
海洋微生物来源天然产物研究现状与态势
2021-11-02 01:07马丽丽田新朋李桂菊赵晏强殷建平
热带海洋学报 2021年5期
马丽丽, 田新朋, 李桂菊, 赵晏强, 殷建平
1. 中国科学院武汉文献情报中心, 湖北 武汉 430071;
2. 科技大数据湖北省重点实验室, 湖北 武汉 430071;
3. 中国科学院南海海洋研究所, 广东 广州 510301
海洋微生物、微型藻类以及海洋无脊椎动物是海洋天然活性物质的三大主要来源。海洋环境的特殊性(如高压、高盐、缺氧、低温、黑暗、贫营养等)及海洋生物物种间复杂广泛的生态作用赋予了海洋微生物有别于陆生生物的新陈代谢途径及适应机制,从而产生结构独特、丰富多样、生物活性显著的次级代谢产物, 是新药来源的重要基础, 被认为是天然药物资源的新宝库(柏凤月 等, 2016)。近年来, 从海洋微生物中发现新的海洋天然产物数量呈明显上升趋势。2018年报道的海洋天然产物有55%(857/1554)来源于海洋微生物(Carroll et al, 2020)。从海洋微生物中分离得到的活性物质涵盖了萜类、甾醇类、生物碱类、甙类、多糖、肽类、核酸、蛋白质、酶类等, 很多物质具有抗菌、抗病毒、抗肿瘤、抗心脑血管疾病、抗氧化、抗炎、酶及酶的抑制活性、抗寄生虫、抗过敏反应等作用, 这些先导化合物的发现为药物研究提供了重要的资源。本文从海洋真菌/细菌来源天然产物的研究现状、海洋微生物天然产物的人工合成、海洋微生物天然产物的生物活性评估以及成药性评价等方面进行了概述, 分析了当前海洋微生物天然产物研究的态势。
1 海洋微生物天然产物研究现状
海洋微生物天然产物主要来自海洋真菌和海洋细菌, 除了生存在水体及海底沉积环境中的海洋细菌和真菌外, 还包括很多与其他的海洋生物处于共生、附生等关系的细菌和真菌群体。如图1所示, 通过检索海洋真菌和海洋细菌天然产物研究相关的SCI论文情况可知(来自Web of Science数据库, 检索时间截止到2020年3月16日), 国际上海洋微生物来源天然产物的研究始于20世纪90年代, 整体呈增长趋势, 尤其是近10年发展快速,反映出近年来世界各国对海洋微生物天然产物的研究与开发越来越重视。

图1 1992—2019年海洋真菌/细菌天然产物SCI论文发表情况Fig. 1 SCI papers published during 1992—2019 in the research field of natural products from marine fungi or marine bacteria
