
甘草生长素反应因子(ARF)基因家族的鉴定及表达分析
2021-11-13 09:01:40关思静徐蓉蓉葛甜甜颜永刚刘阿萍程萌格
中国农学通报 2021年29期
关思静,高 静,徐蓉蓉,葛甜甜,王 楠,颜永刚,张 岗,陈 莹,刘阿萍,程萌格
(1陕西中医药大学药学院/陕西省秦岭中草药应用开发工程技术研究中心,陕西西安 712046;2陕西中医药大学/陕西中药资源产业化省部共建协同创新中心,陕西咸阳 712083)
0 引言
生长素(auxin)是现已报道发现最早、研究最深入的植物激素,在胚胎发生、顶端优势、维管伸长、开花、果实发育和侧根发生等多个生长发育过程中起着中心作用。分子水平分析发现,生长素通过一定的基因控制和应答机制实现对植物生长调控作用,早期的应答基因主要包括Aux/IAA家族、GH3家族和受生长素调控的RNA家族[1]。生长素反应因子(ARFs),作为植物生长中的关键转录因子,其主要功能为在上游区获取生长素信号后,寻找位于下游区的生长素响应基因的启动子序列,与其中的生长元素响应元件相结合,激活或抑制生长素应答基因的表达,进而实现对植物生长发育的控制[2]。研究表明,绝大多数ARF蛋白具有3个保守结构区域,N端为植物特有的B3型DNA结合结构域,该结构域保守性极强,能直接与生长素响应基因启动子区域的生长素应答元件(AuxRE)结合;中央ARF功能域,其氨基酸组成决定了ARF蛋白是否起激活或抑制作用;以及C端Aux/IAA结构域,具有决定ARF蛋白间的同源聚集或ARF蛋白与生长素应答基因间的异源聚集功能,并用于确定ARF的聚集状态[3]。
目前,模式植物拟南芥基因组中共23个ARF转录因子被鉴定[4],其中部分成员的功能已得到验证。分析结果表明,AtARF1与AtARF2具有一些共同的功能,可能正向调控植物叶片衰老的发生以及花器官的脱落[5-6];Kelley等[7]研究发现AtARF3与KANADI蛋白质发生物理相互作用,形成蛋白复合物,构成生长素依赖性调节模块的一部分,是拟南芥中被膜发育和叶极性建成所必需的功能复合物;AtARF2-4和AtARF5之间有重叠和非冗余的功能,并且对拟南芥雌雄配子体的发育至关重要[8];拟南芥生长素转录因子AtARF6、AtARF8和AtARF17在调节雌蕊和雄蕊的发育,果实发育、受精,花药发育和花粉形成中起到重要作用[9-11];AtARF7/AtARF19、AtARF11、AtARF10与AtARF16等转录因子则直接或间接介导了拟南芥侧根的形成[12-15]。
