
管涔山林下草本层植物对模拟增温的响应
2021-11-10 08:28徐满厚杨晓辉杨晓艳于秀立
中国野生植物资源 2021年10期
徐满厚,杜 荣,杨晓辉,杨晓艳,于秀立
(太原师范学院地理科学学院,山西晋中030619)
全球气候变暖已成为毋庸置疑的事实,全球表面平均温度在过去一个世纪已上升了0.6℃[1],陆地生态系统的结构和功能深受影响[2]。作为当今生态学与环境科学领域研究的热点问题,植物群落特征及其地上生物量的变化对陆地生态系统的结构和功能极为重要[3-4]。因此,研究植物群落盖度及其地上生物量变化的响应机制对应对未来气候变化具有重要意义[5-6]。众多学者通过大量野外试验或室内模拟研究了山地生态系统植物群落特征及植被地上生产力对气候变化的响应,但不同物种与功能型植物及植被地上生产力对气候变暖的响应还未有一致规律[7-12]。研究显示,增温促进了植被生产力[9-10],或抑制植被生产力[11,13],或对植被生产力的影响不显著[12];不同植物物种或功能型之间的响应也具有差异[14-15]。当前,有关山地草地植物群落盖度及其地上生物量对气候变暖响应的研究多集中于高山草甸/亚高山草甸,且研究结果显示,不同增温幅度和增温时间对草甸植物群落盖度及其地上生物量的影响程度存在差异[16],有必要进一步研究其在不同时空尺度下对全球气候变暖的响应。
吕梁山位于黄土高原东部,水土流失严重,是典型的生态环境脆弱区。近年来有关吕梁山的研究多是针对山地植被多样性的空间变化格局[17-18],并开始逐渐向山地草地响应气候变暖的研究转变[15,18,21]。
因此,本研究选择吕梁山典型山地——管涔山为研究区,利用开顶箱在管涔山不同海拔梯度处设计不同增温幅度试验,调查不同年份增温样地内植物群落的生长特征数据,探究模拟增温对山地林下草本层植物盖度及地上生产力的影响,为山地林下植被生态系统应对气候变暖提供基础数据和理论依据。
1 材料与方法
1.1 研究区概况
本研究区位于山西省吕梁山北段的管涔山(38.25°~39.05°N、111.76°~112.9°E),海拔1 310~2 770 m,其腹地位于山西省宁武县东寨镇,山体呈东北西南走向。管涔山属于温带大陆性季风气候,昼夜温差大,年均气温6~7°C,1月均温-10°C左右,7月均温20.5°C,年均降水量450~500 mm,无霜期105 d。管涔山土壤类型主要为山地褐土、棕壤,土壤pH平均值为5.63,主要分布寒温带针叶林,典型树种为云杉(Piecea asperata)和华北落叶松(Larix gmeliniivar.principis-rupprechtii)。
1.2 样地设计
为了剔除山地不同乔木树种对增温试验的影响,于2016年9月选取管涔山2 222 m和2 571 m两个海拔梯度(均为华北落叶松林),采用随机区组设计,在物种分布均匀、人为干扰小的林下草本层植物群落进行模拟增温实验样地布设,各样地面积为35~40 m2,共计2个样地。对每个样地设置对照(CK)、低度增温(OTC1)和高度增温(OTC2)3种试验处理(图1),每种处理3个重复,共计18个增温样方,各样方间距1~3 m。其中,OTC1和OTC2样方采用无需电力供应的开顶箱(Open-top chamber,OTC)进行模拟增温处理,箱体为半封闭式正六边形,采用有机玻璃(透光率90%以上)构建,各斜边与地面夹角均为60°,OTC1和OTC2两种增温装置的垂直高度分别为40 cm、80 cm;CK样方不进行增温处理,植被处于自然生长状态,取样面积为1 m×1 m[15,18,21]。

图1 模拟增温装置图Fig.1 Simulated warming equipment in the study
1.3 植被特征调查