从研究国家分布来看, 在海洋真菌来源天然产物研究方面, 我国在该领域的SCI论文量排名首位, 其次美国、德国、日本、韩国也是海洋真菌天然产物领域的重要研发国家; 在海洋细菌来源天然产物研究方面, 我国的相关论文产出较少于海洋真菌来源天然产物, 稍逊美国排名第二位, 印度、德国、日本也是该领域的世界前五国家(图2)。总体来看, 中国、美国和德国3个国家海洋微生物天然产物论文发表数量占全球的54.6%。
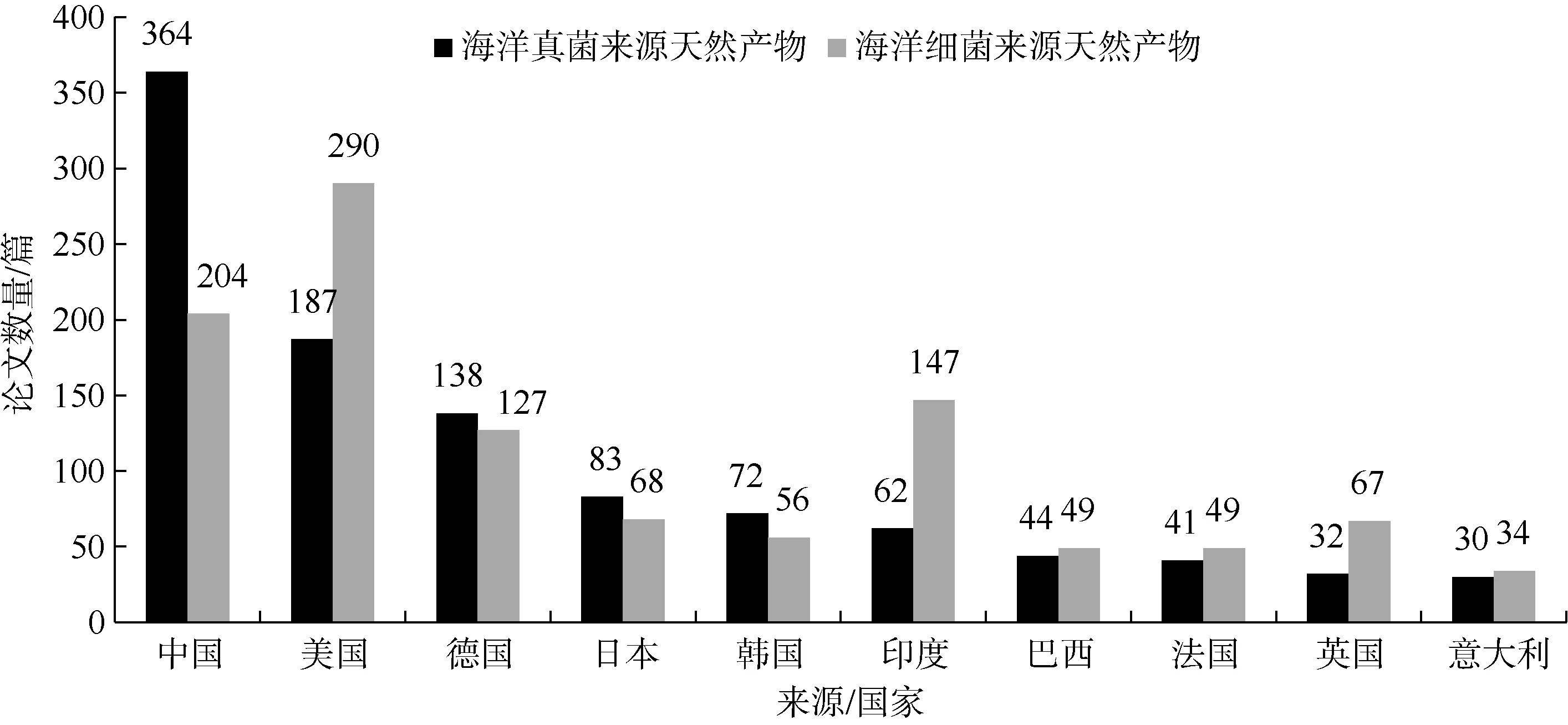
图2 海洋真菌/细菌天然产物研究领域的主要国家(前10)Fig. 2 Top-10 countries in the research field of natural products from marine fungi or marine bacteria
从各国相关论文的年份分布情况看, 美国、德国、印度、日本开展海洋微生物天然产物研究的时间较早。相比之下, 我国的海洋微生物天然产物研究起步较晚, 主要起始于近10年, 但近10年里我国在该领域的研究发展较快, 数量上一直领先美国等其他国家(图3)。其中, 中国科学院、中国海洋大学、中山大学、沈阳药科大学、浙江大学等在我国的海洋微生物天然产物研究领域发挥了重要作用。

图3 1992—2019年海洋微生物天然产物研究领域重要国家的年度发文数量情况无黑点的位置表示该年度论文发文量为零; 论文发文量多少与图中黑点大小成正比Fig. 3 Annual numbers of papers published by top-10 countries in the research field of natural products from marine microorganisms during 1992—2019
从高频作者关键词来看, 海洋真菌和海洋细菌, 尤其是海洋放线菌(如链霉菌)是海洋微生物天然产物的热门来源, 其中聚酮、二酮哌嗪、肽类是出现频次较多的化合物类型。海洋微生物天然产物的生物活性(包括抗菌活性、细胞毒性、抗真菌活性、抗癌、抗肿瘤、抗污、抗生物膜活性、抗氧化活性等)、全合成、生物合成(如生物合成基因簇、聚酮合酶合成途径)、基因组挖掘、代谢组学、结构解析、药理学和药物发现等关键词也出现频次较多, 也侧面反映出这些内容是当前海洋微生物天然产物领域的重要研究主题(表1)。另外, 通过科学知识图谱VOSviewer工具绘制了该领域论文关键词的平均发表年份分布图谱(图4), 可知微生物组、生物合成基因簇、基因组挖掘、代谢组学等是该领域的研究前沿和热点, 气相色谱-质谱联用技术(Gas chromatography-mass spectrometry, GC-MS)、单菌多次级代谢产物(One-Strain-Many-Compounds,OSMAC)策略、异源表达、共培养策略、质谱技术等是该领域的热门研究技术, 抗菌素耐药性、抗氧化活性、抗炎活性、治疗阿尔茨海默病等是海洋微生物天然产物生物活性研究方面的热点。
