
染料木黄酮抑制AKR1C3抗去势抵抗前列腺癌的生长及其机制研究
2021-11-14 11:23伏天雨周恒平孙永霞陈思琪朱彦锋
食品工业科技 2021年22期
陈 晋,伏天雨,刘 江,周恒平,孙永霞,陈思琪,艾 丽,朱彦锋
(成都医学院公共卫生学院,四川成都 610500)
前列腺癌是中老年男性常见肿瘤之一,中国前列腺癌的发病率也逐年增加[1-2]。前列腺癌是一类雄激素依赖性肿瘤[3]。早期发现雄激素剥夺治疗(ADT)以降低睾酮水平能有效抑制前列腺癌的生长,但后续证实,经过ADT后,前列腺癌会发展成为去势抵抗前列腺癌(CRPC),且患者生存期明显降低[4-6]。因此,CRPC的防治是现今前列腺癌研究的重点[3,7]。现阶段,临床上有很多化疗药、激素抑制剂、肿瘤疫苗用于治疗CRPC[7-9]。但这些化学合成药物治疗后均带来副作用。因此,寻找能够用于治疗CRPC的低毒性药物,对于CRPC的临床治疗具有重要意义。
Aldo-keto还原酶家族1成员C3(AKR1C3)是一种类固醇合成酶,是雄激素(T和DHT)合成最后两步的关键酶[10-11]。前列腺癌作为一种雄激素依赖性的肿瘤,雄激素的含量变化对其生长具有较大的影响[12],文献表明,在CRPC组织中AKR1C3 mRNA和蛋白水平均有上调,AKR1C3 siRNA能够抑制AKR1C3 mRNA,进 而 抑 制CRPC的 生 长[13-14]。AKR1C3的异常表达不仅能对恶性肿瘤和内分泌疾病产生影响,更与CRPC的恶性程度直接相关,是前列腺癌进展成为CRPC的适应性反应[6,15-16]。近期研究发现,AKR1C3抑制剂在癌症领域中的使用逐渐广泛,对于抑制AKR1C3表达来降低雄激素合成的方式,可作为抗CRPC的新靶标[17-18]。
染料木黄酮(GEN)作为一种高活性的天然植物雌激素,在大豆中的含量高于其他植物。GEN具有多种分子效应,如抑制炎症、促进细胞凋亡、调节甾体激素受体和代谢途径[19]。有学者通过研究证实染料木黄酮不仅能影响癌细胞的增殖,也能减轻炎症和脂肪的生成,预防肥胖和代谢综合征[20-22]。所以GEN具有的多种分子效应在预防和治疗常见的疾病中发挥着重要的作用。
近年来,GEN被证实具有抑制CRPC细胞增殖的效应[23],但其作用机制尚未完全阐明。为探究GEN抑制AKR1C3的表达,继而发挥抗去势抵抗前列腺癌生长的作用机制。本研究从雄激素合成途径出发,拟通过膳食中最常见的大豆异黄酮活性单体GEN处理CRPC细胞,利用细胞培养、蛋白印记及其他分子生物学技术方法,从体外探讨GEN对去势抵抗前列腺癌增殖影响及GEN降低AKR1C3蛋白的表达,研究大豆异黄酮类植物激素物质抗去势抵抗前列腺癌的新机制,为预防和治疗去势抵抗前列腺癌丰富理论基础。
