
尖笔帽螺(Creseis acicula)研究进展及其在大亚湾暴发机制初探*
2021-11-02 01:08齐占会史荣君戴明韩婷婷黄洪辉
热带海洋学报 2021年5期
齐占会, 史荣君, 戴明, 韩婷婷, 黄洪辉
1. 中国水产科学研究院南海水产研究所, 广东省渔业生态环境重点实验室, 农业农村部外海渔业开发重点实验室, 广东 广州510300;
2. 南方海洋科学与工程广东省实验室(广州), 广东 广州 511485
1 尖笔帽螺的生物生态学特征
1.1 分类地位
尖笔帽螺(Creseis acicula, Rang 1828)隶属软体动物门(Mollusca)、腹足纲(Gastropoda)、后鳃亚纲(Opisthobranchia), 翼足目(Pteropoda)、龟螺科(Creseidae)、笔帽螺属(Creseis)的海洋浮游软体动物。该种为大洋性暖水种营浮游生活, 是中国黄海和东海浮游软体动物群落的优势种(张福绥, 1966;徐兆礼, 2008; 胡剑 等, 2008)。
1.2 形态学特征
尖笔帽螺软体部可分为4部分: 头、足、内脏团和外套膜(图1)。头部有一对触须和眼, 口的两侧有一对翼足, 用于游泳和收集食物, 这种翼足结构是该种类最典型的形态学特征。尖笔帽螺在游泳时摆动翼足, 像蝴蝶煽动翅膀, 因此又称为海蝴蝶(sea butterfly)。在笔帽螺属中, 该种的个体最大。尖笔帽螺壳细长, 呈针形, 一般呈笔直状, 但也有的个体壳尖端向左或右略弯。大亚湾尖笔帽螺暴发期间所采集的样品壳长范围为4.13~14.72mm(9.60±2.17mm), 与印度洋海域和日本海域尖笔帽螺的壳长相似(9~12mm)(Nishimura, 1965; Pillai et al, 1979)。壳宽范围为0.25~1.27mm (0.86±0.25mm),体湿重范围为2~3.8mg (2.8±0.6mg)。尖笔帽螺的壳表面平滑, 顶端部位具缢勒两个, 前方者不太明显,贝壳无色透明或呈浅黄褐色。
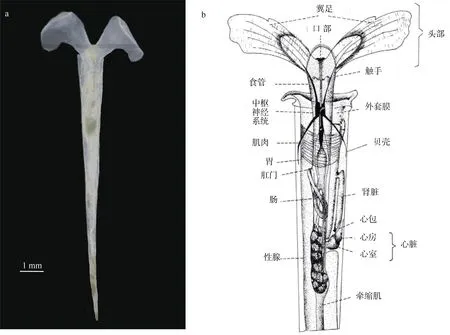
图1 尖笔帽螺形态特征a. 大亚湾尖笔帽螺样品照片; b. 示意图[参照Trégouboff 等(1957)]Fig. 1 Morphological characteristics of Creseis acicula. (a) picture taken in July 2020 in Daya Bay; (b) after Trégouboff et al (1957)
1.3 摄食与消化
尖笔帽螺的摄食方式为滤食性, 通过翼足上纤毛的摆动捕获和收集硅藻、甲藻和微型甲壳动物等有机颗粒物, 进入口部的食物通过颌的运动输送到口腔前庭或咽部, 咽部生有带状齿舌, 齿舌上附生有很多微小的齿。食物经由食管进入胃部, 胃部有一列咀嚼板结构用于磨碎食物, 胃部连接弯曲的肠道, 经消化吸收后的食物残渣最终从位于身体中部的肛门排出(Herman, 1998)。
2 尖笔帽螺的分布特征
2.1 地理分布及其影响因素
尖笔帽螺在印度洋、太平洋和大西洋等海域均有分布, 主要生活在500m以浅水层(Albergoni,1975)。世界上多处海域都曾有尖笔帽螺暴发的记录, 其中以印度洋暴发的次数最多, 包括Karwar、Goa、孟加拉湾等(Krishnamurthy, 1967; Peter et al,1978; Naomi, 1988), 此外, 在西北太平洋日本海中部新泻岛海域(Nishimura, 1965; Morioka, 1980)、墨西哥湾(Hutton, 1960), 以及大西洋地中海(Kokelj et al, 1994)也都有报道。