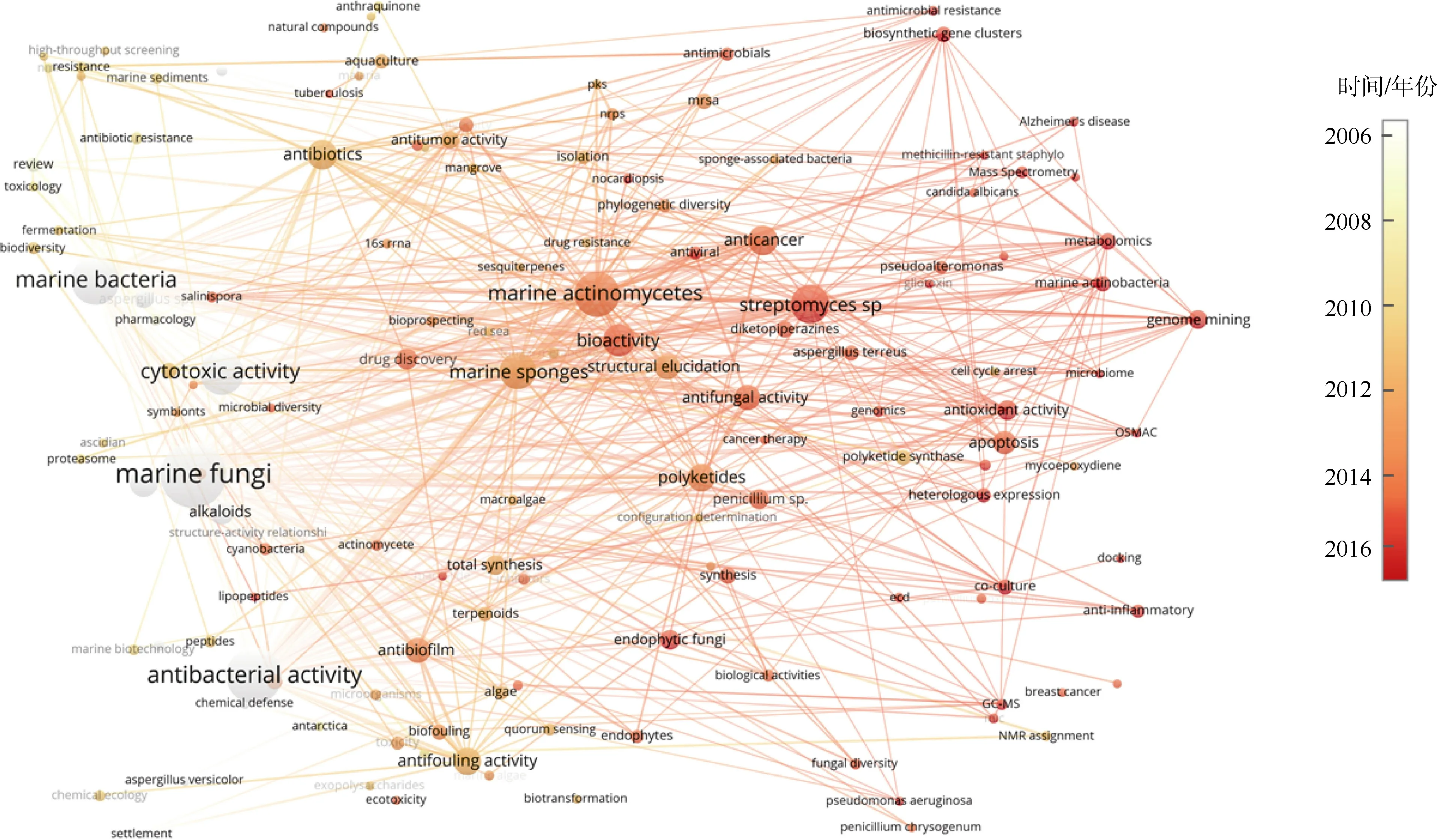
图4 2006—2016年海洋微生物天然产物领域论文关键词(出现频次≥5)的平均发表年份分布图中文字大小跟该关键词出现的频次成正比, 出现频次越多, 文字越大; 反之亦然Fig. 4 Distribution of average publish year of keywords (occurrences ≥ 5) in the research field of natural products from marine microorganisms during 2006—2016

表1 海洋微生物天然产物研究领域高频关键词(前30)Tab. 1 Top-30 high-frequency keywords in the research field of natural products from marine microorganisms
1.1 海洋真菌来源天然产物
海洋真菌的次生代谢产物由于具有结构多样、生物活性丰富、“创新指数”高、产量大等特点, 已经成为化学、生物学、药学等领域关注的焦点(金黎明 等, 2016)。近几年来新发现海洋真菌活性次级代谢产物的数目也随之在持续增加, 《海洋天然产物》2020年报告最新报道, 2018年海洋真菌来源代谢产物新增617个(Carroll et al, 2020)(图5), 与2017年相比增加了38%, 而与前3年平均数量相比增加了85%。因此, 报告中预测如果依这种增长趋势发展, 海洋真菌天然产物的数量(目前为4708种)可能超过腔肠动物, 在2021年将成为海洋天然产物的第二大来源, 到2024年将成为最大的来源。海洋真菌天然产物的来源可分为5个方面: 1) 海洋动物来源真菌天然产物, 主要包括在海洋动物内部组织中生活的内生真菌、在海洋动物体表附着的附生真菌、在海洋动物肠道、外骨骼等处寄生的真菌3类; 2) 红树林来源真菌天然产物, 这是海洋真菌的最大类群之一; 3) 海藻来源真菌天然产物, 目前报道的海藻真菌约占全部海洋真菌的三分之一; 4)海水来源真菌天然产物; 5) 海洋沉积物来源真菌天然产物, 一般为泥沙微粒(金黎明 等, 2017)。
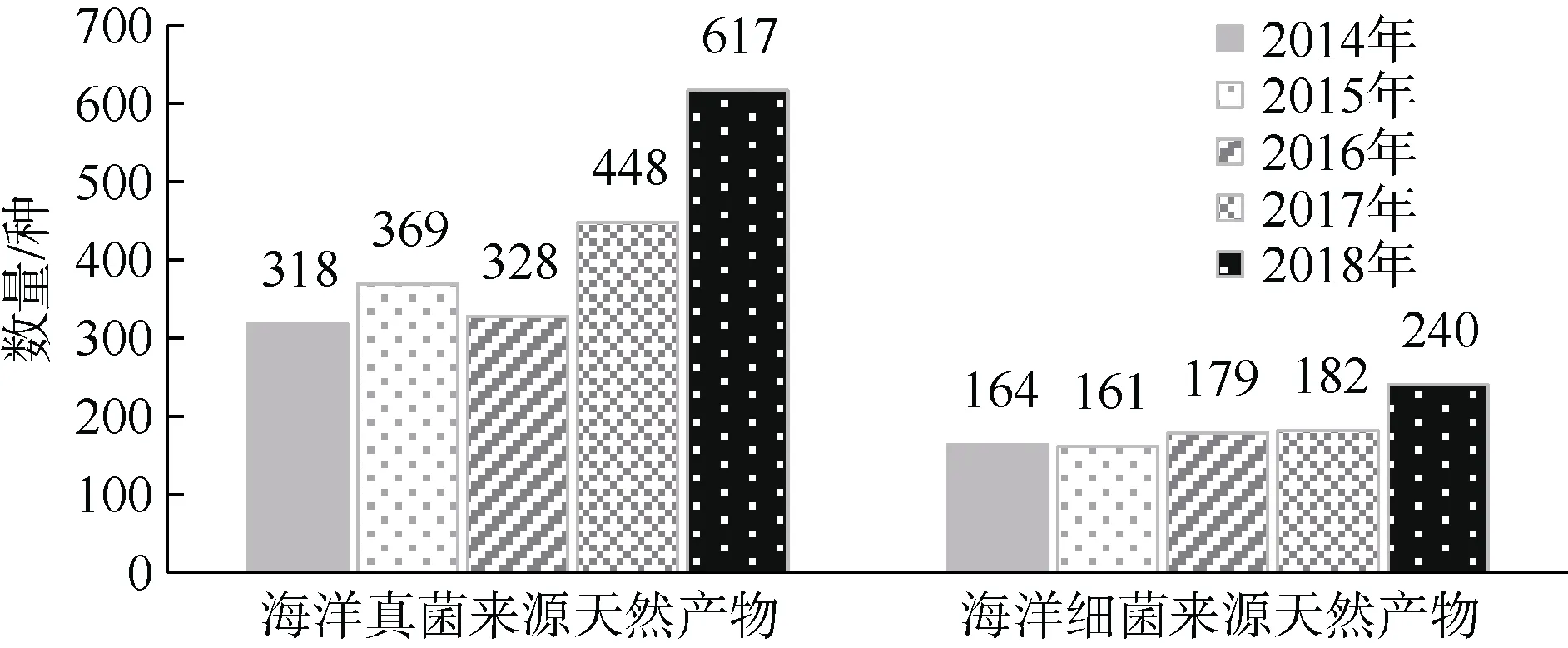
图5 2014—2018年海洋真菌/细菌来源天然产物数量新增趋势Fig. 5 Trends of new natural products from marine fungi or marine bacteria during 2014—2018
