
四氯虫酰胺对棉铃虫生物活性的温度效应及其解毒酶相关机制
2021-11-05 08:33窦亚楠孙小诺安静杰高占林郭江龙党志红李耀发
新疆农业科学 2021年8期
窦亚楠,孙小诺,安静杰,高占林,郭江龙,党志红,李耀发
(1.河北省农林科学院植物保护研究所/河北省农业有害生物综合防治工程技术研究中心/农业农村部华北北部作物有害生物综合治理重点实验室,河北保定 071000;2.保定市气象局,河北保定 071000)
0 引 言
【研究意义】棉铃虫HelicoverpaarmigeraHübner是一种世界性害虫,其寄主范围广,包括棉花、玉米、花生、大豆等200余种作物,也包括多种蔬菜、果树、花卉等[1,2]。棉铃虫在我国从南到北各省均有不同程度的发生,其中长江流域、黄河流域和西北内陆棉区受害最严重[3]。棉铃虫成虫对棉花具有明显的产卵偏好性,Bt棉花破坏了其寄主转移过程,棉铃虫种群数量得到了有效控制[1]。2010年以来,随着种植结构调整,长江流域和黄河流域棉花种植面积不断减少,玉米、蔬菜面积种植比率逐渐升高,棉铃虫在棉花之外的各寄主上的危害逐年加重,同时多地花生、油葵、大豆和番茄等作物及蔬菜均受到严重危害[4]。研究杀虫剂的温度效应,分析其相关影响机制,对提高双酰胺类杀虫剂对棉蚜虫的防治有实际意义。【前人研究进展】温度不仅影响杀虫剂自身的理化性质,还能影响昆虫的生长发育和新陈代谢,产生不同的温度效应[5]。已有研究发现,乙酰胆碱酯酶抑制剂类的有机磷类和氨基甲酸酯类杀虫剂,通常被认为是正温度系数杀虫剂[6-7],而钠离子通道抑制剂类的拟除虫菊酯类杀虫剂在低温下的毒力显著高于较高温度,常表现为负温度效应[8]。对于不同种类昆虫,同一种杀虫剂也会出现不同程度的温度效应,比如辛硫磷对于麦长管蚜,在10~25℃表现为较显著的正温度效应,而对于绿盲蝽来说,温度对其生物活性的影响很小[9]。鱼尼丁受体抑制剂双酰胺类杀虫剂对鳞翅目害虫防治效果优异,且对非靶标生物安全,其中四氯虫酰胺(tetrachlorantraniliprole)是沈阳化工研究院创制的双酰胺类杀虫剂品种,具有极高的应用前景和开发价值[10]。研究表明,鱼尼丁受体抑制剂类杀虫剂对多种鳞翅目害虫均为正温度效应,但其温度效应程度存在显著差异[11-12]。昆虫体内重要的解毒酶系,羧酸酯酶(CarE)、多功能氧化酶(MFO)、谷胱甘肽S-转移酶(GST)以及尿苷二磷酸糖基转移酶(UGT),在机体代谢外源物质、保护细胞免受急性毒性化学物质攻击中起重要作用[13]。经不同杀虫剂诱导之后,麦长管蚜GST活性与负温度系数杀虫剂密切相关,CarE则与正温度系数杀虫剂有关[14];15~35℃诱导绿盲蝽解毒酶发现,GST参与了负温度效应杀虫剂溴虫腈的代谢[15]。