
洒金柏色素变化及光合作用响应特征
2021-11-13 09:01郭丽琴刘国彬姚砚武
中国农学通报 2021年29期
廖 婷,付 琳,郭丽琴,刘国彬,王 烨,姚砚武,曹 均
(1北京市农林科学院林业果树研究所,北京 100093;2内蒙古农业大学林学院,呼和浩特 010018)
0 引言
彩叶植物因其独特和丰富的色彩变化在园林绿化中具有较高的观赏价值,也是森林景观的重要组成材料。从园林植物造景角度,根据叶色和观赏期,可将彩叶植物分为春色叶植物、秋色叶植物、常色叶植物、双色叶植物和斑色叶植物5类[1-3]。洒金柏(Plantycladus orientaliscv.Semperourescens)是柏科(Cupressaceae)侧柏属侧柏(P.orientalis)的栽培变种,树姿优美,树冠外围叶片一年四季呈现不同程度的黄色,基部叶片始终为绿色,观赏期长,是重要的庭院、公园、景区常色叶观赏树种。探明其叶色动态变化的生理响应过程和机制对常色叶园林树种的选育具有重要意义。
植物叶片呈色与体内色素组成种类、相对含量及在叶片中的分布和定位有关[4];此外,外界环境因子,如光照强度、温度等对色素合成和显色有所影响[5-7]。叶片颜色发生变化最直接的原因是叶片中色素种类和比例的改变,对黄色叶植物来说,主要是叶绿素(Chl)和类胡萝卜素(Car)的比例发生了变化,与Chl和Car的生物合成与降解过程有关[8-9]。通常对各类色素的研究,大多以花瓣为材料进行[10-12]。近年来,对秋色叶树种在变色期的生理变化规律研究较多,如银杏、槭树、美国红枫、枫香、金叶榆、栎属植物等,主要包括光合特性、色素含量、相关酶活性、内含物质含量、糖代谢等方面[13-15],对常色叶树种的生理动态变化规律的研究较为罕见。而叶片中色素分布于质体膜上,叶绿素是光合作用的关键色素,其含量影响净光合作用,因此突变的黄色叶植株是研究叶绿体发育、叶绿素代谢、光合特性的理想材料。
基于此,本研究以洒金柏为研究材料,探究自然状态下不同月份黄叶的色素变化过程及光合响应特征,揭示黄叶形成的原因和叶色动态变化规律。同时,以相同枝条上的绿色叶作为对照,消除遗传背景不一致的影响。该研究可为洒金柏的遗传育种工作提供参考依据,对园林植物常色叶品种选育和推广具有重要的理论意义和实际应用价值。