在研究过的约几千种海洋真菌种类中, 研究最多的是青霉属(Penicillium)和曲霉属(Aspergillus)。据不完全统计, 自1991年Kobayasht等报道第一个海洋青霉菌来源的天然产物fellutamide A到2014年, 共报道了392个海洋青霉菌来源的新天然产物(马红光等, 2014)。自1992年Shinggu等首次报道来自海洋曲霉属的新天然产物fumiquinazolines A-C 以来,海洋曲霉属来源的新天然产物数量也在逐年增加,至今已发现506个海洋曲霉属来源的新天然产物。这些海洋青霉属和曲霉属来源的天然化合物结构包括聚酮、生物碱、萜类、甾体、卤代物、脂肪酸、肽类、糖苷等多种类型, 表现出抗菌、自由基清除、抗寄生虫、抗癌等多种生物活性。除此之外, 至2014年已报道的除青霉属和曲霉属以外的124种其他海洋真菌来源的新天然产物1133个, 其中我国学者是曲霉属和青霉属以外其他真菌来源的海洋新天然产物的主要发现者(赵成英 等, 2014; 朱统汉 等, 2014)。
海洋真菌来源天然产物的生物活性涉及抗菌、抗炎、抗病毒、抗肿瘤、抗污损和酶抑制活性等。其中对人类病原菌如金黄色葡萄球菌(Staphylococcus aureus)、大肠杆菌(Escherichia coli)、枯草芽孢杆菌(Bacillus subtilis)、绿脓杆菌(Pseudomonas aeruginosa)、白色念球菌(Monilia albican/canidia Albicans)等抗菌活性的研究较多(金黎明 等, 2017)。另外, 近年来相关研究表明海洋真菌来源天然产物表现出抗植物病原菌活性。农用生物活性较好的海洋真菌主要来源于红树林和深海沉积物, 其中曲霉属和青霉属真菌具有较好的抗植物病原菌和卤虫致死等活性, 在生物农药研发、环境保护等领域具有重要的研究意义和广泛应用前景, 但大多数研究仍处在实验室研究阶段, 实践应用的还较少(王聪 等, 2018; 王丹等, 2019; 付兵 等, 2020)。近年来不断有研究报道从支顶孢属(Acremonium)、链格孢属(Alternaria)、曲霉属(Aspergillus)、假头状孢子属(Pseudotrichosporon)、毛壳属(Chaetomium)、分支孢子菌属(cladosporiosis)、地霉属(Geotrichum)、镰刀属(Fusarium)、青霉属(Penicillium)、盘多毛孢属(Pestalotia)、茎点霉属(Phoma)、葡萄穗霉属(Stachybotrys)、帚霉属(Gliocladium)、木霉属(Trichoderma)、腐质霉属(Humicola)等海洋真菌中分离得到抗肿瘤活性物质, 其表现出的对人白血病、肺癌、肝癌、乳腺癌、胃癌、组织细胞性淋巴癌、宫颈癌、胰腺癌、结肠癌、前列腺癌等的细胞毒性使海洋真菌天然产物成为海洋医药的研究热点(文雯 等, 2019; Wali et al, 2019)。