【本研究切入点】昆虫体内解毒酶参与调节了杀虫剂温度效应的影响机制,但有关于双酰胺类杀虫剂对鳞翅目害虫温度效应解毒酶影响机制的研究却未见系统报道。棉铃虫寄主种类多样,其发生地区分布广泛,有效控制棉铃虫种群的化学防治易受到如温度等外界因素的影响,棉铃虫发生地区地理跨度大且发生历期长,造成了其生境极大的温度差异,影响药剂防治效果。研究杀虫剂的温度效应并分析其相关影响机制。【拟解决的关键问题】研究四氯虫酰胺对棉铃虫生物活性的温度效应,测定不同温度下经四氯虫酰胺影响后的棉铃虫体内主要解毒酶的比活力,分析双酰胺类杀虫剂对棉铃虫温度效应的影响机制,为不同环境温度下有效防治棉铃虫提供理论依据。
1 材料与方法
1.1 材 料
1.1.1 供试昆虫
棉铃虫HelicoverpaarmigeraHübner种群来源于河北省农林科学院植物保护研究所室内长期饲养种群,在(25±1)℃、60%~70%RH和光暗比16 h∶8 h(L∶D)条件下以人工饲料饲养。挑选生理状态一致,健康活泼的3龄幼虫供试。
1.1.2 供试药剂
94.1%四氯虫酰胺原药tetrachlorantraniliprole,沈阳科创化学品有限公司。
1.2 方 法
1.2.1 试验设计
试验所有步骤均在温控室内进行,可调温度范围(10±0.5)℃~(35±0.5)℃。温控室设有5个温度,15、20、25、30和35℃,将试虫提前24 h置于各个温度下饲养以适应温度。将四氯虫酰胺原药用丙酮稀释并配制成一定浓度的母液,再用0.1%吐温80水溶液将其稀释为5~7个浓度梯度备用。采用浸叶法测定四氯虫酰胺对棉铃虫幼虫的生物活性。将甘蓝叶片用打孔器打成直径1 cm的小圆片后,完全浸渍于药液中30s,取出后于吸水纸上自然晾干后将提前饥饿2 h的试虫接入其中。0.1%吐温80水溶液作为对照。每个处理4次重复,每个重复15头试虫,72 h后检查试虫死亡情况,以用镊子轻触虫体无正常反应为死亡标准。
1.2.2 解毒酶比活力测定
1.2.2.1 试虫处理
取生物活性测定得到的各温度下的0.1×LC50值为作为亚致死浓度,以0.1%吐温80水溶液为对照,在各温度下进行药剂处理,其余步骤同1.2.1生物活性测定方法。72 h后将试虫挑入1.5 mL的离心管中,液氮速冻后保存至-80℃下备用。
1.2.2.2 酶源制备