于2018年和2019年植物生长季对管涔山各样地林下草本群落进行群落结构特征调查,统计林下草本层植物的种类、频度、盖度、高度,计算每个物种的重要值。同时调查所在林地乔木的生长特征指标,包括高度、数量、基径和胸径,以此计算乔木林的盖度。对于林下草本植物频度和盖度的调查,采用的工具为0.5 m×0.5 m样方框,根据样方框大小定制网格尺寸为0.1 m×0.1 m的铁丝网,共计25个网格,测量时将铁丝网平铺于样方框内,统计每个物种在网格中出现的次数和占有的面积。地上生物量采用直接刈割的方法进行采集,运回实验室后先剔除枯枝落叶及土块等杂质,只保留当年植物活体,然后将处理后的样品在80℃的烘箱中烘干,持续24 h至恒重,最后选用精度为1/1 000的电子天平进行称重,获得各样地草本植物地上生物量值。此外,本研究将乔木、灌木、藤本植物的幼苗均归为草本层,各物种名由《中国植物志》得出,再根据植物功能型定义,将所有调查到的植物种划分为禾草、莎草和杂草三种类型。
1.4 环境因子的观测
植物群落特征调查完毕,进行水热环境因子的测量,所使用仪器均来自美国Onset计算机公司生产的HOBO系列产品。在距地面10 cm高度处,架设带有防辐射罩(RS3-B)的温湿度传感器(S-THBM008)测定空气温度和湿度;在距地面10 cm深度处,埋设测量土壤水分(S-SMD-M005)和土壤温度(S-TMB-M006)的传感器。不同海拔梯度的水热环境因子观测是在每天的同一时间点开始连续观测24 h,并采用数据采集器(H21-USB)以1 min为时间间隔进行数据存储。
1.5 数据处理

利用IBM SPSSStatistics 20.0软件进行双因素方差分析,分别检验增温、年份及其交互作用以及增温、海拔及其交互作用对地上生物量、群落总盖度、禾草类植物盖度、莎草类植物盖度和杂草类植物盖度的影响。
利用Canoco for Windows 5.0软件对不同增温处理下温度、水分、植被关系进行排序分析,首先对植被群落结构特征数据进行去趋势对应分析(De⁃trended correspondence analysis,DCA),确定用线性模型进行直接梯度分析(4个排序轴的最大梯度长度小于3),再选择其中的冗余分析(Redundancy analysis,RDA)模型进行约束性排序分析。
2 结果与分析
2.1 林下草本群落水热因子对增温的响应
增温提高了空气温湿度,降低了土壤温湿度,且空气温度、土壤温度具有显著的年际差异性,土壤湿度具有显著的海拔差异性。
OTC1和OTC2处理下,在2018年,空气温度分别提高0.21℃和0.31℃,空气湿度分别升高0.12%和下降0.08%,土壤温度分别下降0.1℃和0.08℃,土壤湿度分别升高6.5%和下降7%;在2019年,空气温度分别升高0.16℃和0.21℃,空气湿度分别升高1.15%和5.83%,土壤温度分别升高0.04℃和0.07℃,土壤湿度分别下降0.33%和4.43%(图2)。

图2 不同增温处理下水热因子在不同年份和海拔的变化Fig.2 Variations of hydrothermic factors along various years and altitudes in different warming treatments
OTC1和OTC2处理下,在低海拔区,空气温度分别升高0.03℃和0.07℃,空气湿度分别升高0.96%和2.44%,土壤温度分别下降0.155℃和升高0.04℃,土壤湿度分别下降3.38%和11.27%;在高海拔区,空气温度分别升高0.34℃和0.44℃,空气湿度分别升高0.31%和3.31%,土壤温度分别升高0.1℃和下降0.05℃,土壤湿度分别升高9.56%和下降0.16%(图2)。
总体看来,与对照相比在不同增温处理下,空气温度分别增加了0.18℃(OTC1)和0.25℃(OTC2),空气湿度分别增加0.63%(OTC1)和2.87%(OTC2),土 壤 温 度 分 别 下 降 了0.03℃(OTC1)和0.01℃(OTC2),土壤湿度增加了3.1%(OTC1)和下降了5.6%(OTC2)。
2.2 林下草本群落植物地上生物量对增温的响应
地上生物量在增温处理下出现显著差异(P=0.007),且存在极显著年际差异(P<0.001),但其海拔差异并不显著(P>0.05)(表2)。