1 材料与方法
1.1 试验材料
本试验所用洒金柏种植于北京市农林科学院林业果树研究所侧柏资源圃内,位于北京市海淀区,树龄20~30年,前期筛选出优良单株16株。洒金柏经过长期引种驯化,已经完全适应当地气候,通过对洒金柏的生长、株型、叶色、抗性等性状的长期观测和研究,从16个优良单株中选择顶部与基部叶片颜色差异最大的1株作为试验材料。
生理指标测定和采样时间为洒金柏叶片生长期,2018年3—10月,每个月取样1次,分别从植株4个不同方向上采集新鲜叶片,迅速装入冻存管,置于液氮中,随后冷藏在-80℃的超低温冰箱中,为后续试验做准备。
1.2 试验方法
1.2.1 叶色测定 用英国皇家园林协会(RHS)植物比色卡分别对洒金柏黄叶和绿叶每月进行比色并记录叶色变化,比色卡的颜色表达由数字和字母组成,前面的数字代表颜色,数值越大表示颜色越深;后面的字母则代表亮度,字母升序表示亮度越大。
1.2.2 色素含量测定 选取洒金柏不同时期顶部黄叶和基部绿叶进行相关色素含量测定,测定方法采用酒精萃取,具体参照蔡庆生植物生理学实验中的方法[16],用紫外可见分光光度计(型号UV-4802)测定波长665、649、470 nm下的吸光度,以95%乙醇作为空白对照,生物学重复3次。
叶绿素a(Chl a)、叶绿素b(Chl b)、Chl、Car的计算见公式(1)~(4)。
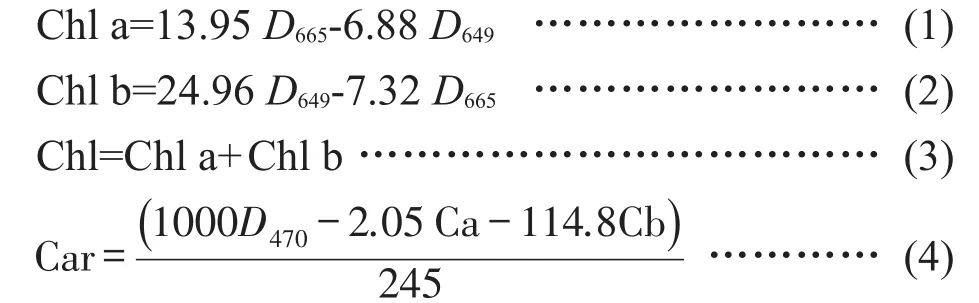
参考孔祥生等[17]的方法测定类黄酮含量。制作芦丁标准曲线后,以70%甲醇溶液作为提取剂,60%乙醇溶液作为空白对照,测定波长367 nm处的吸光度。
1.2.3 光合指标测定 光响应曲线测定:利用Li-6400便携式光合仪对洒金柏黄叶和绿叶分别进行光响应曲线测定,设定光合有效辐射(PAR)为0、50、100、150、200、400、600、800、1000、1200、1400、1600、2000 μmol/(m2·s),CO2浓度为 400 μmol/mol,相对湿度 60%~65%,流速500 μmol/mol。
