
糖蜜添加量对杂交构树青贮发酵品质和微生物多样性的影响
2021-11-06 02:58姜富贵成海建魏晨张召坤苏文政时光宋恩亮
生物技术通报 2021年9期
姜富贵 成海建 魏晨 张召坤 苏文政 时光 宋恩亮
(山东省农业科学院畜牧兽医研究所 山东省畜禽疫病防治与繁育重点实验室,济南 250100)
饲料是畜牧业发展的基础。随着我国居民生活水平提高,消费者对肉蛋奶的需求日益增长,导致饲料工业面临原料短缺的局面,尤其是蛋白质资源的国外进口依存度高达80%,严重制约我国畜牧业的发展[1]。构树(Broussonetia papyrifera)又称褚树或构皮树,桑科构树属的双子叶植物。杂交构树(hybrid Broussonetia papyrifera L. Vent)是中国科学院采用杂交育种技术培育出的新树种,具有生长速度快、生物产量大、抗逆性强和饲用价值高等特点[2]。此外,杂交构树粗蛋白的含量高且在瘤胃中的降解率较高[3],以杂交构树作为饲料资源有利于缓解我国蛋白饲料短缺的局面。青贮是当前杂交构树饲料保存的主要手段,不仅生产成本低而且能够改善杂交构树的适口性。但是,由于杂交构树的水溶性碳水化合物含量低且缓冲能高,单独青贮的发酵品质较差,需要添加剂提升其发酵品质[4]。
糖蜜是制糖过程中的一种副产物,不仅含有糖类、蛋白质、维生素、矿物质等营养物质,而且含有低聚糖、磷脂、酚类等生物活性物质[5]。在畜牧生产上,糖蜜不仅能够作为一种能量饲料来提高动物的生产性能和饲料转化效率[6-7],而且能够作为青贮发酵促进剂来快速降低pH,同时抑制丁酸产生和蛋白水解,进而改善青贮饲料的发酵品质[5]。黄秋连等[8]研究发现,添加糖蜜可提升羊草青贮的粗蛋白含量并降低氨态氮含量。付锦涛等[9]研究发现,添加糖蜜可降低全株构树与稻草混合青贮的pH,同时降低中性洗涤纤维(NDF)和酸性洗涤纤维(ADF)含量。穆麟等[10]研究发现,添加糖蜜可降低籽粒苋与稻秸混合青贮的干物质损失,提高乳酸含量和有氧稳定性。然而,当前研究主要是探讨单一糖蜜添加量对某种饲草营养成分含量和发酵品质的影响,针对不同糖蜜添加量对杂交构树青贮发酵品质和微生物多样性影响的研究较少。因此,本研究以杂交构树为原料,探究不同糖蜜添加量对杂交构树青贮营养成分、发酵品质和微生物多样性的影响,为杂交构树的饲料化应用提供数据支持。
1 材料与方法
1.1 材料