其中由海洋曲霉属来源的天然产物halimide衍生而来的plinabulin (NPI-2358)已进入Ⅱ期临床研究, 用于治疗非小细胞肺癌,成为20个海洋药物之一(赵成英 等, 2014)。Deshmukh等(2018)总结了近年来海洋真菌中分离得到的199种天然产物有很大潜力发展成为抗癌药物的化合物, 可针对多种特定癌症细胞系表现出良好的细胞毒性。
1.2 海洋细菌来源天然产物
海洋细菌是海洋中分布最广、数量最多的一类微生物群体。海洋细菌大多是从海洋沉积物、海水、海藻、海洋动物体表等分离获得, 主要集中在假单胞菌属(Pseudomonas)、弧菌属(Vibrio)、微球菌属(Micrococcus)、芽孢杆菌属(Bacillus subtilis)、肠杆菌属(Enterobacter HormaecheandEdwards)和互生单胞菌属(Domonas)。《海洋天然产物》2020年报告最新报道, 2018年海洋细菌来源的新增天然产物为240个(图5), 呈持续显著增长趋势(Carroll et al,2020)。海洋细菌次生代谢产物可分为生物碱、黄酮类、苯丙素类、甾醇类、多糖类、多肽类等, 不同的海洋细菌产生的活性物质种类以及各化合物比例存在一定差异, 活性研究方面主要涉及到抑菌和抗肿瘤活性(Carroll et al, 2020; 李慧玲 等, 2020)。
海洋放线菌在海洋细菌天然产物中研究最多,海洋沉积物、海绵和红树林等多种海洋资源中都蕴含着丰富的放线菌, 这些海洋放线菌具有独特的代谢途径, 能够产生结构新颖活性良好的代谢产物,在新药开发中蕴藏着巨大的潜能(赵婷婷 等,2018)。目前海洋沉积物来源放线菌仍是研究重点,已报道的产活性化合物的海洋放线菌约有一半分离自海洋沉积物。海洋放线菌在抗癌药物的研究方面较海洋真菌、海绵等起步晚, 但有研究表明, 海洋放线菌的代谢产物中蕴藏着结构较新奇的天然产物,为抗肿瘤药物提供先导化合物, 显示出巨大的发展潜能, 如20S蛋白酶体抑制剂Salinosporamide A(NPI-0052)于2012年进入临床试验, 2015年就被美国食品药品监督管理局(Food and Drug Administration,FDA)批准为治疗多发性骨髓瘤的孤儿药(Marizomib)(Niewerth et al, 2014)。另有很多研究报道了海洋动植物共附生的放线菌能够产生具有结构多样的抗菌、抗肿瘤等活性物质。