光合特性测定:在晴朗无风的天气,上午8:30—11:30,使用Li-6400便携式光合仪对洒金柏顶部黄叶和基部绿叶进行光合作用测定,仪器设定的参数为PAR=1400 μmol/(m2·s),其余参数与光响应曲线参数一致。记录的光合参数包括净光合速率(net photosynthetic rate,Pn)、气孔导度 (stomatal conductance,Gs)、胞间 CO2浓度(intercellular carbon dioxide concentration,Ci)和蒸腾速率(transpiration rate,Tr)。水分利用效率(water use efficiency,WUE)通过公式(5)计算得出[18],每次测定包含3次生物学重复。

1.2.4 数据处理 运用SPSS 22软件对所测定的指标进行单因素方差分析,采用邓肯多重检验方法比较进行多重比较(α=0.05),通过皮尔森相关系数分析各指标之间的相关性。利用Excel进行绘图。
2 研究结果
2.1 洒金柏黄叶叶色变化
如图1和表1可知,洒金柏顶部叶片在3—10月间叶色呈现出不同程度的黄色或者黄绿色的动态变化。3月份新叶开始萌发,枝梢顶部长出嫩黄色的叶片,随着叶片的生长,枝梢顶部的叶色逐渐向黄绿色转变,6、7、8月份叶片偏向黄绿色,9月后叶片又逐渐变为黄色,10月份叶片呈现浓郁的金黄色。而绿叶只在8月颜色略微变深,其余月份绿叶颜色差异不大。
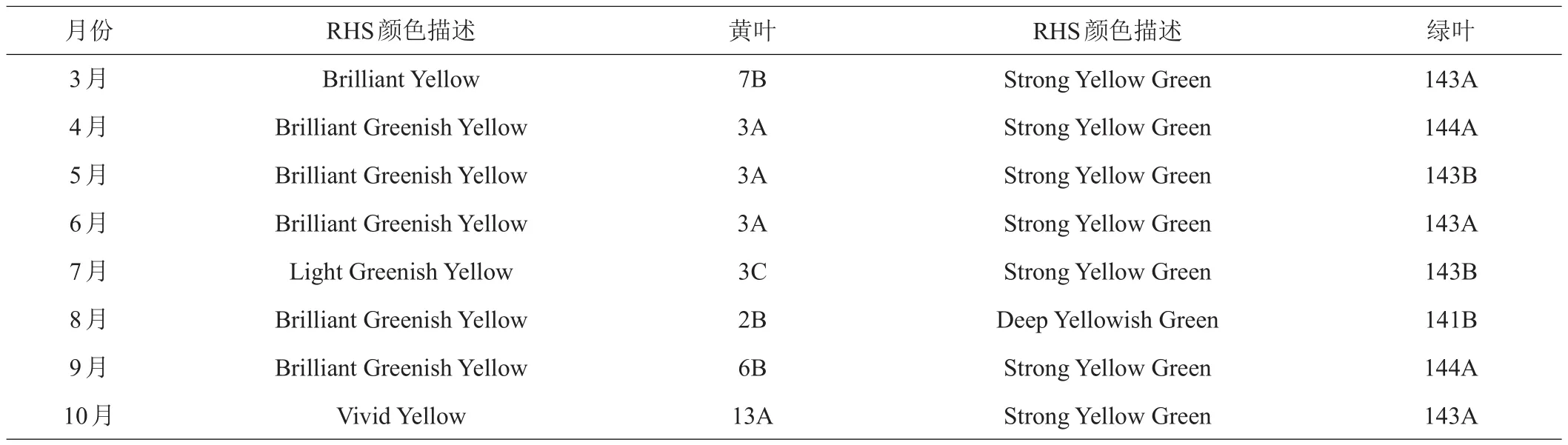
表1 洒金柏黄叶和绿叶RHS叶色

图1 3—10月洒金柏顶部黄叶叶色变化
2.2 洒金柏色素含量变化
如图2A所示,洒金柏顶部黄叶Chl a含量始终低于基部绿叶,且差异显著;黄叶与绿叶中Chl a含量的变化趋势基本类似,即从3月开始呈上升趋势,6—9月快速生长时期含量变化不大,仅绿叶Chl a含量在7月份有明显下降,9月后呈现下降趋势。黄叶和绿叶中Chl b含量变化趋势与Chl a趋势基本一致(图2B)。在黄叶中,虽然有小幅度的变动,但并不显著;在绿叶中,3月、4月含量显著低于5—10月含量。黄叶与绿叶中的Chl含量变化趋势与Chl a含量变化趋势基本一致(图2C),黄叶的Chl含量每个月均显著低于绿叶,5—9月具有最高的Chl含量水平。