构树原料于2020年6月18日取自山东省华悦中科生态科技有限公司种植基地(116°16' E,36°45'N),在株高1.2 m时采用自动收割机进行刈割,留茬高度约为30 cm,其营养成分见表1。糖蜜主要成分为木糖,含糖量约为55%,购自山东耀佳化工有限公司。
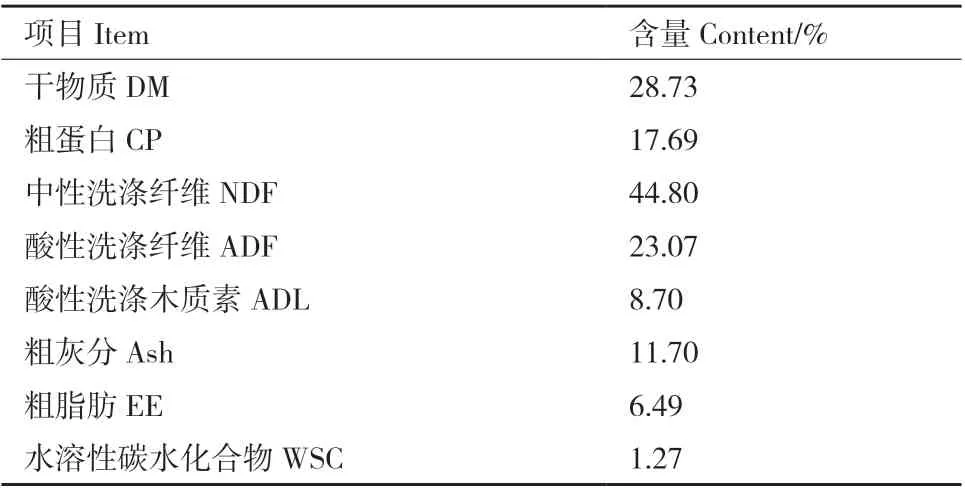
表1 杂交构树原料的营养成分(干物质基础)Table 1 Nutritional composition of hybrid B. papyrifera L.Vent(dry matter basis)
1.2 方法
1.2.1 试验设计 试验采用单因子完全随机设计,共设置4个处理组,以鲜重为基础,试验组糖蜜添加量分别为5 g/kg(M5组)、10 g/kg(M10组)和20 g/kg(M20组),对照组(CK组)添加与试验组等体积的蒸馏水。各组混合均匀后,取约1 kg样品装入聚乙烯厌氧袋(22 cm×40 cm)中,每组4个重复,抽真空密封,于室温(25-30℃)避光贮藏60 d后开袋取样,后续测定相关指标。
1.2.2 测定指标和方法
1.2.2.1 常规营养成分测定 称取500 g青贮样品并平铺在不锈钢托盘上,于65℃鼓风干燥箱中烘至恒重,用于常规营养成分测定。干物质(DM)、粗蛋白(CP)、粗灰分(Ash)和粗脂肪(EE)含量测定参照AOAC(2005)[11]的方法,其中采用重量法测定DM含量,采用凯氏定氮法测定CP含量,采用灰化法测定Ash含量,采用索氏浸提法测定EE含量。NDF、ADF和酸性洗涤木质素(ADL)含量测定参照Van Soest等[12]的方法,利用Ringbio纤维分析仪(R-2000,英国)测定。水溶性碳水化合物(WSC)含量采用蒽酮-硫酸比色法[13]测定。
1.2.2.2 发酵指标测定 称取青贮样品20 g,加入180 mL蒸馏水,利用榨汁机进行榨汁并用4层纱布过滤获取浸提液,立即使用pH计(HANNA HI 9125,意大利)测定pH值,剩余滤液-20℃保存用于测定乳酸、挥发性脂肪酸(VFA)和氨态氮(NH3-N)含量。乳酸和VFA含量使用高效液相色谱法测定[14]。NH3-N含量采用苯酚-次氯酸钠比色法测定[15]。
1.2.2.3 微生物多样性