乌拉尔甘草(GlycyrrhizauralensisFisch.)是豆科甘草属植物,其根茎是极为重要的常用中药材。目前甘草的全基因组测序已经完成,这为研究相关基因的功能提供了基础[16]。本研究利用生物信息学手段对甘草ARF基因家族进行分析鉴定,对其编码蛋白的理化性质、保守结构域及保守基序、基因结构、顺式作用元件、系统进化关系等进行了预测和分析,并利用转录组数据研究了不同胁迫条件下的基因表达模式,旨在为深入研究甘草抗逆机理研究提供参考。
1 材料与方法
1.1 材料及处理
实验于2019年10—12月在陕西中医药大学陕西中药资源产业化省部共建协同创新中心组织培养室进行。甘草种子消毒灭菌后,在恒温气候箱中催芽1周,挑选长势一致的幼苗移至霍格兰营养液中,放置在培养室水培1月。分别采用15 g/L聚乙二醇(PEG-6000)、150 mmol/L氯化钠(NaCl)、10 μmol/L磷酸二氢钾进行干旱、高盐以及低磷胁迫处理。对照(CK)用营养液培养。在胁迫处理1周后采集甘草幼苗茎叶和根,每个处理3个生物学重复,委托杭州景杰生物科技有限公司进行转录组测序。上述处理的甘草种子和幼苗在气候箱与培养室中的培养条件温度为(25±2)℃,湿度为60%~70%,光周期为12 h光/12 h暗。
1.2 方法
1.2.1 甘草ARF基因的鉴定及序列分析 模式植物拟南芥的ARF序列下载自http://www.UniProt.org/,甘草的全基因组序列数据获取自http://ngs-data-archive.psc.riken.jp/Gur-genome/。以拟南芥23个ARF家族成员的蛋白序列作为查询序列,运行TBtools工具的本地检索程序,筛选阈值设为1×10-5,初步获得候选序列。将候选ARF蛋白序列提交到NCBI,进行在线blastp比对。