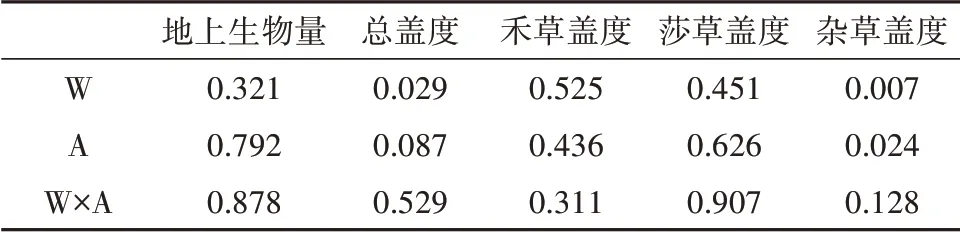
表2 增温和海拔对植物地上生物量、总盖度及不同功能群盖度的影响(P值)Table 2 P-values for the effects of warming and altitude on plant aboveground biomass,total coverage,and coverage of dif⁃ferent functional groups
OTC1和OTC2处理下,在2018年,地上生物量分别减少0.7%和增加了18.12%;在2019年,地上生物量分别减少43.26%和增加了70.78%(图3)。
OTC1和OTC2处理下,在低海拔区,地上生物量分别减少25.03%和增加18.51%;在高海拔区,地上生物量分别减少了3.8%和增加了60.63%(图3)。
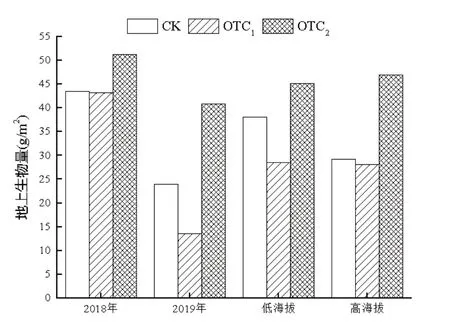
图3 增温对不同年份和不同海拔植物地上生物量的影响Fig.3 Effects of warming on plant aboveground biomass in dif⁃ferent years and altitudes
总体来看,与对照相比在不同增温处理下,植物地上生物量分别减少了15.81%(OTC1)和增加了36.81%(OTC2),在两年短期模拟增温下高度增温促进了林下草本层植物地上生物量的积累。
2.3 林下草本群落不同功能型植物对增温的响应
2.3.1 重要值的变化
不同增温处理下禾草、莎草、杂草类植物的重要值差异并不显著(P>0.05),表明禾草、莎草以及杂草类植物对增温响应不敏感。
OTC1和OTC2处理下,在2018年,莎草类植物重要值上升1.67%和22.09%,杂草类植物重要值下降0.02%和8%;在2019年,莎草类植物重要值下降39.83%和40.33%,杂草类植物重要值上升11.17%和14.14%(表3)。

表3 增温对不同功能型植物重要值的影响Table 3 The effectsof warming on importance value of different functional plants
OTC1和OTC2处理下,在低海拔区,莎草类植物重要值上升9.98%和38.10%,杂草类植物重要值下降7.84%和11.84%;在高海拔区,莎草类植物重要值下降37.62%和39.92%,杂草类植物重要值上升22.57%和21.79%(表3)。
总的来说,与对照相比不同增温处理下,禾草和杂草的重要值有增加趋势,莎草的重要值有下降趋势。因此,增温促进了群落中禾草类、杂草类植物的生长,抑制了莎草类植物的生长。
2.3.2 盖度的变化
莎草类植物盖度存在显著的年际差异(P<0.05),增温显著改变了群落总盖度和莎草类植物盖度(P<0.05),增温和年份的交互作用显著改变了莎草类植物盖度(P<0.05)(表1)。

表1 增温和年份对植物地上生物量、总盖度及不同功能群盖度的影响(P值)Table 1 P-valuesfor the effects of warming and year on plant aboveground biomass,total coverage,and coverage of different functional groups
OTC1和OTC2处理下,在2018年,植物群落总盖度分别增加了21.65%和63.50%(P<0.05),莎草类植物盖度分别增加了12.25%和25.39%(P<0.05),杂草类植物盖度分别增加了9.28%和36.34%;在2019年,植物群落总盖度分别减少了2.90%和增加了28.36%,莎草类植物盖度分别下降了4.87%和2.37%,杂草类植物盖度分别增加了0.91%和30.46%(图4)。