1 材料与方法
1.1 材料与仪器
人去势抵抗前列腺癌22RV1细胞、人去势抵抗前列腺癌VCaP细胞、正常前列腺上皮细胞RWPE-1细胞 中国科学院;1640培养基、DMEM培养基
中国上海;胎牛血清 美国Gibco;染料木黄酮干粉、AKR1C3抑制剂(ASP-9521) MCE(中国);AKR1C3 siRNA 吉玛基因(中国上海);转染试剂盒
赛默飞(美国);CCK-8试剂盒、IP细胞裂解液碧云天(中国);BCA蛋白定量试剂盒 Solarbio(中国上海);PSA单抗、AKR1C3单抗(Abcam);BACTIN单抗 中杉金桥。
电泳装置 Bio-Rad(美国);冷冻离心机、酶标仪、凝胶成像仪 Thermo(美国)。
1.2 实验方法
1.2.1 细胞培养 培养22RV1、VCaP和RWPE-1细胞株,22RV1用无酚红RPMI-1640培养基(含10%的活性炭-葡聚糖处理的胎牛血清)培养,VCaP细胞用无酚红DMEM培养基(含10%的活性炭-葡聚糖处理的胎牛血清)培养,RWPE-1用RPMI-1640培养基(含10%的胎牛血清)培养,细胞培养环境为37 ℃、5% CO2的培养箱中。
1.2.2 细胞转染 取对数生长的22RV1、VCaP细胞,按每孔2×105个接种至6孔板底部,待密度长至80%时更换培养液加入无酚红RPMI-1640、DMEM培养基1.5 mL,分别用5 μg的小干扰RNA,AKR1C3-homo-77、AKR1C3-homo-447和 AKR1C3-homo-1008与5 μL转染试剂联合处理,置于培养箱培养4~6 h后在荧光显微镜下观察NC孔,如有荧光且在细胞中分布均匀后,则将每孔用PBS润洗两次后换DMEM完全培养基继续培养48 h。48 h后进行细胞裂解并提取蛋白质,进行Western blot检测[24]。如表1所示为AKR1C3 的siRNA引物序列。

表1 AKR1C3 的siRNA引物序列Table 1 siRNA primer sequences of AKR1C3
1.2.3 CCK-8法检测细胞生长情况 取对数生长期的22RV1、VCaP和RWPE-1细胞接种于96孔板中培养24 h,37 ℃、体积分数5%CO2、饱和湿度条件下培养24 h,次日用无菌PBS润洗后,加入(0、12.5、25、50、100)μmol/L的GEN工作液处理48 h,每组设5个复孔。加入工作液24、48、72 h 后,分别加入含CCK-8 的培养基100 μL孵育4 h。采用酶标仪测定每孔吸光度(OD),波长设为450 nm[25],按公式进行计算:细胞活性(%)=(OD加药-OD空白)/(OD对照-OD空白)×100。