(1)测序流程 称取青贮样品10 g于三角瓶中,加入90 mL蒸馏水,制备浸提液,于离心机10 000 r/min离心15 min,取沉淀;采用E.Z.N.A.®soil试 剂 盒(Omega Bio-tek,Norcross,USA)抽提DNA,利用NanoDrop2000检测DNA浓度和纯度,利用1%琼脂糖凝胶电泳检测DNA质量;采用338F(5'-ACTCCTACGGGAGGCAGCAG-3') 和806R(5'-GGACTACHVGGGTWTCTAAT-3')引物对V3-V4可变区进行PCR扩增;利用AxyPrep DNA Gel Extraction Kit(Axygen Biosciences,Union City,CA,USA)进行纯化,利用QuantiFluorTM-ST(Promega,USA)进行检测定量,纯化后扩增片段构建PE2×300的文库,在Illumina公司的Miseq PE300平台进行测序。
(2)信息分析 利用Trimmomatic软件对低质量序列进行过滤,进一步利用FLASH软件进行拼接,同时利用UCHIME软件去除序嵌合体,然后使用UPARSE软件按照97%的一致性对序列进行OUT聚类。获取OTU后,使用 QIIME 软件筛选出每个OTU 的代表序列,利用RDP classifier软件与Silva数据库(SSU138)比对并进行物种分类注释,保留置信区间大于0.7的结果。
1.2.3 数据统计分析 营养成分和发酵品质数据使用SAS 9.1软件中GLM过程进行方差分析和Duncan氏多重比较检验,结果以最小二乘均值表示;营养成分和发酵品质指标随糖蜜添加量的直线、二次和三次曲线反应利用CONTRAST语句实现。微生物多样性数据采用Majorbio生物云在线平台进行OTU分析、Alpha多样性分析、β多样性分析、群落组成分析和物种差异分析,其中Alpha多样性组间差异分析采用Student's t检验,β多样性组间差异采用ANOSIM分析,物种差异分析采用Kruskal-Wallis秩和检验。P < 0.05为差异显著,0.05 ≤ P < 0.10为存在趋势。
2 结果
2.1 糖蜜添加量对杂交构树青贮营养成分含量的影响
由表2可知,随糖蜜添加量的增加,杂交构树青贮的DM、CP和WSC含量显著线性增加(P < 0.05),NDF、ADF和Ash含量显著线性降低(P < 0.05)。糖蜜添加量对EE和ADL含量的影响较小,差异不显著(P = 0.733)。M5组、M10组和M20组的DM含量较CK组显著增加(P < 0.05),而Ash含量显著降低(P = 0.009)。M20组的CP和WSC含量较CK组显著增加(P < 0.05),而NDF和ADF含量显著降低(P < 0.001)。