温度和盐度是影响尖笔帽螺地理分布最重要的环境因素, 尖笔帽螺其密度沿温带地区向热带地区递增(Herman, 1998)。由于尖笔帽螺的体液渗透压受所海水盐度的影响, 因此其地理分布范围受海水盐度的限制。尖笔帽螺等翼足类在大西洋、印度洋和太平洋的温暖水域分布最为广泛, 而在红海等蒸发量较大, 表层盐度高于40‰的海域生存的翼足类种类仅有其他海域的一半左右(Herman et al, 1969)。地中海的盐度低于红海而高于外海大洋, 该海域的翼足类种类约为同一纬度具有相似温度海域的75%左右。另一方面, 在黑海以及某些河口等低盐度水域, 翼足类分布也很少(Herman,1998)。这些现象都表明, 盐度是影响翼足类分布的重要因素。
2.2 尖笔帽螺在中国海域的分布
尖笔帽螺在中国的黄海、东海和南海均有分布,是很多海域浮游软体动物群落的优势种。该种为暖水性种类, 适温下限为15~20℃, 数量随着纬度的增加而显著减少, 分布范围随季节变化明显, 在黄海水域分布的最北限为34°N, 在31°—34°N海域出现6个月, 表现为春、夏季随水温的上升, 其分布逐渐向北扩展, 8—9月达到最北, 在秋、冬季伴随着水温的下降其分布又逐渐向南回缩, 伸缩的范围超过6~7个纬度; 在28°—31°N海域全年当中出现8个月(张福绥, 1966)。该种在东海南部的丰度高于北部, 外海高于近岸, 丰度与温度正相关, 台湾暖流是决定丰度的重要因素(Xu et al, 2006b)。盐度方面, 尖笔帽螺在黄海海域基本上沿着33‰~34‰的等盐线分布, 虽然有时也会扩展到32‰的水域范围, 但不会到盐度更低的区域中去(张福绥,1966)。
在黄东海北部和南部海域尖笔帽螺的丰度分别为0.1个·m-3和0.52个·m-3(张福绥, 1966)。在长江口海域尖笔帽螺是浮游软体动物的优势种, 其生物量占浮游软体动物总量的55%, 平均丰度为0.6个·m-3, 最高值约4.2个·m-3(胡剑 等, 2008)。在福建及台湾海峡海域尖笔帽螺也是浮游软体动物群落的优势种, 数量最高达1.32个·m-3(黄加祺 等,1993)。南海中部海域尖笔帽螺在秋季数量达到0.08个·m-3(戴燕玉, 1995)。在南海北部的大亚湾水域, 根据农业部南海区渔业生态环境监测中心2011—2019年间采用垂直拖网方法(网口直径0.5m,网孔505μm)开展的浮游动物调查结果, 3—11月都发现了尖笔帽螺, 但种群数量维持在较低水平, 大多低于0.50个·m-3, 仅在2012年6月在个别站位发现2.0个·m-3的高值。可见虽然尖笔帽螺是中国很多海域数量最丰富的浮游软体动物之一, 但其数量此前从未达到暴发成潮的水平。
3 大亚湾尖笔帽螺的暴发
3.1 尖笔帽螺的暴发过程
2020年6月12日在大亚湾西南部大亚湾核电厂和岭澳核电厂(以下简称核电站)的冷源取水口附近发现尖笔帽螺成群聚集, 甚至在风浪的推动下堆积在岸边(图2)。农业部南海区渔业生态环境监测中心开展了应急监测, 采用垂直拖网(网口直径0.5m, 网孔505μm)的方法对尖笔帽螺的数量进行了调查, 发现尖笔帽螺的密度高值区集中在西南部沿岸区域, 尤其是岭澳核电站冷源取水口附近, 但在大亚湾东部、北部以及湾口区域均未发现尖笔帽螺暴发。7月4—8日为暴发的高峰期, 尖笔帽螺平均密度超过1000个·m-3,核电站冷源取水口附近站位平均密度约1851个·m-3;7月5日尖笔帽螺的数量达到最高峰, 最高值约5600个·m-3; 7月9日后尖笔帽螺的密度明显下降,低于100个·m-3; 7月14—17日在海底沉积物中采集到了尖笔帽螺死亡后的空贝壳, 3天内空壳的密度从120个·m-3增加至1020个·m-3, 说明有大量个体死亡后沉降到海底; 7月17—19日, 通过海上密目拖网日夜走航调查, 并同步进行声学探测, 未发现尖笔帽螺及其声学信号, 表明尖笔帽螺潮已经消退。本次大亚湾尖笔帽螺暴发从发现到消退的持续时间约1个月, 是中国海域尖笔帽螺种群暴发的首次记录。