图4 增温对植物总盖度及不同功能型植物盖度的影响Fig.4 Warming effects on plant total coverage and coverage of different plant functional groups
杂草类植物盖度存在显著的海拔差异(P<0.05),增温显著改变了群落总盖度、杂草类植物盖度(P<0.05),增温和海拔的交互作用对群落总盖度、禾草类植物盖度、莎草类植物盖度和杂草类植物盖度无显著影响(P<0.05)(表2)。
OTC1和OTC2处理下,在低海拔区,植物群落总盖度分别增加了12.61%和34.28%,莎草类植物盖度分别增加了5.25%和15.35%,杂草类植物盖度分别增加了6.23%和18.80%;在高海拔区,植物群落总盖度分别增加了6.14%和57.58%(P<0.05),莎草类植物盖度分别增加了2.13%和7.67%,杂草类植物盖度分别增加了3.96%和48%(P<0.05)(图4)。
总体来看,与对照相比在不同增温处理下,植物总盖度分别增加了20.76%(OTC1)和101.69%(OTC2),莎草盖度分别增加了39.08%(OTC1)和121.85%(OTC2),杂草盖度分别增加了14.33%(OTC1)和93.96%(OTC2)。因此,高度增温显著提高了植物群落总盖度和杂草盖度(P<0.05)。
2.4 林下草本群落植物与水热因子关系对增温的响应
植物地上生物量和总盖度与水热因子的相关性均未达到显著水平,但其相关性随温度升高而增强(图5)。植物地上生物量在不同增温处理下,与空气温度、空气湿度、土壤温度及土壤湿度均呈正相关关系。

图5 不同增温处理下植物与水热因子的关系Fig.5 Correlations among temperature,moisture,and vegetation in different warming treatments
植物总盖度在CK处理下,与空气温度、土壤湿度呈正相关,与空气湿度、土壤温度呈负相关;在OTC1处理下,与空气湿度、土壤温湿度呈正相关,与空气温度呈负相关;在OTC2处理下,与空气温湿度以及土壤温湿度呈正相关。
在CK处理下,禾草、莎草、杂草盖度与空气温度、土壤湿度呈正相关,与土壤温度呈负相关。在OTC1处理下,莎草、杂草盖度与空气湿度、土壤温湿度呈正相关,与空气温度呈负相关;禾草盖度与空气温度呈正相关,与空气湿度、土壤温湿度呈负相关。在OTC2处理下,禾草、杂草盖度与土壤温度呈负相关,与空气温湿度、土壤湿度呈正相关;莎草盖度与空气温度呈负相关,与空气湿度和土壤温湿度呈正相关。
总体而言,在OTC1处理下空气湿度对植物群落的贡献率最高,而在OTC2处理下土壤湿度对植物群落的贡献率最高(表4)。这表明气候变化通过改变水分有效性对植物群落产生间接影响。

表4 不同增温处理下环境因子的贡献分析Table 4 Contribution analyses of environmental factors in dif⁃ferent warming treatments
3 讨论
3.1 增温对水热因子的影响
本研究采用OTC作为增温装置,设置低、高两个增温梯度,分别使空气温度最高增加0.36℃和0.5℃。姜炎彬等[19]发现,OTC高度越高、开口越小,则增温幅度就越大,这与本研究的结论相一致。由于OTC阻挡了空气的水平运动,空气湍流减弱,使热量不易散失,因而提高了箱内空气温度[20]。本研究发现:增温对空气温湿度产生一定的正效应,且空气温度具有显著的年际差异性。空气温度升高增强土壤水分蒸发和植物蒸腾,但由于近地面植被层和OTC的阻挡作用,水分不易扩散,从而使得OTC内空气湿度增加[21]。李英年等[22]报道,OTC内地表蒸散高于对照,导致OTC内地表至植物层出现暂时相对干燥的低湿度环境,这与本研究的结论相类似。本研究测定的空气温湿度是距地面10 c m高度的植被层,受植被盖度影响较大,因而在近地面易形成空气湿度相对较高的环境。本研究发现增温对土壤温湿度产生一定的负效应,且土壤温度具有显著的年际差异性,土壤湿度具有显著的海拔差异性。其中增温对土壤湿度产生负效应,这与大多数学者的研究一致,即增温增强了土壤水分蒸发,导致土壤出现干化趋势[19,23-24]。