去除重复序列后,利用在线网站SMART(http://smart.embl-heidelberg.de/)和 Pfam(https://pfam.xfam.org/)对筛选的甘草ARF蛋白进行结构域验证,并删除不含ARF蛋白特征结构域的基因。最后利用ExPASY-ProtParam(https://web.expasy.org/protparam/)对甘草ARF蛋白序列进行氨基酸数目、分子量、等电点等理化性质预测,同时借助Cell-PLoc2.0(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/)进行蛋白亚细胞定位预测。
1.2.2 甘草ARF基因的结构分析 从甘草全基因组序列信息中检索出甘草ARF基因外显子-内含子位置信息,利用在线网站GSDS2.0(http://gsds.cbi.pku.edu.cn/)分析甘草ARF基因完整序列得到外显子-内含子结构。
1.2.3 甘草ARF家族保守基序分析 利用MEME 5.1.1在线工具(http://meme-suite.org/tools/meme)分析甘草ARF蛋白的保守基序,最大基序检索数值设为10,最适基序长度为6~200个氨基酸。
1.2.4 甘草ARF启动子顺式作用元件分析 提取甘草ARF基因上游2 kb的序列信息,使用在线工具PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)预测基因顺式作用元件。
1.2.5 甘草ARF系统进化分析 利用多重序列比对软件MAFFT 7.0对甘草、拟南芥ARF蛋白进行序列比对计算,比对结果进一步通过MEGA 7.0软件采用邻接法构建系统进化树,其中校验参数设置为1000,其余均为默认参数。
1.2.6 甘草ARF基因的表达模式分析 利用转录组测序数据分析甘草ARF基因的表达特征。转录组数据包含对照、甘草干旱、高盐以及低磷胁迫处理的基因表达数据,以不同实验条件下的RPKM值为表达水平,通过对数据进行均一化处理,利用TBtools的HeatMap程序做热图分析[17]。
2 结果与分析
2.1 甘草ARF基因家族的鉴定