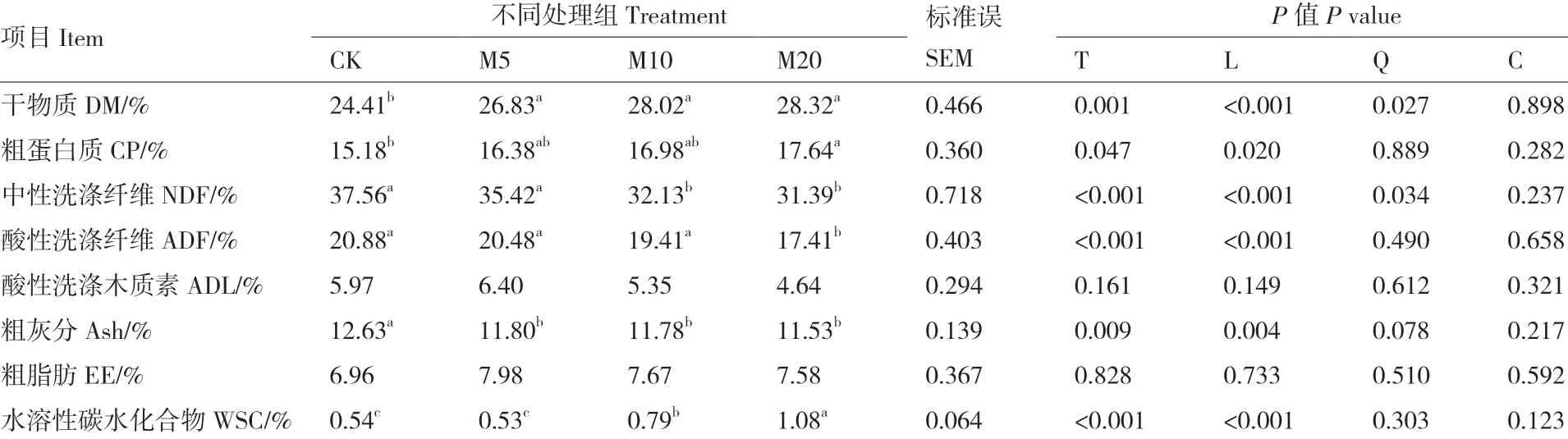
表2 糖蜜添加量对杂交构树青贮营养成分含量的影响(干物质基础)Table 2 Effects of addition amount of molasses on nutrient contents of hybrid B. papyrifera L. Vent silage(dry matter basis)
2.2 糖蜜添加量对杂交构树青贮发酵品质的影响
由表3可知,随糖蜜添加量的增加,杂交构树青贮的pH值、丙酸、丁酸含量和NH3-N/TN显著线性降低(P < 0.05),乳酸含量显著线性增加(P =0.002)。糖蜜添加量对乙酸含量的影响较小,差异不显著(P = 0.131)。M10组和M20组的乳酸含量较CK组显著增加(P = 0.024),而丙酸含量显著降低(P = 0.043)。M5组、M10组和M20组的pH值、丁酸含量和NH3-N/TN较CK组显著降低(P < 0.01)。
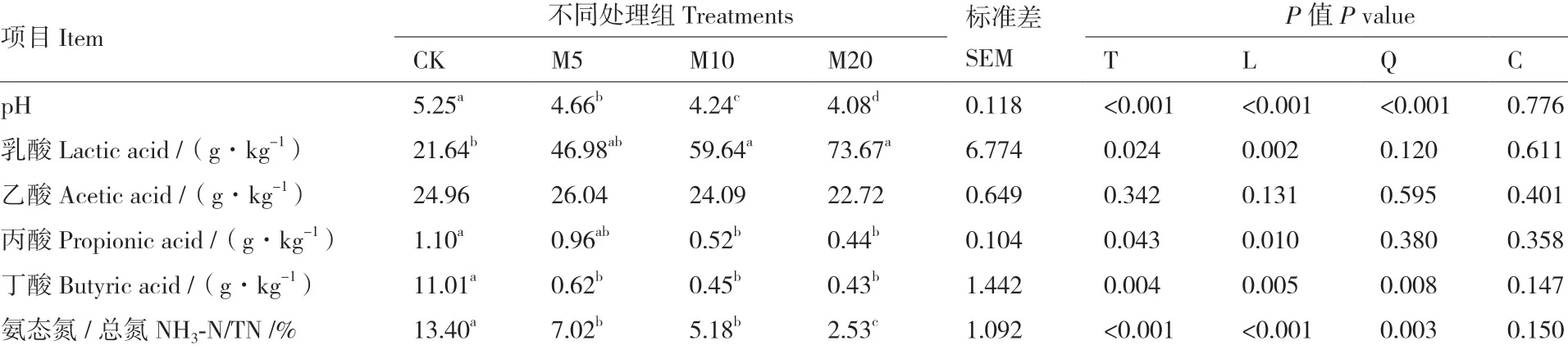
表3 糖蜜添加量对杂交构树青贮发酵品质的影响(干物质基础)Table 3 Effects of addition amount of molasses on the fermentation quality of hybrid B. papyrifera L. Vent silage(dry matter basis)
2.3 糖蜜添加量对杂交构树青贮微生物多样性的影响
2.3.1 Alpha多样性分析 通过对杂交构树4个鲜样和16个青贮样品进行测序分析,共获得723 460条高质量序列并聚类成310个OTUs,各处理的覆盖深度(Coverage指数)均大于0.99。基于OUT水平的Alpha多样性分析结果如图1所示,M20组青贮的Ace和Chao指数较鲜样显著增加(P < 0.05),而Shannon和Simpson指数无显著差异。随糖蜜添加量的增加,Ace、Chao和Simpson指数显著线性增加(P< 0.05),而Shannon指数显著线性降低(P = 0.010)。
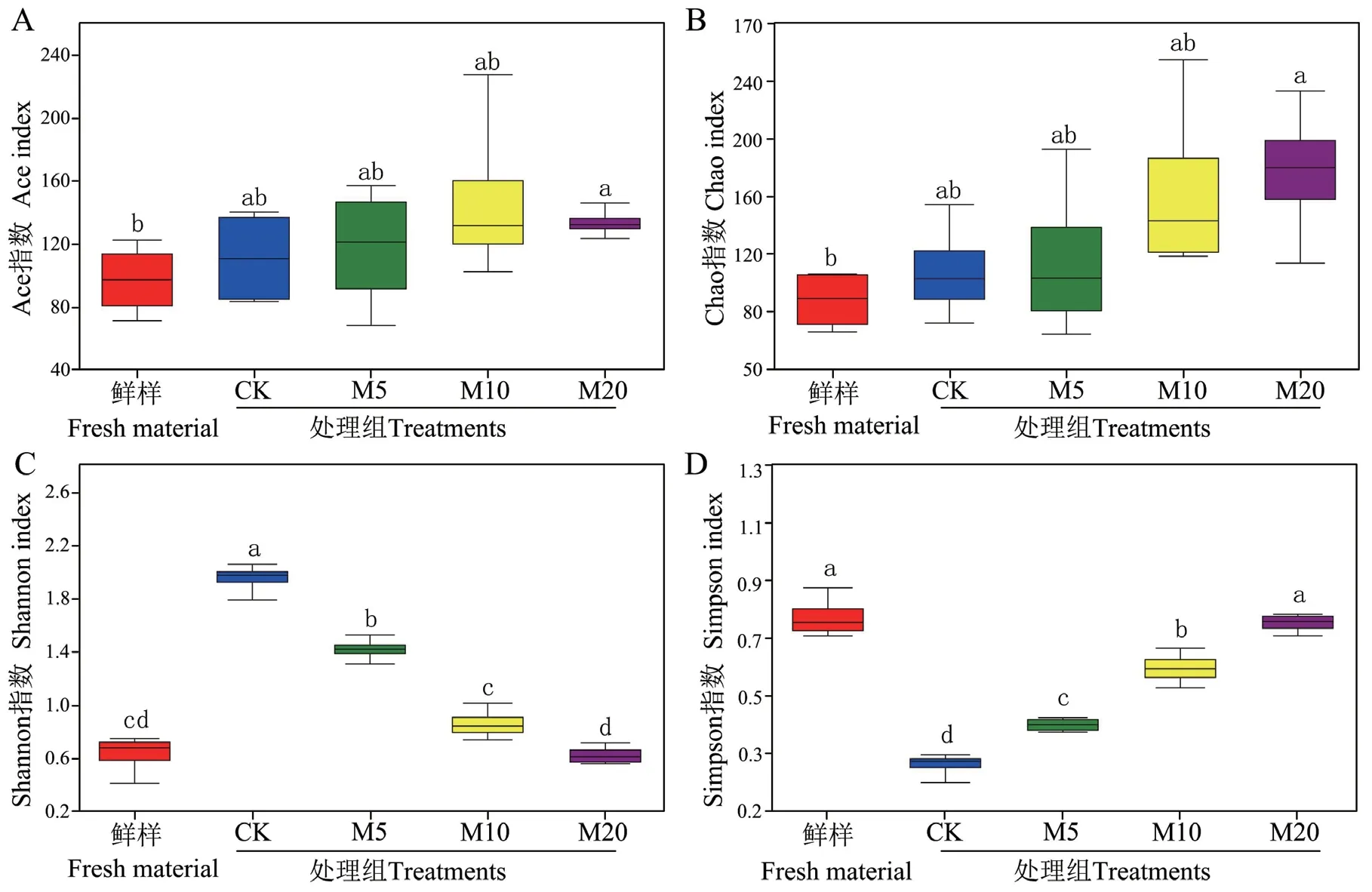
图1 杂交构树鲜样和青贮的Alpha多样性分析Fig. 1 Alpha diversity analysis of hybrid B. papyrifera L. Vent fresh material and silage
