是一个需要消耗能量且与呼吸作用有关的过程。本研究采用不同波长的单色光照射处理水稻已开颖花,表现出相同的关闭效应,说明光照对颖花关闭的效应并不在于辐射热,还说明颖花关闭过程不属于光形态建成现象,与光敏色素等光受体无关。光照、温度和湿度经常联系在一起影响植物的生长发育。Pissarek发现,生长于恒定光、温和湿度气候箱内的黑麦颖花将随机开放,但在完全黑暗中不能开放[15]。然而,这种穗经30~3 000 lx光照处理几分钟后,100%能够开花,少数颖花甚至能响应强度低至0.6 lx的光照。本研究发现,水稻已开颖花完全浸没在清水中也能关闭,但较空气中变慢。这与相对湿度在40%~100%范围内变化,不影响开颖的结论类似[4]。同属禾本科植物的水稻和黑麦,开颖和闭颖对光照的反应不同,环境因素对水稻颖花关闭的作用机理值得进一步研究。 与颖花关闭直接关联的自身构造主要涉及浆片、副护颖和小穗轴。浆片膨大颖花开放,浆片缩小颖花关闭[2,9],浆片的萎缩是细胞吸水后细胞膜破裂,发生自溶引起,牵涉到大量酶促反应的参与[9],其机理可能与叶片保卫细胞体积和形状改变导致气孔开闭类似,涉及淀粉与糖的转化、离子积累与撤离和大量有机酸的转化等[18]。人为捣碎已开颖花的浆片,对颖花关闭效应与自然萎缩类似,佐证了颖花关闭是由浆片体积缩小引起。浆片细胞体积的变小除了与细胞膜破裂、发生自溶有关外,还与浆片内容物的撤离和小穗轴的主动运输有关[10]。小穗轴内具有发达的维管束系统[9-10]。内外颖、浆片、花丝、子房等器官都着生在小穗轴上,这些器官的维管束与小枝梗的维管束在小穗轴处汇集,形成错纵复杂的输导体系,由它控制物质的运输,它既可以把来自小枝梗的养分和水分泵向颖花内各器官,又可以进行各器官养分的相互调配[9]。开颖时,可能通过它向浆片和花丝输送养分和水分,开颖后又通过它把浆片中的物质抽回调运到其他器官。因此,切除小穗轴就会表现出闭颖慢或不能闭颖的现象。浆片膨大推压外颖,外颖进而又压迫小穗轴和副护颖,使小穗轴和副护颖变形而出现伤痕,与此同时,小穗轴和副护颖又对外颖产生了反向弹性力[9-11]。颖花开放后剪除副护颖,使副护颖挤压外颖的弹性力消失,故颖花关闭慢。 不育系水稻颖花开放高峰不如可育水稻明显,日开花较分散,開颖不集中导致闭颖也不集中,一天之中无明显的闭颖高峰[19-20],有的颖花甚至在夜间开闭颖。颖花关闭缓慢,开闭历时较长,还有闭颖不严、裂颖现象严重等特点。不育系水稻雄蕊发育畸形,花粉败育,呼吸代谢较弱,颖腔内CO2浓度不容易升高以及释放的能量有限。不育系的浆片和小穗轴维管系统不发达,以及薄壁细胞酶活性很低[9-10,21-23],导致浆片吸水和失水均较慢,开颖后浆片较长时间仍然保持膨胀状态,小穗轴和外颖基部的细胞结构生长定型后,小穗轴就失去了恢复到原位的弹性力,导致颖壳不能关闭或关闭不良。由浆片渗透压调节其水势变化引起浆片吸水膨大而颖花开放,闭颖运动则主要受小穗轴控制,剪除小穗轴的颖花因为浆片内水分等内容物不能外运而抑制闭颖[9]。 了解水稻颖花开闭习性和影响颖花开闭的自身构造及环境因子,有助于对存在颖花开闭障碍的水稻和逆境胁迫下颖花开闭不正常的水稻进行颖花开闭调控,对解决杂交水稻种子裂颖问题和提高种子产量和质量非常重要。 参考文献: [1]金银根,周桂香,王增春,等. 水稻小穗的结构及其功能[J]. 植物学通报,1996,31(4):34-37. [2]曾晓春,周燮,吴晓玉. 水稻颖花开放机理研究进展[J]. 中国农业科学,2004,37(2):188-195. [3]Haupt W,Feinleib M E. Encyclopedia of plant physiology new series volume 7:physiology of movements[M]. New York:Springer,1979. [4]顾蕴洁,王忠,高煜珠. 环境因素对水稻颖花开闭影响的机理[J]. 植物生理学报,1993,19(4):345-352. [5]王忠,卢从明,顾蕴洁,等. 水稻开颖机理的探讨Ⅰ.温度对水稻开颖及花粉生活力的影响[J]. 