利用本地blast比对检索与在线blastp比对,得到初步候选序列,并进一步利用Pfam和SMART进行结构域验证,去除不含有ARF保守结构域或结构域不完整的序列,最终鉴定出10个甘草ARF基因序列,将它们命名为GuARF1~GuARF10(表1)。其中除GuARF8和GuARF10含有B3、Auxin_resp和Aux/IAA结构域外,其余8个GuARF蛋白只含有B3和Auxin_resp结构域(图1)。分析甘草10个ARF蛋白序列,发现编码ARF蛋白的氨基酸长度在301~945 aa之间,分子量范围为33.07~104.37 kDa,等电点范围为5.93~8.50,其中6个GuARF等电点小于7,4个GuARF等电点大于7,且平均等电点小于7,表明GuARF为弱酸性,推测可能在酸性亚细胞环境中发挥作用。不稳定系数分析结果表明10个GuARF的不稳定指数均大于40,为不稳定蛋白。亚细胞定位预测表明所有的GuARF均定位在细胞核(表1)。
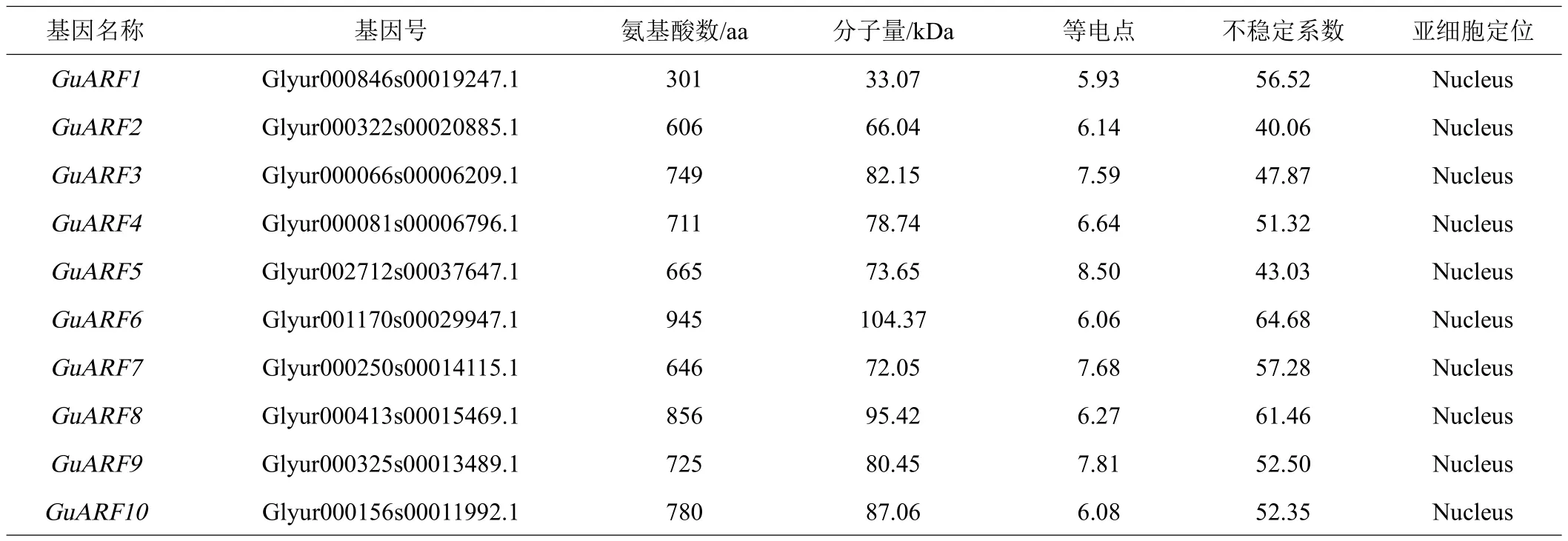
表1 甘草ARF基因家族信息
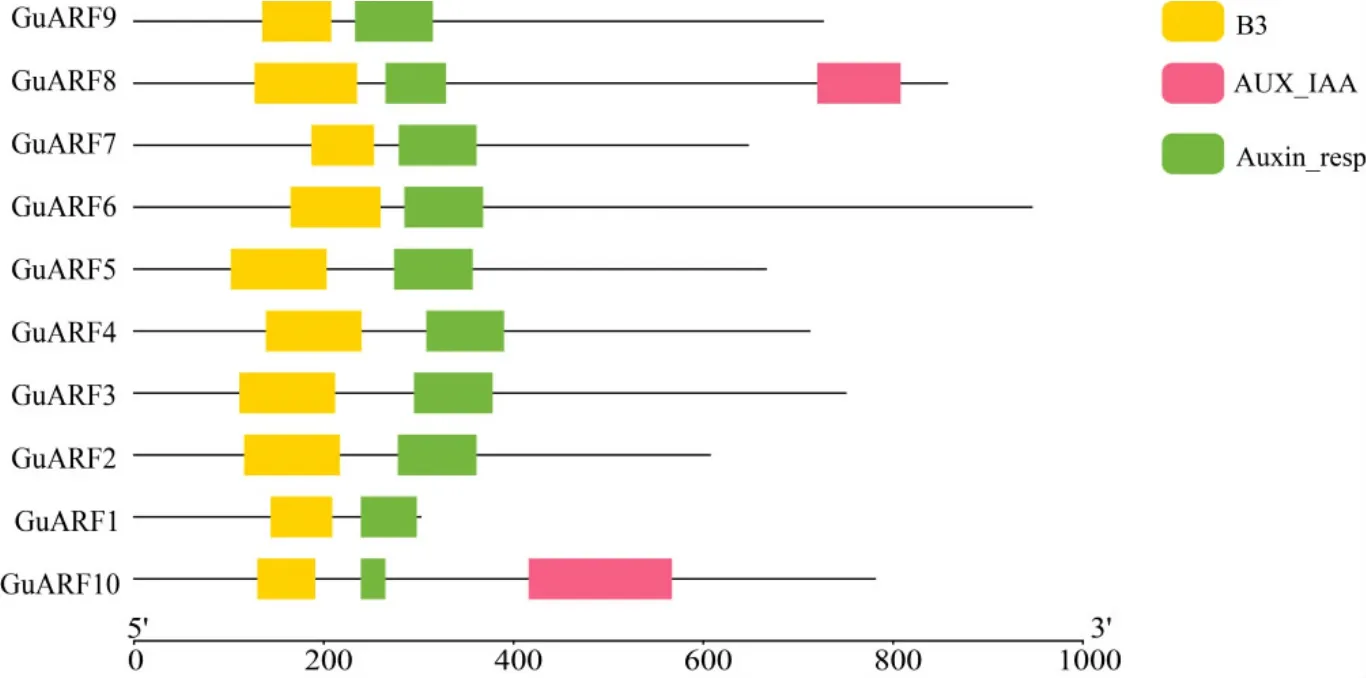
图1 GuARF基因家族蛋白保守结构域
2.2 甘草ARF基因的结构和系统进化分析
为了进一步了解甘草ARF基因的特征,使用在线工具GSDS2.0对甘草ARF基因结构进行分析(图2),结果表明ARF基因家族大多数成员的结构比较复杂,含有外显子数量6个(GuARF2、GuARF4、GuARF5)到22个(GuARF22)不等。系统进化分析表明,位于同一亚家族的GuARF基因具有相似的外显子-内含子分布模式(图2)。

图2 GuARF基因家族内含子-外显子结构
利用MEME在线数据库对甘草ARF蛋白进行保守基序分析,共得到10个保守基序(图3)。10个GuARF蛋白中均含有motif 4、motif 5和motif 7,除此之外,8个GuARF蛋白含有motif 1和motif 2,7个GuARF蛋白含有motif 8,4个GuARF蛋白含有motif 3、motif 9和motif 10,3个GuARF蛋白含有motif 6。其中GuARF3、GuARF4和GuARF5含有8个基序,GuARF1、GuARF2、GuARF6、GuARF7、GuARF8、GuARF9含有7个基序,GuARF10只含有4个基序。结果表明,出现频率较高的motif为GuARF蛋白结构域中非常重要的保守基序。