2.3.2 β多样性分析 基于Bray-Curtis距离的PCoA分析结果如图2所示,杂交构树鲜样与青贮样品的微生物群落组成存在显著差异(P = 0.004);CK组与M5组的微生物群落组成存在显著差异(P =0.034);M10组与M20组的微生物群落组成存在显著差异(P = 0.031);M10组和M20组的微生物群落组成与CK组和M5组存在显著差异(P = 0.001)。
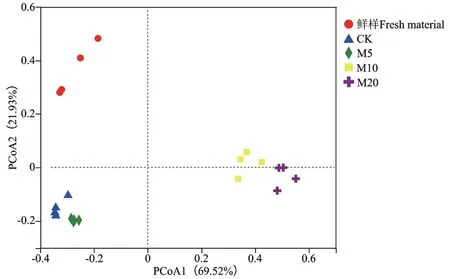
图2 杂交构树鲜样和青贮的PCoA分析Fig. 2 Principal co-ordinates analysis of hybrid B. papyrifera L. Vent fresh material and silage
2.3.3 微生物群落组成分析 如图3-A所示,在门水平上,杂交构树鲜样和青贮的优势菌门均为厚壁菌门(Firmicutes),相对丰度分别为94.41%和94.11%,其次为变形菌门(Proteobacteria)相对丰度分别为5.39%和5.83%。M5、M10和M20组厚壁菌门的相对丰度较CK组均升高,分别为98.71%、99.02%和99.08%。
如图3-B所示,在属水平上,杂交构树鲜样中优势菌属为魏斯氏菌属(Weissella)和肠杆菌属(Enterobacter),相对丰度分别为87.82%和4.20%。CK组优势菌属为魏斯氏菌属、乳杆菌属(Lactobacillus)、Clostridium_sensu_stricto_12和 肠 杆菌属,相对丰度分别为49.41%、31.07%、8.05%和4.61%。M5组的优势菌属为魏斯氏菌属,相对丰度为62.96%,其次为乳杆菌属,相对丰度为32.88%。M10组和M20组的优势菌属均为乳杆菌属,相对丰度分别为82.13%和91.81%,其次为魏斯氏菌属,相对丰度分别为16.25%和6.95%。
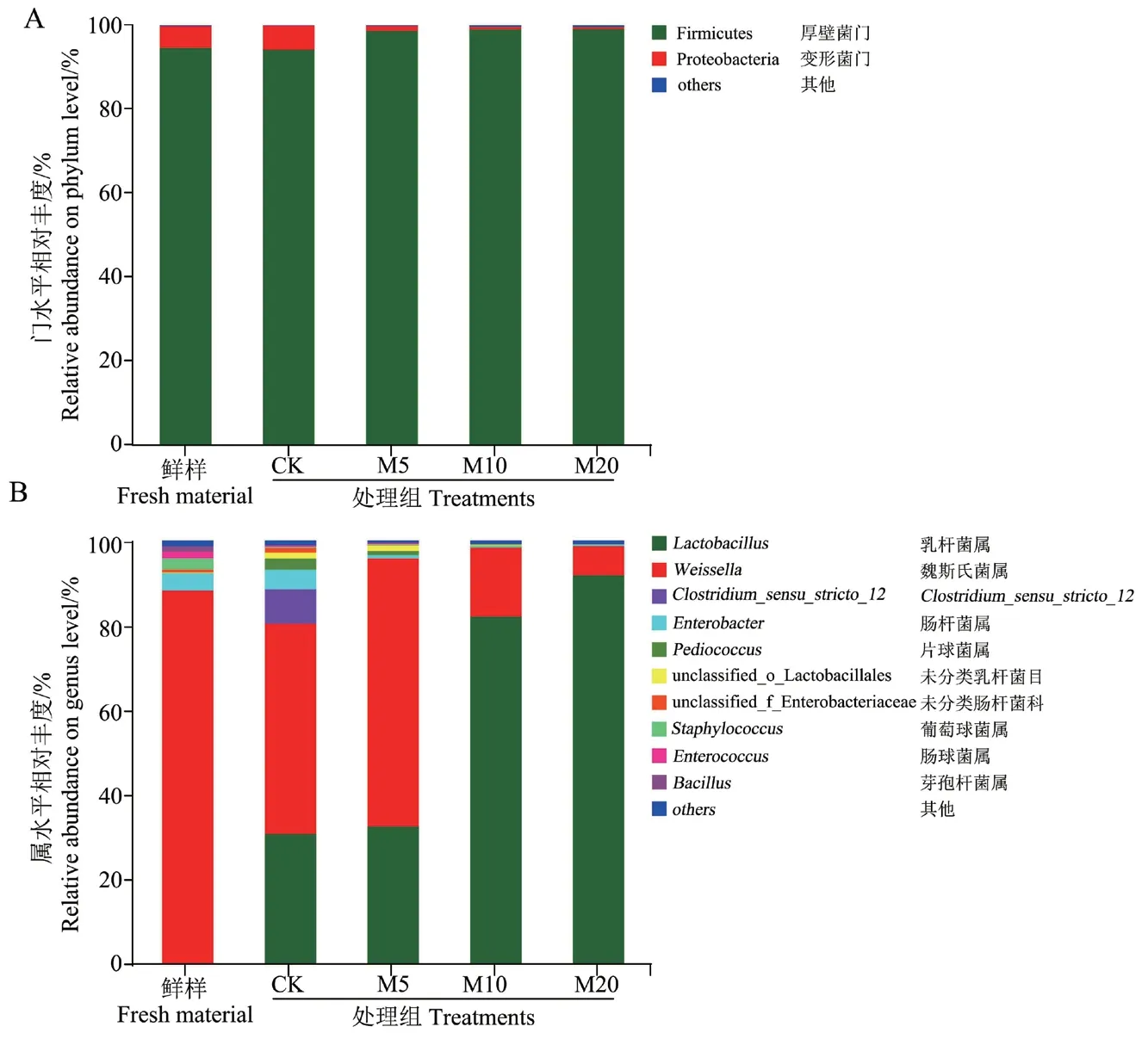
图3 杂交构树鲜样和青贮在门水平(A)和属水平(B)的微生物群落组成Fig. 3 Microbial community structure of hybrid B. papy-rifera L. Vent fresh material and silage at phylum(A)and genus(B)level