作物学报,1988,14(1):14-21. [6]Caldwell M M.Solar uv irradiation and the growth and development of higher plants[M]. Amsterdam:Elsevier,1971. [7]Cline M G,Salisbury F B.Effects of ultraviolet radiation on the leaves of higher plants[J]. Radiation Botany,1966,6(2):151-163. [8]Hashimoto T,Tajima M.Effects of ultraviolet irradiation on growth and pigmentation in seedlings[J]. Plant & Cell Physiology,1980,21(8):1559-1571. [9]王忠,顾蕴洁,高煜珠. 水稻开颖机理的探讨:Ⅲ.浆片的结构及其在开颖过程中内含物的变化[J]. 作物学报,1991,17(2):96-101,161. [10]王忠,顾蕴洁,高煜珠. 水稻开颖机理的探讨:Ⅳ.水稻小穗轴的结构及其与开闭颖的关系[J]. 作物学报,1992,18(5):331-336,401. [11]王忠,顾蕴洁,高煜珠. 水稻小穗轴的结构及其在开闭颖过程中的变化[J]. 江苏农学院学报,1994,15(1):1-10. [12]刘晓霞,陈立云. 杂交水稻种子裂颖特性研究进展[J]. 作物研究,2005,19(增刊1):278-283. [13]王元凯,李贤勇. 杂交水稻种子闭颖度对种子耐储力的影响[J]. 西南农业学报,2006,19(4):565-567. [14]Huang Y M,Zeng X C,Cao H P. Hormonal regulation of floret closure of rice (Oryza sativa)[J]. PLoS One,2018,13(6):e0198828. [15]Pissarek H P. Untersuchugenúber bau und funktion dergramineen lodiculae[J]. Beitrage zur Biologie der Pflanzen,1971,47:313-370. [16]何永明,曾晓春,向妙莲,等. 水稻花时调控研究进展[J]. 湖北农业科学,2014,53(7):1489-1492. [17]Dubcovsky J,Loukoianov A,Fu D L,et al.Effect of photoperiod on the regulation of wheat vernalization genes VRN1 and VRN2[J]. Plant Molecular Biology,2006,60(4):469-480. [18]许祥明,王忠,顾蕴洁,等. 某些酸类物质对水稻开颖的效应[J]. 植物生理学报,1998,24(2):124-130. [19]陈雄辉,万邦惠,陆燕鹏,等. 光温敏核不育水稻异交结实潜力的研究[J]. 华南农业大学学报,2001,22(1):1-4. [20]文廷刚,王伟中,杨文飞,等. 水稻穗分化期外源植物生长调节剂处理对颖花分化与退化的影响[J]. 江苏农业学报,2019,35(3):514-522. [21]王忠,顧蕴洁,于红亮,等. 野败型不育系水稻畸形籽粒形成原因的探讨[J]. 中国农业科学,1995,28(6):25-31. [22]王忠,金银根,许祥明,等. 野败型不育系稻穗闭颖慢原因的探讨[J]. 中国农业科学,1992,25(2):22-26. [23]王忠,周振兴,张学兵,等. 献改A闭颖难及F1籽粒畸形原因探讨[J]. 杂交水稻,1991(6):38-40. 基金项目:国家自然科学基金(编号:31360297);江西省高校落地计划(编号:KJLD12092)。 作者简介:黄友明(1972—),男,江西上高人,博士,副教授,从事植物激素生理和作物化学调控研究。E-mail:huangyouming52@126.com。 通信作者:曾晓春,博士,教授,博士生导师,从事作物生理研究。E-mail:xchzeng2013@163.com。
猜你喜欢
江西农业大学学报(2023年2期)2023-05-08
作物杂志(2022年6期)2022-02-03
作物学报(2021年10期)2021-08-05
北方水稻(2021年6期)2021-02-17
中国科技纵横(2016年20期)2016-12-28
中国市场(2016年36期)2016-10-19
科技视界(2016年2期)2016-03-30
河北渔业(2015年10期)2015-10-15