Car含量在3—4月份出现1个快速下降的阶段,在黄叶中下降了51%,4—8月份虽平稳上升,但未达到显著水平,8月份后开始大幅度上升(图2D)。8月份,黄叶中Car含量为0.074 mg/g,10月达到0.168mg/g,上升幅度达56%。黄叶中Car含量在各个时期均低于绿叶中含量。

图2 洒金柏3—10月叶色变化过程中色素含量
黄叶和绿叶中类黄酮含量变化趋势基本一致,呈现平稳变化到迅速下降的过程(图2E)。3—7月,黄叶中类黄酮含量显著低于绿叶;7月后,黄叶中类黄酮含量呈平稳下降,而在绿叶中有1个迅速下降的过程。8—10月,两种叶片中类黄酮含量无显著差异。
虽然黄叶中各色素含量比绿叶中更低,但黄叶中类胡萝卜素与总叶绿素(Car/Chl)含量比值高于绿叶(图2F),6—9月无显著差异。3月,黄叶与绿叶中Car/Chl含量比值差异最大,黄叶中达到最大值,为1,绿叶中比值为0.26。
2.3 洒金柏光合参数动态变化
图3A为洒金柏黄叶和绿叶的光响应曲线。如图所示,在PAR<200 μmol/(m2·s)的低光照条件下,Pn对光合有效辐射的响应较为敏感,Pn在绿叶中的增加幅度比黄叶更大,随后Pn在PAR为200~1200 μmol/(m2·s)范围内平稳上升。当PAR>1200 μmol/(m2·s)时,黄叶Pn趋于平稳,而绿叶Pn在PAR>1400 μmol/(m2·s)时趋于平稳,之后随PAR的增加而略微降低,表现出光抑制现象。在相同的PAR条件下,绿叶的Pn始终显著高于黄叶。
根据光响应曲线的结果,选择光饱和情况下(PAR=1400 μmol/(m2·s))测定黄叶和绿叶生长期不同月份的光合气体交换参数。由图3B可知,两种叶片的Pn随月份增加均呈现先升高后降低的趋势,绿叶在8月出现最大值,为13.04 μmol/(m2·s),黄叶在7—9月的Pn差异不显著,最大值为11.11 μmol/(m2·s);3—8月,洒金柏黄叶的Pn始终显著低于绿叶,9—10月无显著差异。