3.2 增温对地上生物量的影响
管涔山林下草本层植物地上生物量对小于三年的短期增温响应不敏感。本研究中,在2018和2019年OTC1处理下抑制了植被地上生物量的累积,而OTC2处理下促进了林下草本群落植被地上生物量累积,总的来看增温有利于地上生物量的累积。Chen等[25]通过对青藏高原高寒草甸3年模拟增温试验发现,温度升高显著提高了禾本科和豆科植物的生物量,总初级生产力有所增强。在温度较低的区域温度是植物生长的主要限制因子[26],本研究中在高海拔增温处理下均提高了地上生物量,这与姜炎彬等[19]的研究结论相一致,即高海拔地区植物所处的环境温度远远低于植物生长所需的最适温度。因此,增温通常都会促进植物的生长和生物量的累积[27]。李英年等[22]研究发现,高寒矮嵩草草甸生物量在模拟增温试验初期呈增加趋势,但增温时间持续5年后生物量反而下降。因此,短期增温对地上生物量有正效应,但影响不显著。
3.3 增温对群落盖度的影响
本研究中增温显著改变了群落总盖度、莎草盖度和杂草盖度,禾草盖度呈增加趋势但不明显,得到高度增温下植物群落盖度的增幅基本都是高于低度增温。不同增温幅度对整个群落的高度、盖度、多样性和生物量产生一定影响[28-29]:增温幅度较低时,群落的高度、盖度和生物量的积累以及多样性会有一定程度的增加;但当温度升高幅度过大时,群落的盖度、高度和地上生物量又逐渐降低[30]。之所以会这样,是由于大幅度增温加剧了土壤水分蒸发,不利于植物生长,从而导致高度增温条件下植被群落特征呈下降趋势。本研究中增温使空气温度最高增加0.36℃和0.5℃,温度升高幅度不够大且研究周期较短。因此,增温控制在一定幅度内能对植被产生正效应。
3.4 增温对不同功能型植物的影响
不同功能型植物对温度的适应性和敏感性不同,因此增温后表现出不同的适应模式。在青海湖鸟岛的研究发现,模拟增温会导致禾草类植物种比例增加,杂草比例有所减少[31]。然而,本研究中气候变暖对禾草、莎草和杂草类植物的重要值没有显著影响,且不同增温处理下禾草和杂草的重要值有增加趋势,莎草的重要值有下降趋势。禾草、莎草和杂草对增温的不同响应,是由其自身的生物特性差异引起的。
增温显著增加了高寒草原莎草和杂草的盖度,这与本研究结论相一致[31];但也有研究发现增温显著增加了高寒草原豆科植物的盖度,降低了禾草和杂草的盖度,并认为土壤水分的变化是禾草类植物盖度变化的重要原因[32]。这说明增温和植物群落种间竞争对植物群落盖度变化均有影响,植物群落种间竞争的结果是植物群落对增温的间接响应[31]。
4 结论
在管涔山进行为期3年的模拟增温实验,增温对空气温湿度产生正效应,气候呈现暖湿化;对土壤温湿度产生负效应,且土壤温度的响应程度具有明显的年际差异性,土壤水分具有显著海拔差异性。增温对植物盖度和地上生物量产生正效应,适度增温可促进植物生长发育和生物量生产。年际间的增温增加了植物群落总盖度、莎草类植物盖度和杂草类植物盖度,但禾草类植物盖度对年际间的增温响应不敏感。莎草类植物盖度存在显著的年际差异,杂草类植物盖度存在显著的海拔差异。因此,短期增温显著影响了林下草本层植物群落总盖度和地上生物量,改变了禾草、莎草和杂草类植物的盖度和重要值,在较长时间尺度上可能会改变植物群落结构。
猜你喜欢
疯狂英语·新读写(2022年1期)2022-11-23
疯狂英语·读写版(2022年1期)2022-01-29
草地学报(2021年7期)2021-08-03
文史博览·文史(2021年1期)2021-04-01
黑龙江工程学院学报(2020年5期)2020-10-21
水土保持研究(2019年6期)2019-10-19
南方农业·下旬(2017年11期)2018-01-09
中国饲料(2017年15期)2017-08-22
杂草学报(2015年2期)2016-01-04
海南热带海洋学院学报(2015年5期)2015-03-14