图3 GuARF蛋白保守基序分析
2.3 甘草ARF基因顺式作用元件预测
提取甘草ARF基因上游2 kb的序列进行顺式作用元件分析,结果如图4所示,甘草ARF基因上游存在5类顺式作用元件:①光响应类相关元件,如G-box元件、Box4元件等;②植物激素响应相关元件,如响应脱落酸代谢的ABRE元件、响应生长素代谢的TGA-element等;③逆境胁迫响应类元件,如干旱胁迫响应元件MBS、低温胁迫响应元件LTR等;④植物生长发育响应元件,如调节栅栏叶肉细胞分化的HD-Zip 1元件、分生组织表达相关的CTA-box等;⑤次级代谢产物合成类响应元件,类黄酮生物合成基因调控元件MBSI;其中逆境胁迫响应类元件占比20%。并在GuARF1和GuARF5的启动子区域检测到3个生长素响应原件(TGA-element)。
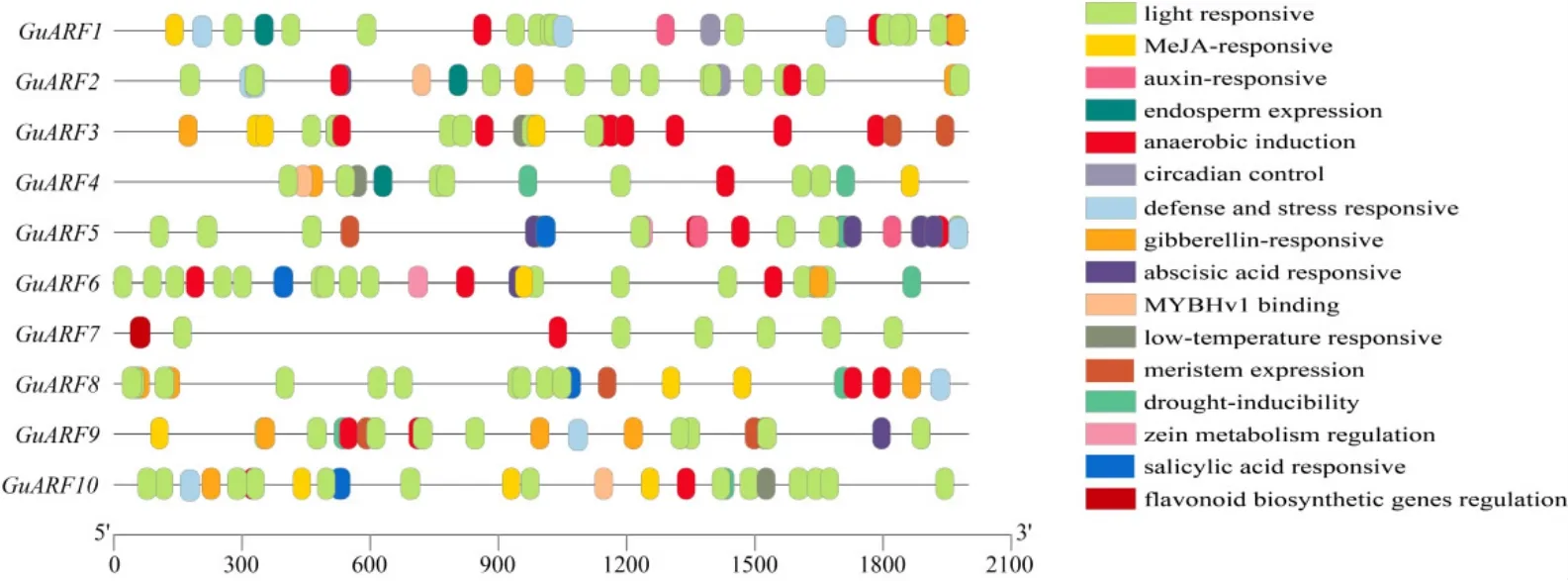
图4 GuARF基因顺式作用元件分析
2.4 甘草ARF系统进化分析
为了进一步了解甘草ARF基因家族成员的进化关系,构建了甘草、拟南芥基因家族系统进化树(图5)。系统发育分析表明甘草和拟南芥共33个ARF蛋白可以分为I、II、III三个家族,其中I家族可以进一步分为Ia和Ib两个亚家族。GuARF7和GuARF10属于Class Ia;Class Ib仅为拟南芥 ARF 蛋白;GuARF1、GuARF6、GuARF8和GuARF9属于Class II;GuARF2、GuARF3、GuARF4、GuARF5属于Class III。其中ARF蛋白的编码基因在Class Ia、Class II和Class III各自包含一个自展值为99的直系同源基因对(GuARF9/AtARF4、GuARF2/AtARF17、GuARF7/AtARF1);此外,从图5中可以看出在Class II和Class III中甘草还含有两个旁系同源基因对(GuARF1/GuARF6、GuARF3/GuARF5)。
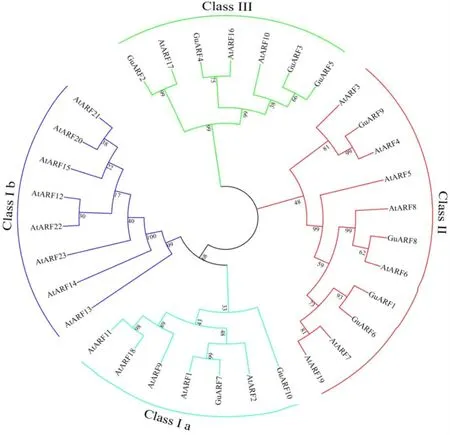
图5 甘草和拟南芥ARF蛋白系统进化树
2.5 甘草ARF基因家族的表达分析