2.3.4 微生物菌群的差异性分析 各处理组在属水平上的菌群差异性分析结果如图4所示,M5组乳杆菌属的比例较CK组无显著变化(P > 0.05),但魏斯氏菌属的比例显著增加(P = 0.003)。M10组和M20组乳杆菌属的比例较CK组和M5组均显著增加(P = 0.005),而魏斯氏菌属的比例显著降低(P =0.003)。M5组、M10组和M20组Clostridium_sensu_stricto_12、肠杆菌属和片球菌属的比例较CK组均显著降低(P < 0.05)。

图4 各组杂交构树青贮在属水平菌群的差异性分析Fig. 4 Difference analysis of flora of hybrid B. papyrifera L. Vent silage groups at genus level
3 讨论
3.1 糖蜜添加量对杂交构树青贮营养成分含量的影响
杂交构树青贮后的DM和WSC含量较青贮前显著降低,与王亚芳等[16]和周昕等[17]的研究结果一致,主要与青贮前期植物细胞的呼吸作用对DM的消耗以及青贮后期乳酸菌利用WSC发酵产生乳酸有关[18]。本试验中,杂交构树原料的营养成分与黄媛等[4]和司丙文等[19]的研究结果存在差异,其原因是杂交构树的营养成分受刈割高度、留茬高度、生长期和田间管理等多因素的影响[20]。曹力凡等[21]发现,随刈割高度的增加,杂交构树原料的NDF、ADF和ADL含量显著增加,而CP含量显著降低;随留茬高度的增加,NDF、ADF和ADL含量显著降低,而CP含量显著增加。本试验中,与CK组相比,M20组的DM和CP含量显著增加,而NDF和ADF含量显著降低,与黄秋连等[8]、穆麟等[10]和周昕等[17]的研究结果一致,M20组高DM和CP含量可能是因为添加糖蜜促进乳酸菌的增殖,pH快速降低抑制了有害微生物生长,进而减少了营养物质的损失[18];CK组 高NDF和ADF含 量 可 能 是 其Clostridium_sensu_stricto_12和肠杆菌属消耗了青贮中蛋白质和糖等营养物质,降低青贮DM含量的同时增加了NDF和ADF在DM中的比例,而M20组低NDF和ADF含量可能与乳酸菌产生的有机酸和纤维素酶对纤维素和半纤维素的降解有关[22]。NDF和ADF含量是评定粗饲料综合营养价值的重要指标,二者含量越低则粗饲料的品质越高[16],因此添加糖蜜可提升杂交构树青贮的营养价值。
3.2 糖蜜添加量对杂交构树青贮发酵品质的影响