按每0.04~0.06 g昆虫加1 mL缓冲液的比例,向装有试虫的离心管中加入预冷的缓冲液,冰浴条件下充分研磨至匀浆,并在4℃、10 000 g条件下离心15 min,取上清液,混匀,冰浴待用。不同解毒酶选用不同的缓冲液,0.04 mol/L pH 7.0磷酸缓冲液用于CarE,0.05 mol/L pH 7.5 Tris-HCL缓冲液用于GST,0.1 mol/L pH 7.8磷酸缓冲液用于MFO,0.1 mol/L pH 7.6磷酸缓冲液用于UGT。
1.2.2.3 比活力
羧酸酯酶(CarE)比活力测定方法参照冉春等[16];谷胱甘肽S-转移酶(GST)参照Clark等[17];多功能氧化酶(MFO)参照Kim等[18];尿苷二磷酸糖基转移酶(UGT)参照娄琳琳[19]和王梦瑶[20]。
1.2.2.4 酶源蛋白质
参考Bradford[21]考马斯亮兰蓝G-250染色法测定酶源蛋白含量。
1.3 数据处理
使用DPS v6.55分析所得数据,得到各温度下的生物活性回归方程及LC50等参数。温度系数记为各温度与最低温度的LC50值比值的绝对值,其中LC50随温度升高而降低,则在温度系数之前标“+”,反之则为“-”。各温度与最低温度下95%置信区间不重叠则温度系数被认为具有显著性差异。
用Ducan’s多重比较分析不同温度下各处理解毒酶比活力差异显著性。
2 结果与分析
2.1 四氯虫酰胺对棉铃虫生物活性的温度效应
研究表明,四氯虫酰胺在不同温度下对棉铃虫的生物活性具有显著差异,在15、20、25、30和35℃,其LC50分别为326.91、59.18、28.91、9.07和0.26 mg/L,15与35℃间LC50相差高达1 257.35倍。与15℃时相比,从20~35℃,其温度系数由+5.52增长为+1 257.35,表现为极显著的正温度系数杀虫剂。表1
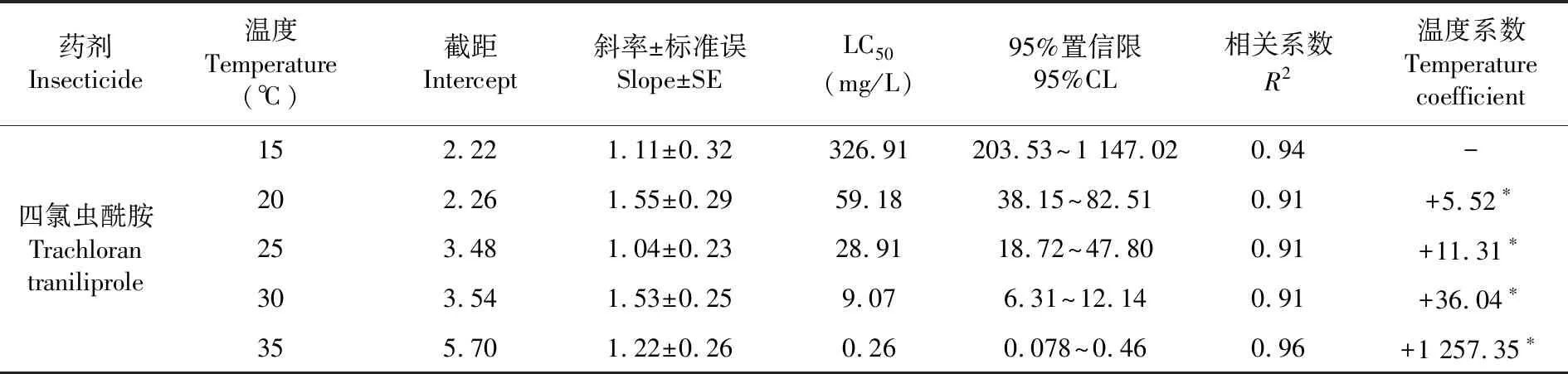
表1 四氯虫酰胺对棉铃虫生物活性的温度效应Table 1 Temperature effect on the bio-activity of tetrachlorantraniliprole to H. armigera
2.2 不同温度下四氯虫酰胺处理后棉铃虫体内解毒酶比活力的变化
2.2.1 羧酸酯酶(CarE)
研究表明,不同温度下棉铃虫CarE比活力均明显高于对照。对照组CarE比活力在15℃最高为94.98 nmol/mg Pr.min而后随温度增长比活力下降,经药剂处理后,CraE比活力在15~30℃由110.67 nmol/mg Pr.min不断增长至172.81 nmol/mg Pr.min,而在35℃出现下降趋势,降至133.63 nmol/mg Pr.min。表2,图1
2.2.2 谷胱甘肽S-转移酶(GST)