图2 大亚湾尖笔帽螺暴发现场照片a. 在大亚湾海水中聚集成群漂浮; b. 在大亚湾岸边堆积Fig. 2 Creseis acicula bloom in Daya Bay. (a) C. acicula population floated in the bay; (b) C. acicula accumulated along the bank
3.2 大亚湾尖笔帽螺的暴发机制探讨
大亚湾海域3—11月都有尖笔帽螺出现, 可以推测本次暴发的尖笔帽螺应是在大亚湾本地繁殖生长的, 而非随海流漂流至此的。我们知道在自然条件下任何种群数量的增长和维持都受到很多环境和生物因素的限制, 包括适宜的温度、盐度、食物的可利用性、竞争者、捕食者、自身的繁殖能力和生长潜力等。只有在多个因素都适宜的情况下, 种群才有可能出现暴发性增殖。在此对尖笔帽螺在大亚湾暴发的可能机制做如下初步分析和探讨。
1) 温度。尖笔帽螺属于大洋性暖水种(Herman et al, 1969), 在温度较高的季节和区域出现较多(黄加祺 等, 1993; 戴燕玉, 1995; 罗文增 等,2016)。尖笔帽螺在大亚湾暴发期间, 其密度高值区分布在大亚湾核电站周边水域, 并以之为圆心向外递减, 高值区均分布在水温高于30℃的范围内。已有研究显示大亚湾核电站冷却水的热排放导致排水口局部区域水温升高6.8℃, 使此区域温度常年高于30℃, 甚至高达35℃ (Song et al, 2004; Li et al, 2011), 显著高于周围水域, 并导致大鹏澳部分区域的表层海水温度与1991年核电站建设前相比升高了1~3℃ (Tang et al, 2003)。这提示我们核电站冷却水排放造成的升温效应值得注意, 水温的升高可能是尖笔帽螺暴发的诱发因素之一。
2) 盐度。尖笔帽螺属广盐性种类, 在外海、沿岸和河口区域, 如长江口(Xu et al, 2006a, b; 徐兆礼, 2008)都有分布, 表明其能够耐受较低盐度和盐度在一定范围内的波动。2020年5—6月大亚湾区域曾有大量降雨导致海水盐度骤降。对此我们可以假设此时的海水盐度或许并不是尖笔帽螺所最适宜的, 但在其耐受范围之内, 而与之具有相似生态位存在饵料竞争的种类及其捕食者, 则可能因为无法适应盐度的降低者剧烈波动而大量死亡。竞争压力和捕食压力显著降低, 使尖笔帽螺的幼体有更大的存活率, 并发育生长为成体, 从而在6月份形成种群的暴发。
3) 食物可利用性。尖笔帽螺是滤食性种类, 其食物包括浮游动、植物和其他颗粒有机物。食物的数量、质量和可利用程度是决定其种群增长和维持的重要条件。根据大亚湾核电站附近水文浮标的监测数据, 6月初水体叶绿素a的质量浓度从1mg·m-3升高至约5mg·m-3, 说明这段时期水体中浮游植物的数量很高, 可能为尖笔帽螺的暴发提供了营养支持, 至7月下旬高密度种群对食物的大量消耗是其迅速消退的重要原因。