利用甘草在干旱、高盐和低磷胁迫的RNA-Seq转录组数据,得到10个GuARF基因对应转录本的RPKM值,做对数值转换,利用TBtools的HeatMap程序生成热图(图6)。结果显示,对照处理中GuARF5与GuARF6在茎叶中大量表达,而在根中表达量很低;GuARF2、GuARF3、GuARF8、GuARF9在根中均有表达,且GuARF9的表达量很高,而在茎叶中均未见表达或表达量很低,其余GuARF基因在所检测的组织中表达量很低或未检测到。
茎叶和根中的GuARF在不同逆境胁迫下的表达呈现差异性。如图6-A所示,茎叶中的GuARF基因其表达模式可以分为3类,第1类包含3个GuARF基因(GuARF2、GuARF1、GuARF10),在低磷胁迫下大量表达,在对照和其他胁迫处理下表达量低或未见表达;第2类包含2个GuARF基因(GuARF5、GuARF6),仅在对照中具有较高的表达量;剩下5个GuARF基因在干旱胁迫或盐胁迫处理下表达量明显上调,其中GuARF9在干旱胁迫处理中大量表达,GuARF4在盐胁迫处理中大量表达,GuARF7、GuARF8、GuARF3在干旱和盐胁迫处理中均上调表达。
胁迫处理下GuARF基因在根中的表达模式可以分为两类(图6-B),每一类都包含5个GuARF基因,第1类中,除GuARF9在盐胁迫处理下表达量没有明显变化,其余基因在对照和盐胁迫处理中均上调表达,而在干旱胁迫和低磷胁迫处理中表达量很低或下调。第2类中5个GuARF基因在盐胁迫中表达量均上调,除GuARF6在干旱胁迫中上调表达,其余基因在对照、低磷和干旱胁迫处理中表达量很低或下调。
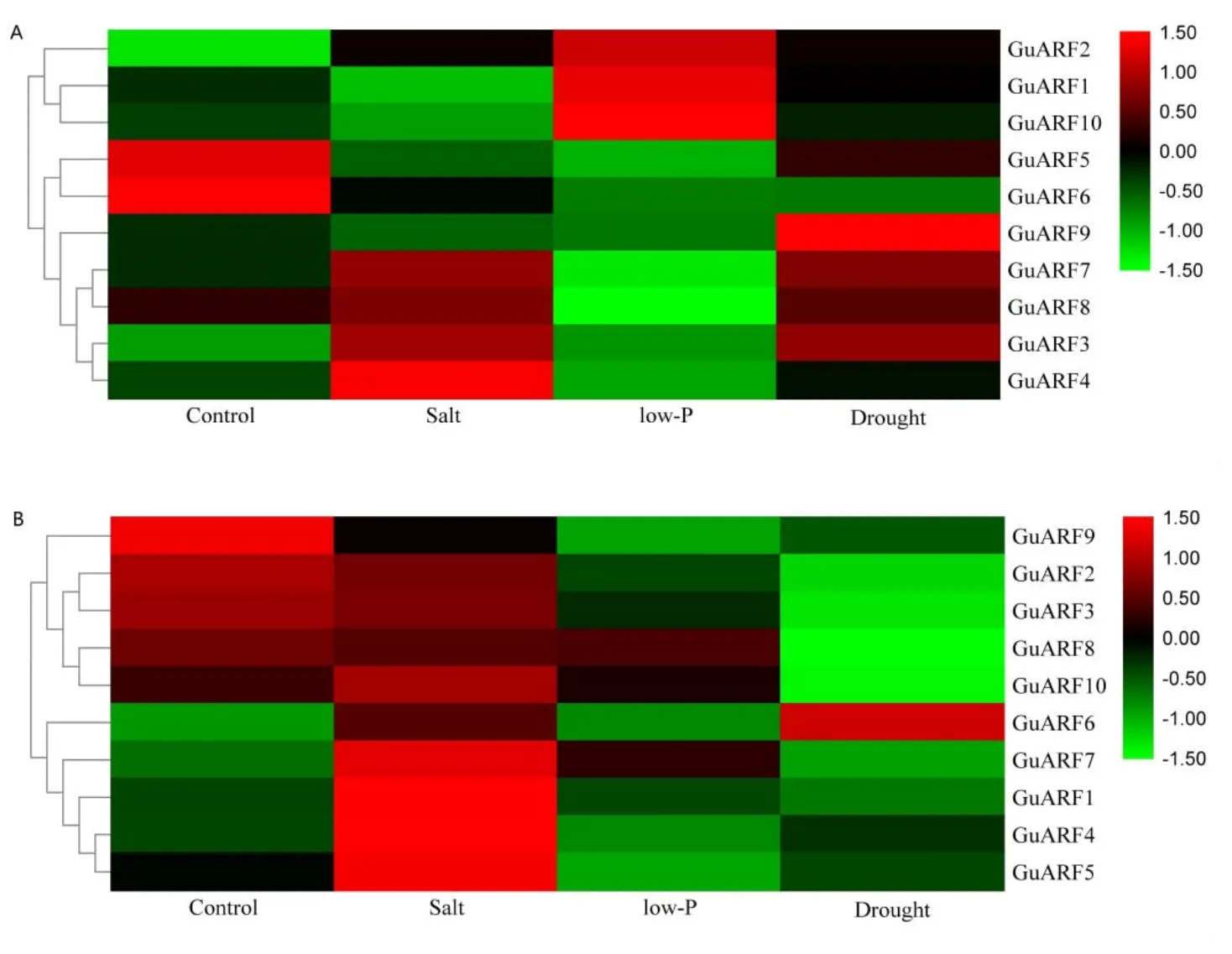
图6 非生物胁迫下GuARF基因在甘草茎叶(A)和根(B)中的表达
3 讨论与结论
近年来,随着生物信息技术的不断进步,越来越多的植物全基因组测序完成。本研究基于甘草全基因组序列数据,最终鉴定出10个ARF基因,比已鉴定出的拟南芥(23个)、水稻(25个)、番茄(22个)和紫苜蓿(24个)的ARF基因家族成员数目少[4,17-19]。保守结构域分析结果表明10个ARF蛋白均含有保守的B3结构域和Auxin_resp结构域,GuARF8和GuARF10含有Aux/IAA结构域。