研究表明,在供试温度15~35℃,对照组GST比活力表现出一定的差异。在25℃时GST比活力达到最高为18.08 nmol/mg Pr.min,与其他温度下的比活力相比差异较为显著,出现明显的“钟型”曲线现象。经四氯虫酰胺处理后,GST比活力仅在15℃时略高于对照,为15.81 nmol/mg Pr.min,之后随着温度上升,其比活力迅速下降,20~35℃的GST比活力之间未出现显著差异,但均显著低于15℃。表2,图1
2.2.3 多功能氧化酶(MFO)
研究表明,随着温度升高,对照组MFO比活力未出现显著变化。经四氯虫酰胺处理后,MFO比活力均明显低于对照,其比活力在15℃最高为620.55 nmol/mg Pr.30 min,从15到30℃间出现明显下降趋势,而随温度升高,其比活力从484.28 nmol/mg Pr.30 min(30℃)升至571.22 nmol/mg Pr.30 min(35℃)。表2,图1
2.2.4 尿苷二磷酸糖基转移酶(UGT)
研究表明,对照组的UGT比活力在15~30℃呈不断上升趋势,30℃时达到4.43ΔOD/mg Pr.20 min。而经四氯虫酰胺处理后,UGT比活力明显高于对照,在15~20℃和30~35℃间呈下降趋势,30℃下的比活力为8.43ΔOD/mg Pr.20 min,显著高于其他温度。表2,图1

表2 四氯虫酰胺处理后棉铃虫体内4种解毒酶比活力变化Table 2 The variation of tetrachlorantraniliprole to the specific activities of four detoxification enzymes of H. armigera
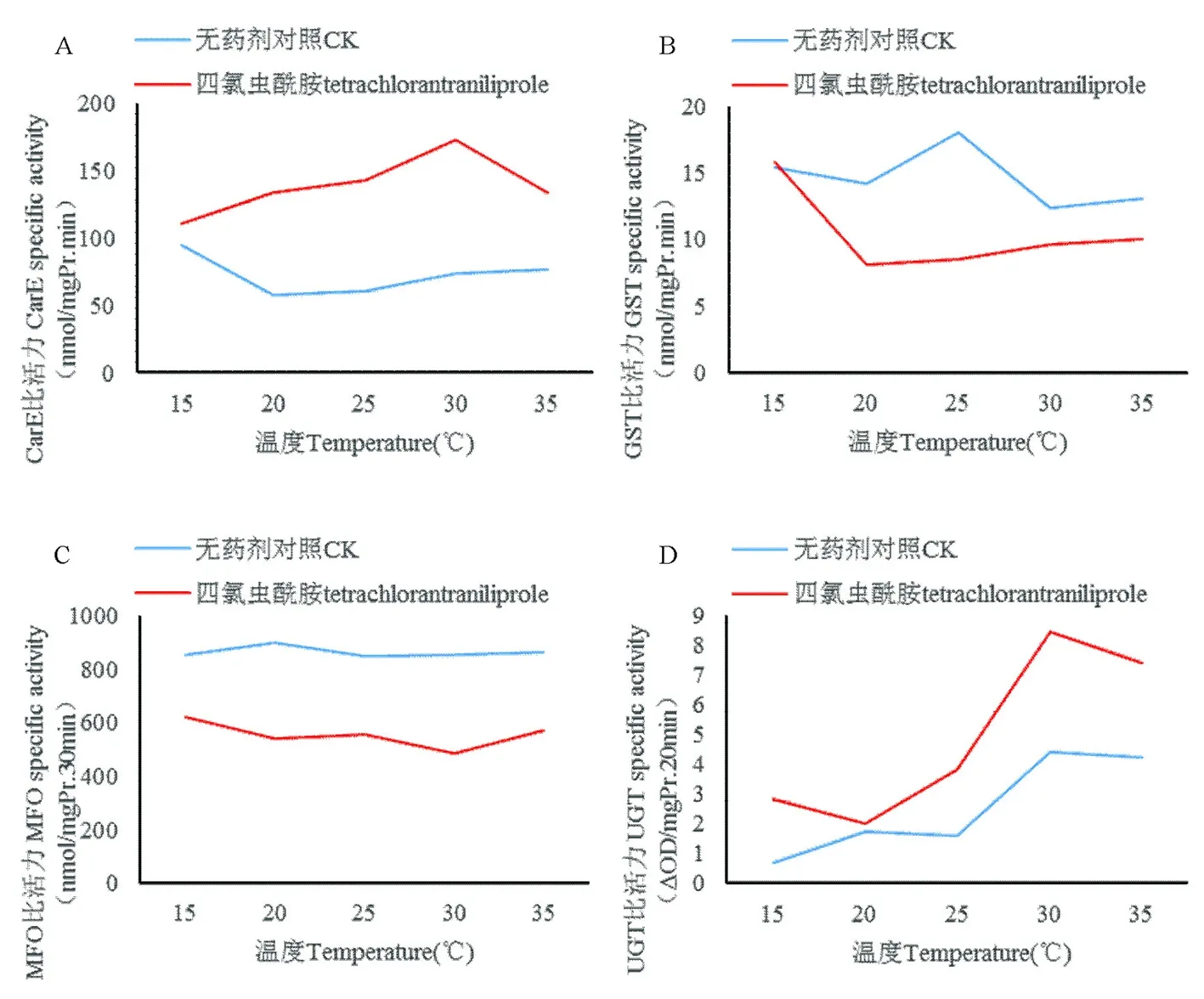
注:四氯虫酰胺处理后4种解毒酶比活力变化,其中A:CarE,B:GST,C:MFO,D:UGT
3 讨 论
温度对杀虫剂生物活性的影响因子是多元的,药剂在不同温度下的挥发程度,在昆虫表皮的渗透速度以及昆虫体内的解毒代谢均能引起杀虫剂生物活性的差异[5]。有关于温度和杀虫剂对昆虫的相关研究多为单一因素影响,而将三者相结合的研究相对较少且未成一定体系,不同昆虫以及不同药剂所表现的温度效应以及解毒酶等影响因素不尽相同。经短期高温处理后,33.5℃均能提高烟粉虱、蚜虫和黄曲条跳甲的多种酶活性,而36℃则表现为抑制作用;对于毒死蜱抗性品系小菜蛾,在33.5℃下,其各种酶活力未受到显著抑制,相较于33.5℃,36℃却显著抑制了其细胞色素P450含量[22]。小菜蛾经四氯虫酰胺亚致死浓度处理后,酯酶(EST)和MFO较对照组活性下降,但却对GST表现为诱导作用;氯虫苯甲酰胺处理家蚕24 h后,P450和GST活性显著升高[23-24]。