Gs整体表现出与Pn一致的趋势(图3C),黄叶在5月有1个降低的过程,绿叶在4月有1个降低的过程,8、9月,黄叶的Gs显著高于绿叶。黄叶和绿叶均在8月份有最大的Gs值,分别为0.293 mol/(m2·s)和0.242 mol/(m2·s)。Ci的变化趋势与Gs基本一致,随着时间的增加呈现出先升高后降低的趋势。但黄叶在4—5月有1个波动的变化过程(图3D)。
由图3E可知,Tr整体呈现先升高后降低的趋势,黄叶Tr最大的月份为8月,达4.95 mmol/(m2·s);绿叶7月份Tr最大,为4.04 mmol/(m2·s)。两种叶片Tr在9—10月份无显著差异。在整个生长季中,黄叶的WUE低于绿叶,3—8月变化趋势较为平缓,只在5月时出现较大波动(图3F);8—10月,WUE随时间而显著升高。
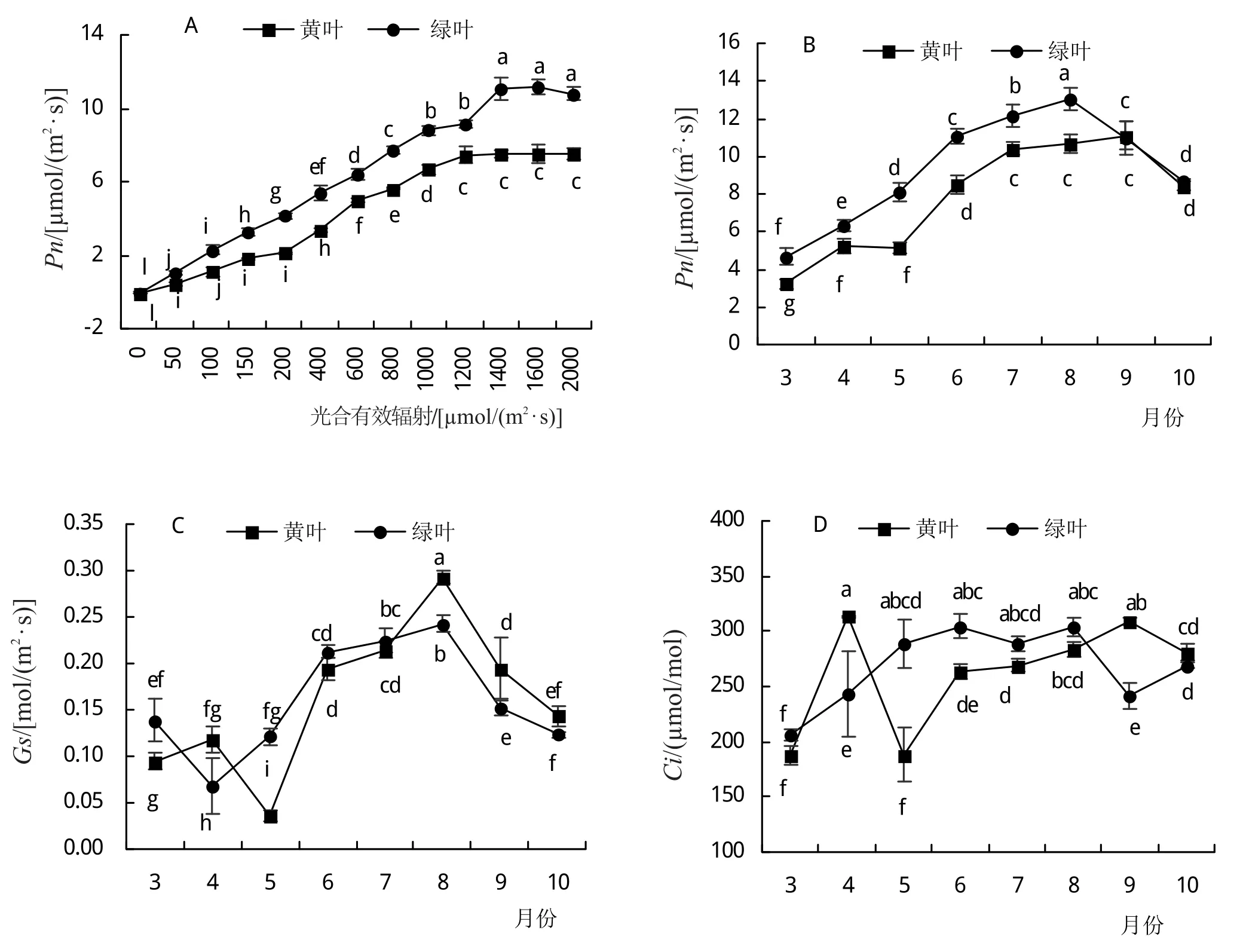
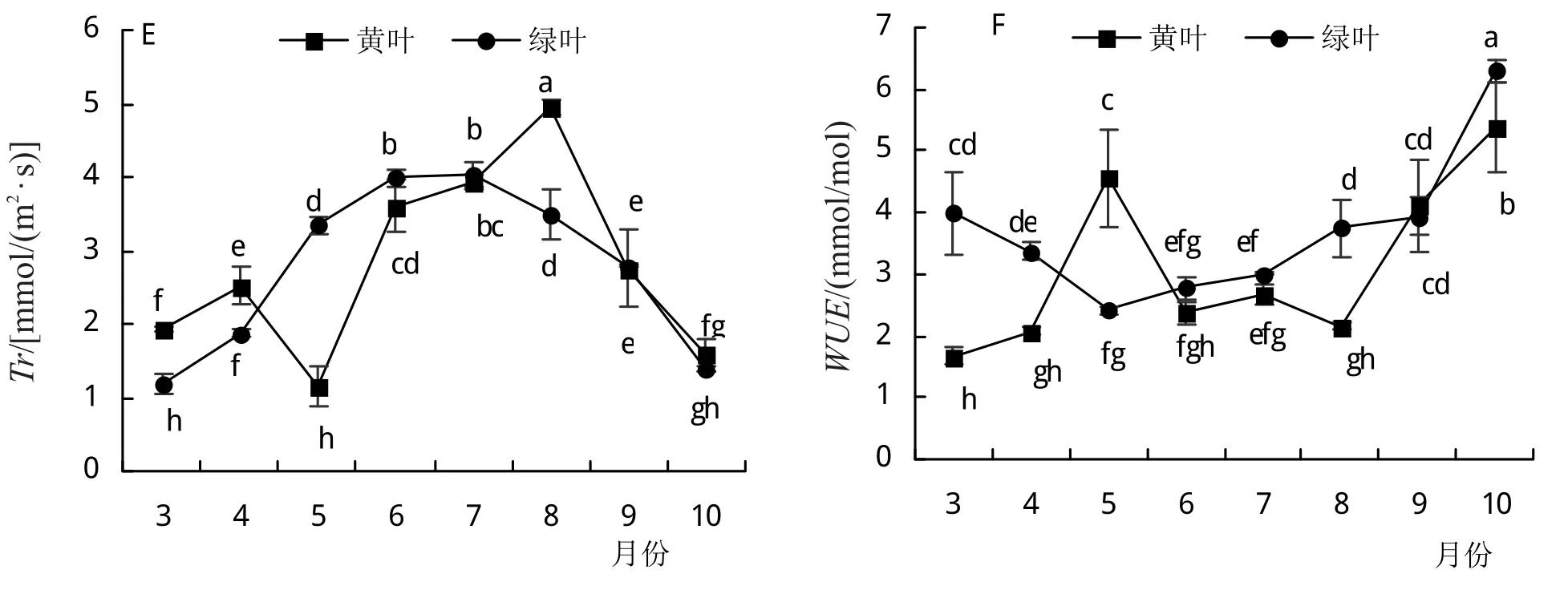
图3 洒金柏叶色变化过程中光合作用
2.4 洒金柏光合参数与色素含量相关性分析
由表2可知,黄叶与绿叶中,色素含量指标Chl a、Chl b、Chl之间呈极显著正相关性,4个光合参数指标之间也呈显著或极显著正相关。黄叶中Gs和Tr的相关性最大,相关系数达0.917;其次是Pn和Gs,相关系数为0.82。Tr与WUE呈极显著负相关关系,黄叶中相关系数为-0.559,绿叶的相关系数为-0.668,表明蒸腾作用越强,WUE越低。
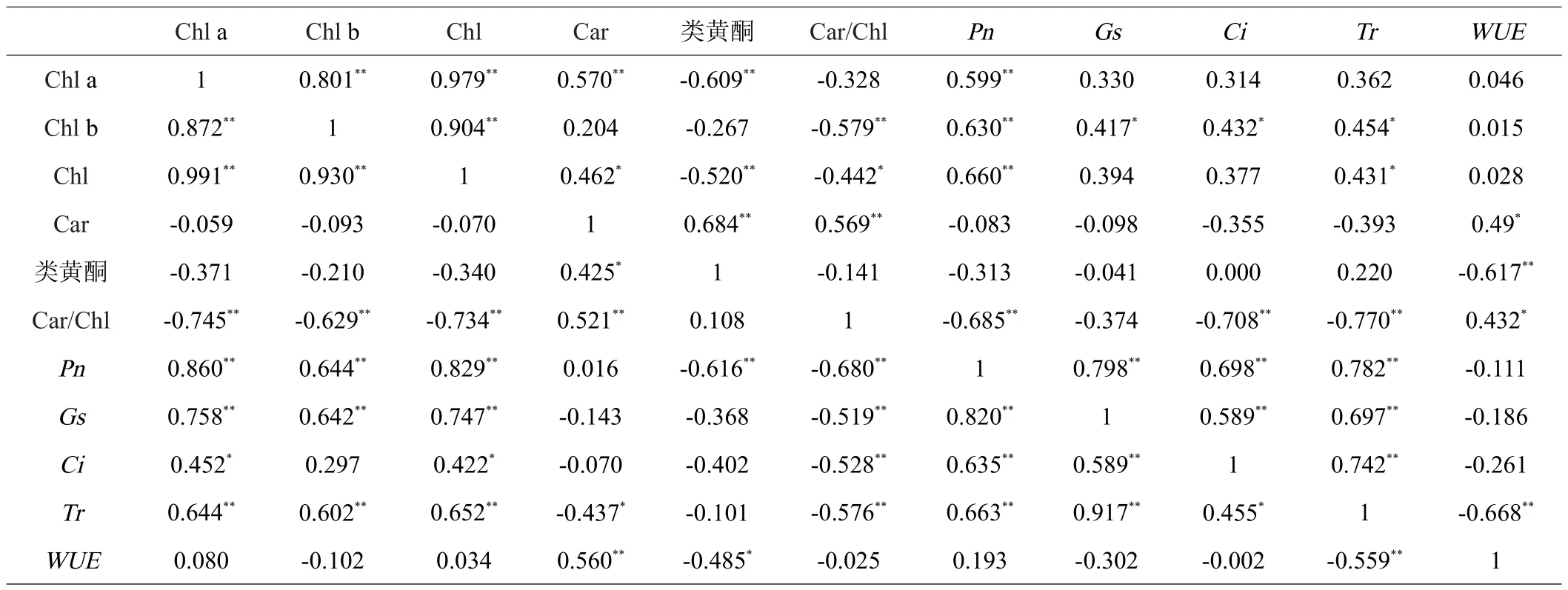
表2 洒金柏色素含量与光合参数相关性
在黄叶中,Car/Chl与Chl a、Chl b、Chl含量之间呈极显著负相关性,且相关系数均大于绿叶中的相关系数,表明黄叶受Car/Chl影响可能更大。此外,黄叶中Chl含量、Chl a含量与光合参数指标之间呈显著正相关性,与Car/Chl呈显著负相关性,相关系数大于绿叶中的相关系数,表明黄叶中光合作用受色素含量影响较大,绿叶光合作用除了受色素含量直接影响外,可能与其体内羧化酶活性有很大关系。
3 结论