1.2.4 小干扰RNA和AKR1C3抑制剂对AKR1C3表达量的影响 为证实GEN抗去势抵抗前列腺癌细胞的增殖是通过抑制AKR1C3的表达实现的,用AKR1C3 siRNA和AKR1C3抑制剂(ASP-9521)处理细胞,观察二者的效果。体外培养22RV1、VCaP细胞,按每孔2×105个接种至六孔板底部,待密度长至80%时更换培养液,加入无酚红RPMI-1640、DMEM培养基1.5 mL,用最优AKR1C3 siRNA 447进行试验,使用5 μg AKR1C3 siRNA 447和GEN(50 μmol/L)单独或联合处理48 h。使用50 nmol/L AKR1C3抑制剂(ASP-9521)和GEN(50 μmol/L)单独或联合处理48 h,Western blot分析22RV1、VCaP细胞中AKR1C3的表达[26]。
1.2.5 Western blot法检测 用5个不同浓度(0、12.5、25、50、100)μmol/L GEN处理细胞48 h后,加入200 μL的IP裂解液,放置在冰上裂解30 min,在4 ℃下12500 r/min离心10 min,得到上清液置于-20 ℃保存备用。制备SDS-PAGE凝胶,在80 V(浓缩胶)/120 V(分离胶)恒压条件下分离蛋白质,在250 mA恒流转膜90 min。PVDF膜用20 mL封闭液放置于常温下封闭2 h,取出PVDF膜用1×TBST清洗3次,每次5 min,裁剪所需要条带并放入相应的一抗中,4 ℃封闭过夜。次日取出条带,用1×TBST溶液洗膜3次,每次6 min,再用二抗,室温孵育90 min。用1×TBST溶液洗膜3次,每次6 min,准备化学发光成像[26]。
1.3 数据处理
2 结果与分析
2.1 GEN对CRPC细胞活力的抑制作用
GEN作为一种植物雌激素,能够有效抑制去势抵抗前列腺癌细胞的增殖、抑制转移和诱导凋亡[26-27]。研究结果显示,GEN(50、100 μmol/L)处理后,能够明显抑制22RV1、VCaP细胞增殖(图1A)。为进一步研究GEN对CRPC细胞功能的影响,以Western blot测定PSA蛋白的表达。PSA作为前列腺癌治疗的分子目标,是因为其不仅活跃在前列腺组织,在前列腺癌信号通路也会产生扩散、入侵、转移、血管生成、细胞凋亡、免疫反应、肿瘤微环境监管等影响,因此血清中PSA水平与疾病进展相关[28]。本研究结果显示,GEN对RWPE-1细胞活力无显著的抑制作用(P>0.05),GEN(50、100 μmol/L)对PSA表达具有显著抑制作用(P<0.05)(图1B)。
2.2 染料木黄酮对22RV1、VCaP细胞中AKR1C3表达的抑制作用
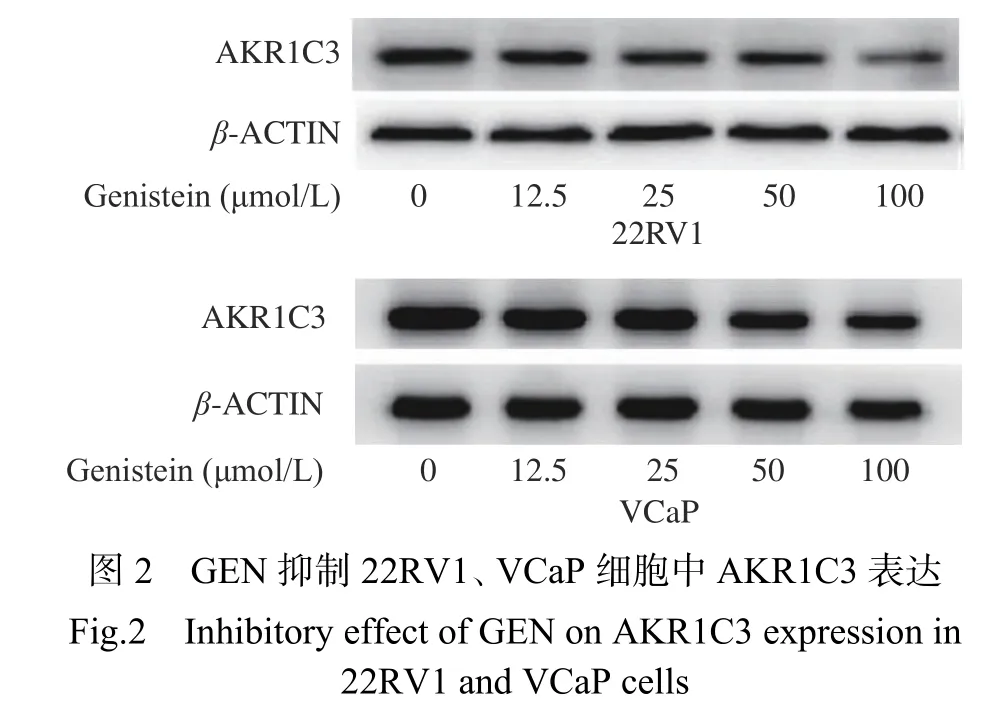