最常见且研究最多的海洋放线菌为链霉菌属(Streptomyces)。从1976年报道首例海洋链霉菌来源的aplasmomycins A-C至2016年, 一共报道了547个海洋链霉菌新天然产物(王聪 等, 2016)。2017年海洋放线菌链霉菌属新增天然产物137个, 2018年继续新增167个(Carroll et al, 2019, 2020)。海洋链霉菌来源天然产物结构类型呈现多样性, 涉及生物碱等含氮化合物(约61%)、聚酮(约35%)、卤代物(约10%)、萜类(约7%)等, 67.3%的化合物表现出肿瘤细胞毒、抗菌、抗疟、抗寄生虫, 以及糖苷酶抑制等生物活性。调研结果显示, 中国在海洋链霉菌天然产物方面的研究论文排名首位, 美国、印度、德国、韩国、日本等国也是海洋链霉菌天然产物的主要贡献者(王聪 等, 2016)。近年来, 海洋拟诺卡菌(Nocardiopsis)由于其独特的代谢功能, 成为海洋放线菌次级代谢产物的重要来源, 对拟诺卡菌的研究仅次于链霉菌, 至2019年已报道海洋拟诺卡菌来源天然产物67个, 结构包括吡喃酮类、二酮哌嗪类、多肽类等, 其中约46%是吡喃酮类化合物(王聪 等,2019)。另外, 主要分布于热带和亚热带海洋沉积环境中的盐孢菌属(Salinispora)是首个被报道的专性海洋放线菌。截止2017年, 从盐孢菌属沙栖盐孢菌(Salinispora arenicola)、太平洋盐孢菌(Salinispora arenicola)、热带海洋盐孢菌(Salinispora arenicola)中分离出共30种新颖次级代谢产物, 其中盐孢菌素A (NPI-0052)是抗肿瘤显著的明星化合物(王可欣等, 2018)。
2 海洋微生物天然产物的化学合成与生物合成
2.1 海洋微生物天然产物的化学合成
骨架新颖、活性独特的海洋天然产物是药物先导化合物的重要来源, 但由于海洋样品难以重复获得、活性成分含量极其低微, 难以通过提取分离手段大量获得, 加之海洋活性天然产物结构复杂, 难以大量提取, 使得许多高活性海洋天然产物无法进行后续的生物学活性和成药性评价以及结构改造优化, 一直是海洋新药研发的瓶颈之一。另外, 直接从海洋微生物中提取出的海洋天然产物, 可能会出现特异性生物活性差异, 甚至毒副效果大或者吸收不完全、代谢不充分、长期使用产生耐药性等不合理的药代动力学, 因此需要对其进一步人工修饰才能作为药物进行临床使用。还有一些化合物本身不宜直接开发, 但是其具有良好的生物活性, 可以将这些海洋天然次级代谢产物作为先导化合物, 以供研究人员对进一步优化和改造其结构, 从而研发出后续系列产品, 这些化合物所具有的活性基团往往也可以作为活性先导结构, 为研制创新型药物提供重要模型。