青贮的pH、乳酸含量和NH3-N/TN是评定青贮发酵品质的关键指标。一般来说,优质青贮的pH应小于4.2,乳酸含量大于4%,NH3-N/TN小于10%[23]。参照此标准,CK组的3项指标均未达到优质青贮饲料的要求,表明杂交构树较难青贮,单独青贮的发酵品质较差。司丙文等[19]认为,杂交构树不易青贮与其附生乳酸菌数量少、WSC含量低和缓冲能高有关。通常情况下,生产优质青贮饲料的原料WSC含量(DM基础)应大于6%[24],本试验用杂交构树原料的WSC含量仅为1.27%,而添加20 g/kg糖蜜(M20组)的pH、乳酸含量和NH3-N/TN均达到优质青贮饲料的要求,因此低WSC含量可能是限制杂交构树发酵的首要因素。本试验中,随糖蜜添加量的增加,pH线性降低,乳酸含量线性增加,与付锦涛等[9]和司丙文等[19]的研究结果一致,主要是因为糖蜜中的WSC为乳酸菌的快速增殖提供了充足的发酵底物,增加乳酸含量的同时降低了pH[8]。本试验中,杂交构树青贮中乙酸含量受糖蜜添加量的影响较小,与付锦涛等[9]的研究结果一致,但显著高于王亚芳[16]和施巧婷[25]报道的全株玉米青贮中的乙酸含量,究其原因,可能与其WSC含量不足有关,而糖蜜添加组可能与其附生布氏乳杆菌的作用有关。在WSC含量不足时,乳酸菌以乳酸为能量来源发酵产生乙酸[26],而布氏乳杆菌属于异型发酵乳酸菌,发酵产生乳酸的同时产生等量的乙酸[27]。青贮中丁酸和氨态氮含量严重降低饲料的营养价值和适口性,NH3-N/TN反映了青贮中蛋白质和氨基酸的降解程度,其值越高则蛋白氮的降解越多[16]。CK组高丁酸含量和高NH3-N/TN主要与其高Clostridium_sensu_stricto_12丰度有关,Clostridium_sensu_stricto_12不仅能够通过氨基酸脱氨基作用生成氨态氮,而且可以利用糖产生丁酸[28]。随糖蜜添加量的增加,丁酸含量和NH3-N/TN显著线性降低,可能是因为低pH抑制了梭菌等有害菌的增殖[29]。综上所述,添加糖蜜可提升杂交构树青贮的发酵品质。
3.3 糖蜜添加量对杂交构树青贮微生物多样性的影响
当前有关青贮前后菌群丰度和多样性变化的研究结果存在分歧,与青贮前相比,大豆青贮[30]和辣木叶青贮[31]的菌群丰度和多样性显著增加,而全株玉米青贮[32-33]显著降低,除与原料种类有关,还可能与发酵时间有关。Ren等[34]发现,甘蔗青贮菌群的丰度和多样性在发酵60 d显著增加,而在发酵90 d显著降低;Jiang等[22]发现,全株玉米青贮菌群的丰度和多样性在发酵45 d显著增加,在发酵90 d显著降低。