本研究采用表型观测与生理测定相结合方法研究了不同月份洒金柏叶色变化、叶片色素含量与光合作用的响应特征。结果表明,3月黄叶的Car/Chl含量比值达到最大值为1,此时绿叶的比值为0.26;黄叶的光合参数与Chl含量之间呈显著正相关性,最大达0.86,而与Car/Chl含量比值呈显著负相关性,达-0.68。表明洒金柏叶片呈现黄色是由于黄叶中Car/Chl含量比值较绿叶中更高导致。
4 讨论
Chl、Car和花色苷等色素种类和比例的变化是彩叶植物叶色多变的直接原因。在洒金柏中,黄叶中的Chl a、Chl b、Chl、Car含量在3—10月均显著低于绿叶中含量,呈显著正相关关系;而黄叶中Car/Chl比值高于绿叶,且变化趋势与黄叶叶色变化趋势相一致,即比值越大,叶色越黄,比值越小,叶色越绿,表明洒金柏呈现黄色可能是由Car/Chl比值较高导致。金叶植物叶片呈色原因在银杏‘万年金’中有相关研究,认为Car/Chl和Car/Chl a比值偏高可能是子代无性系中叶片呈现黄色的主要原因之一[19]。对金叶白蜡的研究也发现,其叶片呈现黄色的内在原因是Chl含量低于对照白蜡,而Car含量高于对照白蜡[20]。与金叶白蜡类似的是,本研究中洒金柏在夏季也出现略微反绿的现象,观赏价值降低,相比春季,夏季洒金柏中黄叶的Car含量显著降低,Chl含量升高,这可能是由于夏季高温、强光照等环境因子不利于Car合成和积累导致。
在有的植物中,黄叶形成与类黄酮的含量紧密相关,类黄酮物质会在植物的叶片、花瓣和果实中大量的积累,随着类黄酮物质含量的变化植物的叶色也会随之改变[21]。本研究中,3—7月,绿叶中类黄酮含量显著高于黄叶,这与杨瞬博对银杏的研究结果相符[22]。但8—10月,类黄酮含量在黄叶中与3—7月含量无差异,绿叶中类黄酮含量显著低于3—7月含量,黄叶与绿叶含量无差异,表明秋季温度的降低并没有促进黄酮物质的积累,因此推测尽管洒金柏中类黄酮绝对含量较高,但类黄酮不是洒金柏黄叶的主要显色物质。