现代有机合成尤其是海洋天然产物全合成的迅猛发展, 特别是对具有生物活性和药理作用的海洋天然次级代谢产物的发展起到重要作用, 为获取复杂小分子海洋活性天然产物提供了重要手段。探究海洋天然产物全合成的新策略和新方法, 建立高效、高选择性的规模化制备技术, 是国际海洋药物研究的热点。乔梁等(2015)概述了6个已被批准上市的小分子海洋药物的全合成研究, 分别是阿糖胞苷(Cytarabine, Ara-C)、阿糖腺苷(Vidarabine,Ara-A)、曲贝替丁(Ecteinascidin-743, ET-743)、脱氢膜海鞘素(Dehydrodidemnin B, Didemnin B)、甲磺酸艾日布林(Eribulin)、海兔毒素(Dolastatins)类化合物。其中, 抗肿瘤和抗病毒药物Ara-C和Ara-A都是以海绵核苷类化合物为先导进行结构优化, 采用全合成技术而获得的药物分子。另外, 目前Ara-A虽然已经可以通过人工合成大量得到或者通过酶催化大量生产, 但目前临床用药则主要来源于陆生链霉菌—抗生链霉菌(Streptomyces antibioticus)培养液中提取。
海洋天然产物立体构型往往与其生物活性息息相关, 因此确定立体构型是海洋天然产物领域的另一个研究难点和重要内容。彭吉星等(2015)综述了目前海洋天然产物立体构型确定的常用方法及其应用研究案例, 例如借助量子化学计算的方法解决了一株海绵来源真菌——黄曲霉(Aspergillus flavus)中分离到的环匹阿尼酸衍生物speradine C的立体构型问题。目前确定海洋天然产物绝对构型的方法包括基于手性试剂的核磁共振法、光谱学方法、参考生物合成或生物信息学方法、X 线单晶衍射方法、有机合成方法、量子化学计算方法等。但在化合物构型解析方面没有一种通用的方法能够解决所有化合物的结构问题, 即使是综合应用波谱学和化学合成方法, 目前仍有相当多的结构无法确定。
尽管有机合成化学在海洋微生物天然产物中已取得较大进展, 但在结构复杂、多手性中心的天然产物全合成中也普遍存在一些问题, 具体表现在合成路线长、产率低、需要昂贵试剂或苛刻的反应条件, 难以实现大量合成或生产等。近年来出现的合成生物学正好弥补了这些问题。
2.2 海洋微生物天然产物的生物合成
天然产物的生物合成是在基因和蛋白功能水平上认识和理解复杂天然产物的生物合成机制, 以化学、生物学知识为指导, 通过合理设计在发酵友好的微生物中实现生物合成途径的重构, 进而通过系统优化实现目标化合物的规模化合成。天然产物化学合成的主要优势在于结构的可变性, 而生物合成的主要优势在于过程的经济性, 两种策略的融合可综合两方面优势, 为复杂海洋微生物天然产物的高效合成提供了新的思路。
2.2.1 海洋微生物天然产物生物合成基因簇或元件研究