值得注意的是, 已有很多研究表明核电站温排水区域的浮游生物群落组成和优势种都发生了显著改变。核电站冷却水出水口区域微型浮游植物的比例降低, 并且相对于硅藻, 水温的升高更有利于鞭毛藻类的生长(Li et al, 2011)。但我们发现在本次尖笔帽螺暴发期间硅藻仍是浮游植物群落的优势种, 一定程度上反映了环境因子对浮游植物影响的复杂性。此外, 温度也是影响浮游动物丰度变化的主要因素(Wang et al, 2012)。Li等(2014)发现核电站温排水口区浮游动物的密度显著高于其他区域, 并与温度呈正相关关系。鸟喙尖头蚤(Penilia avirostris)成为浮游动物群落的优势种, 并且在核电站排水口区域出现最高的丰度值, 认为水温升高是其种群暴发的主要原因之一。另外, 核电站冷却水的升温效应还改变了水体CO2体系, 促进藻类的生长(彭云辉 等, 2001), 而使水体中颗粒有机物数量增加。基于这些发现, 我们可以推测核电站冷却水排放导致局部海域海水温度升高, 对尖笔帽螺的食物可利用性能够产生两方面的影响, 一是温升效应促进了浮游植物生长, 提高了总的颗粒有机物数量;二是改变了浮游生物群落结构, 有可能使尖笔帽螺所喜爱和易于取食的种类数量增加。这些都能为尖笔帽螺种群的暴发提供必须的营养基础。
4 尖笔帽螺暴发的生态效应
自然界任何物种数量的暴发都会对生态系统造成多方面和深远的影响。目前关于尖笔帽螺暴发的生态效应还知之甚少, 我们初步分析可能包括以下几个主要方面:
1) 尖笔帽螺对浮游生物的大量捕食消耗, 一方面会显著降低浮游生物的数量, 尤其是粒径较小的生物的数量, 另一方面其选择性捕食可能会导致浮游生物群落结构的改变; 2) 局部水域超高密度的个体呼吸作用迅速消耗水体的溶解氧, 造成暂时性的低氧区; 3) 大量个体死亡后沉降到海底, 其软体部有机质降解会大量消耗上覆水中的溶解氧, 造成底层缺氧, 沉积物中有机碳及其他元素的早期成岩反应路径改变; 4) 尖笔帽螺死亡后大量碳酸钙质的贝壳沉降到海底, 形成翼足类动物软泥, 改变暴发区海域沉积物理化性质。翼足类的外壳主要由文石构成(Seibel et al, 2007), 比方解石更易溶解于海水中(Mucci et al, 1983)。它们的外壳会比颗石藻(coccoliths)和有孔虫的外壳更快的溶解, 但在具有较高温度、物质循环较慢和沉降速率较高的海盆区域, 会形成碳和钙在很长时间尺度上的沉积与埋藏(Roger et al, 2012)。
5 总结与展望
尖笔帽螺是全球海洋广泛分布的浮游软体动物, 目前对其的研究主要是围绕种属鉴定、地理分布、古生物学和古地质学等方面开展的, 其种群暴发的生理生态机制还不清楚。本次尖笔帽螺在大亚湾的暴发, 高密度区均分布在核电站冷源周围水域, 让我们很自然的将之与核电站冷却水温升效应及其所引发生态效应联系起来,推测可能与尖笔帽螺的暴发有关, 但就目前所掌握的数据资料还不足以确定两者之间是否真的存在关联, 仍需深入研究核电站温升效应对局部水域生态系统的综合影响, 尤其是对尖笔帽螺饵料丰度、捕食者和竞争者数量等方面的影响, 结合暴发期间的水质水文特征, 进行综合分析才能明确。
猜你喜欢
当代水产(2022年6期)2022-06-29
装饰装修天地(2019年9期)2019-10-21
天津诗人(2018年3期)2018-11-13
新少年(2018年10期)2018-10-22
农业与技术(2017年16期)2017-09-16
科技创新导报(2016年24期)2017-03-13
红领巾·萌芽(2016年4期)2016-11-19
红岩春秋(2016年8期)2016-05-14
小天使·五年级语数英综合(2014年6期)2014-07-29
知识窗(2013年7期)2013-05-14