由于在青贮发酵过程中乳酸菌逐渐占主导地位,微生物菌落多样性降低通常作为青贮发酵成功的标志[32],本试验中,随糖蜜添加量的增加,菌群的多样性线性降低,表明添加糖蜜加速了青贮发酵进程。在门水平上,青贮原料的微生物群落结构受原料种类的影响较小,辣木叶[31]和全株玉米[22]均以厚壁菌门和变形菌门为主,与本研究结果一致。但在属水平上,不同青贮原料的微生物群落结构存在较大差异,如全株玉米以明串珠菌属(Leuconostoc)和克雷白氏杆菌属(Klebsiella)为主,全株大豆以肠杆菌属和泛菌属(Pantoea)为主[30],辣木叶以微小杆菌属(Exiguobacterium)和不动杆菌属(Acinetobacter)为主[31]。本试验中,CK组和M5组的优势菌属为魏斯氏菌属,M10和M20组的优势菌属由魏斯氏菌属转变为乳杆菌属,其原因可能是魏斯氏菌属主要在青贮发酵前期发挥作用,在发酵后期数量会发生下降[35],糖蜜作为发酵促进剂加快发酵进程的同时降低了魏斯氏菌属的数量。本试验中,添加糖蜜可显著降低Clostridium_sensu_stricto_12和肠杆菌属的比例,Clostridium_sensu_stricto_12与CK组高NH3-N/TN和高丁酸含量直接相关[28];肠杆菌属为无芽孢兼性厌氧菌,可发酵糖和乳酸产生乙酸,增加干物质损失,降低饲料营养价值[30]。因此,糖蜜可影响杂交构树青贮的微生物群落结构,降低有害微生物的数量。
4 结论
杂交构树青贮中添加糖蜜对其营养价值和发酵品质均有不同程度的提升作用,并可增加乳杆菌属的比例,同时降低Clostridium_sensu_stricto_12和肠杆菌属等有害微生物的数量。本试验条件下,添加20 g/kg糖蜜,杂交构树青贮的营养价值和发酵品质最优。
猜你喜欢
河南医学研究(2022年19期)2022-10-19
中国奶牛(2022年6期)2022-07-01
环境昆虫学报(2021年5期)2021-10-20
山东畜牧兽医(2021年4期)2021-04-28
国际消化病杂志(2021年1期)2021-03-05
中国油脂(2020年5期)2020-05-16
绿色中国(2019年16期)2019-11-26
中国林业产业(2019年3期)2019-03-25
中国扶贫(2017年16期)2017-08-16
西南学林(2016年2期)2016-11-08