经35℃高温预处理后,烟粉虱对噻虫嗪的敏感性增强,后经解毒酶验证发现,高温通过影响P450活性降低烟粉虱对噻虫嗪的耐受性[25]。当杀虫剂和温度同时作用于昆虫时,马云华[9]和刘佳[26]的研究分别发现了GST活性受诱导参与了拟除虫菊酯和溴虫腈对绿盲蝽的负温度效应的变化过程;安静杰[27]在有关于绿盲蝽转录组测序分析中发现,经正温度系数杀虫剂吡虫啉亚致死浓度处理后,在35℃下绿盲蝽体内P450、CarE和过氧化氢酶(CAT)相关基因差异表达显著,对于负温度系数杀虫剂高效氯氰菊酯则在15℃更为显著,UGT某些相关基因也参与其中。
四氯虫酰胺对棉铃虫表现出了显著的正温度效应,而从这4种解毒酶的比活力来看,在不同温度下经四氯虫酰胺处理后,分别出现了不同的变化趋势。随着温度的升高棉铃虫UGT比活力在30℃时达到最高,GST比活力在则25℃下出现峰值,这与刘佳等[15]在对绿盲蝽温度效应的研究结果中提到的“钟型”曲线相似。经四氯虫酰胺亚致死剂量处理之后,15~35℃间GST和MFO比活力明显低于对照,但GST在较低温度15~20℃间的比活力随温度升高显著降低,同时四氯虫酰胺对棉铃虫的LC50在此温度范围内由326.91 mg/L降至59.18 mg/L,两者变化趋势相符;MFO比活力在15~30℃出现下降现象,虽不及GST下降趋势明显但也同样符合毒力随温度上升而增强的趋势。CarE和UGT可能参与了四氯虫酰胺的部分代谢,比活力较对照组显著增强,但在15~30℃间总体与温度表现为正相关,而与四氯虫酰胺生物活性的温度效应未表现出明显相关性,CarE和UGT虽然可能在对四氯虫酰胺的解毒代谢中起到一定作用,但却受温度影响不明显。
4 结 论
四氯虫酰胺对棉铃虫表现出的正温度效应影响机制复杂,其正温度效应可能与GST和MFO有关,其中GST在15~20℃间比活力变化尤为显著。而CarE和UGT则未发现与四氯虫酰胺生物活性的正温度效应相关。
猜你喜欢
分子催化(2022年1期)2022-11-02
农产品质量与安全(2022年4期)2022-08-24
医学概论(2022年4期)2022-04-24
临床与实验病理学杂志(2021年10期)2021-12-13
商品与质量(2021年31期)2021-08-20
中国科技纵横(2021年24期)2021-03-02
农民致富之友(2019年2期)2019-03-16
科技与创新(2018年2期)2018-01-09
小学科学(2015年9期)2015-09-28
科学种养(2015年2期)2015-02-05