研究表明,在前列腺癌转化为CRPC的过程中,会产生更多的睾酮,AKR1C3呈异常高表达,将导致更多具有活性的雄激素合成[29],因此AKR1C3的异常表达可能是前列腺癌由激素依赖性向CRPC转化的生物学标志。结果显示(图2),GEN(25、50、100 μmol/L)处理后条带灰度减弱,因此GEN(25、50、100 μmol/L)对AKR1C3表达具有明显抑制效果。结合图1结果,GEN处理后,GEN 50 μmol/L时首次出现能够明显抑制22RV1、VCaP细胞增殖及PSA表达的现象,因此选用GEN浓度为50 μmol/L进行后续实验。
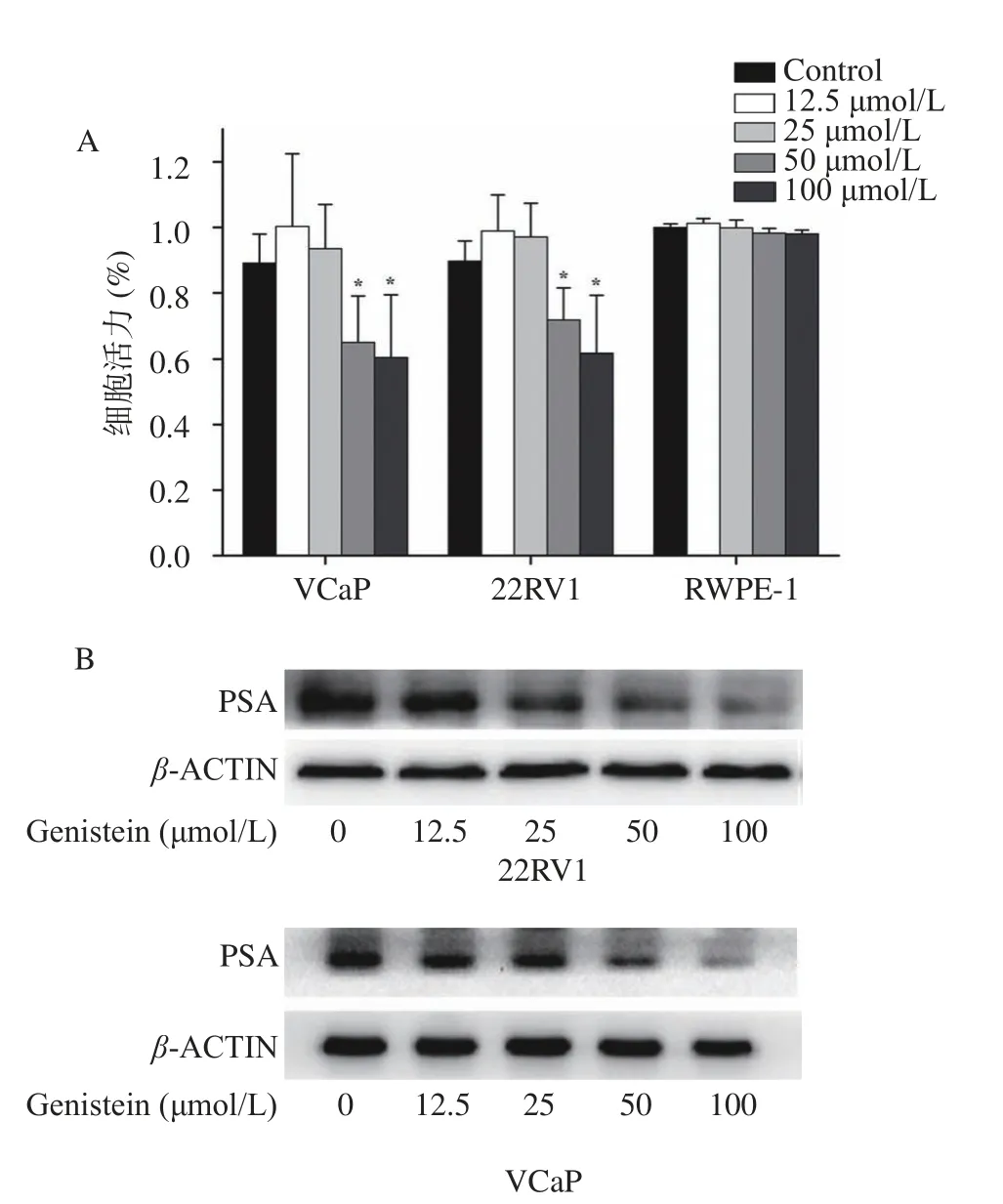
图1 GEN对22RV1、VCaP细胞增殖能力的影响Fig.1 Effect of GEN on the proliferation ability of 22RV1 and VCaP cells
2.3 AKR1C3 siRNA447对22RV1、VCaP细 胞 中AKR1C3表达的抑制作用
图3 结果显示,综合AKR1C3siRNA抑制22RV1、VCaP细胞中AKR1C3表达的结果,可以看出AKR1C3 siRNA447的条带灰度减弱最明显,因此AKR1C3 siRNA447对AKR1C3的抑制效果最为明显。故选用AKR1C3 siRNA447进行后续实验。
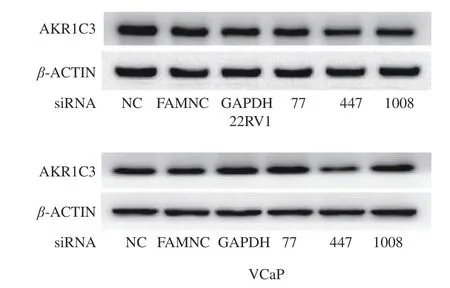
图3 AKR1C3 siRNA抑制22RV1、VCaP细胞中AKR1C3表达的结果Fig.3 Inhibitory effect of AKR1C3 siRNA on AKR1C3 expression in 22RV1 and VCaP cells
2.4 AKR1C3 siRNA 447和GEN(50 μmol/L)对22RV1、VCaP细胞中AKR1C3表达的抑制作用
如图所示(图4),与对照组相比,样品组条带灰度明显低于对照组,且联合使用时现象更为明显。由此推断,AKR1C3 siRNA447、AKR1C3抑制剂(ASP-9521)和GEN(50 μmol/L)对AKR1C3均有抑制作用,且AKR1C3 siRNA447和AKR1C3抑制剂(ASP-9521)分别与GEN(50 μmol/L)联合作用时效果更强(图5)。由此推测,GEN对AKR1C3的抑制机制与AKR1C3 siRNA、AKR1C3抑制剂(ASP-9521)抑制AKR1C3相似。类似Pippione等[18]发现AKR1C3抑制剂可拮抗DHEA诱导的DuCaP生长,以非选择性的方式抑制AKR1C3的表达;以及Batra等[30]发现染料木素联合多糖可抑制前列腺癌细胞内雄激素的合成。