为了进一步研究甘草与拟南芥的ARF同源进化关系,构建了系统进化树,结果显示Class Ib均为拟南芥ARF蛋白,这与其他植物番茄[18]、谷子[20]、鹰嘴豆[21]中的Ib家族类似,这些成员包含拟南芥ARF假基因(ARF13)和7个串联重复基因(AtARF12-AtARF15和AtARF20-AtARF22)[22]。除Ib家族外,大多数甘草ARF蛋白和其他植物一样,与拟南芥ARF蛋白存在对应关系,表明这些蛋白可能与拟南芥ARF蛋白具有相似的功能。此外,甘草ARF基因家族中还含有旁系同源基因,推测在该基因家族进化过程中,可能出现了基因复制事件。
ARF基因具有较高的保守性,具有相同或相似功能的基因可能聚为一类,为预测该基因家族的功能提供了参考。作为模式植物的拟南芥,目前在基因研究方面研究较为深入,通过基因序列比对与聚类分析,推测与拟南芥ARF基因高度同源、处于较近分支的甘草ARF基因其功能与拟南芥中发挥的作用类似。根据AtARF4的功能推测其同源基因GuARF9在器官发育和营养生长中发挥重要作用[23];GuARF2(AtARF17)可能在miR160的调控下表达,在花药发育和花粉形成过程中的功能至关重要[11];而Class III中与AtARF10和AtARF16处于较近分支的GuARF3和GuARF4,可能共同调控侧根的形成[15]。对甘草ARF基因在不同组织的表达分析中发现,7个GuARF基因在茎叶中或根中均有表达,但存在明显的差异,表现出组织表达特异性,这可能与基因功能具有一定的联系。在甘草幼苗的正常发育过程中,GuARF1、GuARF4与GuARF7在茎叶与根的生长发育过程中几乎没有表达,推测这些基因可能在甘草的某个特定发育时期或组织中发挥作用。而Class III中的基因在茎叶与根中都有一定的表达(除GuARF4)。甘草ARF基因在不同组织和发育时期的表达差异以及功能有待进一步的研究与实验验证。