植物叶片气体交换参数是衡量光合特征的重要指标,Chl含量的高低一定程度上能够反应光合作用高低。通过洒金柏的光响应曲线可知,在相同光强条件下,黄叶的Pn显著小于绿叶;黄叶在PAR>1200 μmol/(m2·s)后,Pn无显著差异,达到饱和状态,而绿叶在PAR>1400 μmol/(m2·s)后,Pn达到饱和。这与倪妍妍等[23]对侧柏光响应曲线研究结果基本相符,在侧柏中,5年生接穗的光饱和点最高,达(1824±148)μmol/(m2·s)。但洒金柏中,在PAR<200 μmol/(m2·s)时,Pn的升高幅度较快;当PAR在200~1200 μmol/(m2·s)之间,Pn基本呈匀速上升状态,随后保持平稳。这与对侧柏光响应曲线的研究略微差异。
金色叶植物一般生长量小,光合能力和比叶质量均显著低于相应的绿叶植株[24-25]。如对金叶女贞的研究发现,其光合能力低于小叶女贞,Car/Chl比值则高于小叶女贞[24];中华金叶榆是普通白榆的天然黄叶突变体,黄叶苗生长缓慢,净光合速率显著低于绿叶苗[25];对于金叶国槐的研究发现同一时期普通国槐的净光合速率高于金叶国槐[26]。本研究对洒金柏不同月份光合作用的研究表明,光合气体交换参数基本呈先上升后降低的趋势,且光合指标与色素含量指标之间呈显著正相关性,这与多数研究结果相符。3—8月,黄叶的Pn显著低于绿叶,虽然Tr也低于绿叶,但最终WUE低于绿叶,表明绿叶在生长过程中对水分的利用效率更高。8月之后,叶片趋向成熟,而此时黄叶和绿叶的WUE均显著高于夏季,说明洒金柏秋冬季节对水分有较高的利用效率。这从侧面反映可能其抗旱能力较强,在秋冬季水分条件较差情况下,能够通过调节自身生理系统来满足生长需求,这对于北方缺水城市来说至关重要。
猜你喜欢
今日农业(2022年2期)2022-11-16
保健与生活(2021年13期)2021-07-14
语文世界(小学版)(2021年5期)2021-06-04
中国果树(2020年2期)2020-07-25
岭南音乐(2019年6期)2019-12-31
岭南音乐(2019年5期)2019-10-29
中华诗词(2019年12期)2019-09-21
山东林业科技(2019年2期)2019-06-03
天然产物研究与开发(2018年11期)2018-11-30
天然产物研究与开发(2018年4期)2018-05-07