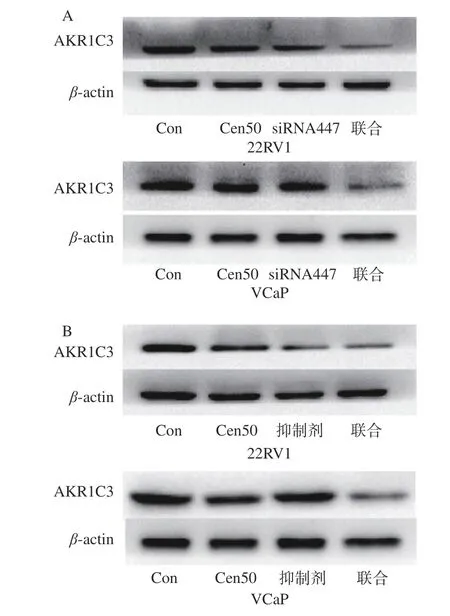
图4 AKR1C3抑制剂(ASP-9521)和GEN(50 μmol/L)联合处理对22RV1、VCaP细胞中AKR1C3的表达的影响Fig.4 Effect of combined treatment of AKR1C3 inhibitor(ASP-9521) and GEN (50 μmol/L) on the expression of AKR1C3 in 22RV1 and VCaP cells
3 结论
实验通体外培养22RV1、VCaP、RWPE-1细胞,使用不同浓度的GEN处理48 h。利用CCK-8检测22RV1、VCaP、RWPE-1细胞增殖活力。结果显示GEN不抑制RWPE-1细胞的生长,GEN(50、100 μmol/L)能明显抑制22RV1、VCaP的生长。后续Western blot检测得出,GEN(50、100 μmol/L)浓度对前列腺癌标志性抗原PSA蛋白和AKR1C3的表达均有抑制作用。由于AKR1C3是雄激素合成过程中的关键酶,通过使用AKR1C3 siRNA、AKR1C3抑制剂(ASP-9521)分别与GEN单独/联合处理22RV1、VCaP细胞,推测GEN抗去势抵抗前列腺癌的生长是通过抑制AKR1C3的表达现实的。在前述实验中,GEN 50 μmol/L为首次出现抑制作用的浓度,所以选择GEN 50 μmol/L处理后续实验。Western blot检测AKR1C3的蛋白表达量,Western blot结果显示,GEN 50 μmol/L能够抑制细胞中AKR1C3蛋白的表达,且与AKR1C3 siRNA、AKR1C3抑制剂(ASP-9521)联合作用时,对AKR1C3的抑制效果更为显著。AKR1C3的抑制可延缓CRPC的发生、发展,增加患者的生存时间,为临床上使用植物雌激素治疗去势前列腺癌的治疗提供的新思路,研究结果为通过多食用豆制品预防前列腺癌提供了理论依据。
猜你喜欢
猪业科学(2022年8期)2022-11-22
保健与生活(2022年12期)2022-06-09
现代仪器与医疗(2021年6期)2022-01-18
中老年保健(2021年5期)2021-08-24
小雪花·成长指南(2021年2期)2021-05-20
世界科学技术-中医药现代化(2021年12期)2021-04-19
猪业科学(2020年11期)2020-12-17
天津医科大学学报(2019年3期)2019-08-13
初中生世界·九年级(2019年4期)2019-05-05
猪业科学(2019年10期)2019-01-08