生长素还参与植物对生物和非生物胁迫的反应[24]。为了进一步了解甘草ARF基因家族在非生物胁迫下的反应,分析了候选的10个甘草ARF基因在不同胁迫处理下的相对表达量(图6)。结果表明,在高盐胁迫下,茎叶和根中的ARF基因都具有明显的变化,尤以Class III中ARF基因表现敏感。低磷胁迫下茎叶中的GuARF1、GuARF2、GuARF10上调表达明显,推测这些基因可能在甘草低磷胁迫下具有正向协同作用。干旱胁迫处理下,GuARF的基因表达量差异较大,茎叶中的GuARF9、GuARF7、GuARF3上调表达明显,而根中除GuARF6表达量较高其余均明显下调表达。在拟南芥干旱响应基因的鉴定中,一些编码生长素反应蛋白的基因被鉴定为干旱下调基因,这表明生长素可能对干旱胁迫信号产生负调控[25]。另一方面,也有研究结果表明,干旱和高盐度胁迫均诱导了编码生长素响应蛋白的基因上调表达[26];例如有研究发现,生长素应答基因(IAA18)在干旱胁迫下被诱导上调[27-28]。因此,植物ARF的时空表达模式较为复杂,且存在物种表达特异性[1]。
本研究从甘草基因组鉴定出10个ARF基因,可分为3大家族,并综合分析了甘草ARF基因的系统发育关系、保守基序、外显子/内含子结构和非生物胁迫下的表达谱。结果表明GuARF基因在非生物胁迫下呈现复杂多样的表达模式,相对于干旱和低磷胁迫,高盐胁迫下的甘草ARF基因表现更为敏感。虽然利用生物信息学方法对甘草ARF基因家族做了较为系统的分析,但由于分析手段本身存在不同设置参数的影响,且缺乏实验部分的验证,其响应高盐、低磷与干旱胁迫的分子机制还需要深入的探讨。
猜你喜欢
新医学(2023年10期)2023-12-09 15:04:51
中学生天地(A版)(2023年1期)2023-02-17 00:33:04
南方医科大学学报(2022年3期)2022-04-13 01:51:26
浙江大学学报(农业与生命科学版)(2021年3期)2021-07-10 07:07:36
生物学通报(2021年4期)2021-03-16 05:41:34
科学大众(2020年12期)2020-08-13 03:22:38
乡村科技(2019年9期)2019-06-26 03:52:04
数码世界(2018年7期)2018-12-24 15:06:05
生命科学研究(2018年1期)2018-05-29 01:12:47
上海农业学报(2017年3期)2017-04-10 12:39:14