研究发现许多微生物基因组中含有多个次级代谢产物生物合成基因簇, 在海洋微生物基因组中,大约10%的DNA序列编码次级代谢产物的生物合成, 许多放线菌8%~10%的基因组序列与次级代谢相关(侯路宽 等, 2017; 鞠建华, 2017)。但常规实验室培养条件下, 许多次级代谢产物生物合成途径是处于沉默状态的, 因此需要通过激活一些沉默基因簇和次级代谢产物代谢途径, 激活海洋微生物的天然产物生物合成潜能。目前合成基因簇激活方法有:1) OSMAC是一种激活次级代谢产物沉默基因簇的传统方法, 主要是通过改变培养基成分、改变发酵条件、添加不同小分子化合物等常规实验室培养条件来影响次级代谢产物的生物合成途径, 包括相关基因转录、蛋白质翻译、酶抑制或者酶激活, 但OSMAC策略改变代谢途径的准确机制仍没有被完全了解; 2) 共培养策略是模仿多种微生物共存的生态环境, 在实验室条件下共同培养两种或者多种不同的菌株, 使菌株因竞争或对抗而刺激产生在常规培养条件下无法获得的沉默基因簇产物; 3) 转录因子调控主要是通过表达正向调控因子或者敲除负向调控因子来激活沉默基因簇表达; 4) 表观遗传调控是在不改变DNA序列的情况下, 主要通过DNA甲基化、组蛋白修饰等途径进行沉默基因簇的激活; 5)异源表达是通过把沉默的基因簇转移到大肠杆菌、酵母等异源表达寄主中来诱导沉默基因表达, 有研究已证实这是一种可获得一些新的天然产物的成功策略(李雪, 2017; 赵江华 等, 2020; 马新华 等,2020)。
2.2.2 海洋微生物天然产物生物合成途径研究
同陆生微生物相似, 目前已经获得的绝大多数的海洋微生物的代谢途径仍然基于Ⅰ型聚酮合酶(Polyketide synthase, PKS)、Ⅱ型聚酮合酶和非核糖体肽合成酶(Non-ribosomal peptide synthetase, NRPS)以及PKS/NRPS杂合途径。因此海洋微生物次级代谢产物的生物合成主要源于4种途径: 聚酮合酶途径, 非核糖体肽合成酶途径, 聚酮-非核糖体肽合成酶杂合途径, 以及其他途径, 如钱士云等(2019)发现海洋长孢葡萄穗霉菌中双吲哚化合物的生物合成途径主要通过色氨酸途径并且受色氨酸合成基因的调节。近年来研究多集中于聚酮合酶和非核糖体肽合成酶体系负责的模块化生物合成领域(黄婷婷 等,2018)。
2.2.3 组合生物合成技术
海洋微生物次级代谢产物生物合成基因簇及其代谢途径的阐明, 为组合生物合成技术在海洋微生物次级代谢产物中的应用奠定了基础。组合生物合成技术通过将克隆的生物合成基因簇进行异源表达,从而快速、大量获取海洋来源的天然产物(庞博 等,2015)。因此, 虽然起步较晚, 组合生物合成技术在海洋微生物天然产物研究中得到了较多关注并取得了一些重要成果, 很多海洋微生物产聚酮类或聚肽类化合物基因组已被克隆(陈俊升 等, 2009)。例如,研究人员已经能够采用纯酶学手段在体外重建肠球菌素和改造Patellamides的生物合成途径, 为利用合成生物学手段改造天然产物提供了参考和元件库。Salinosporamide A是海洋专有放线菌——热带盐水孢菌(Salinispora tropica)通过PKS/NRPS杂合途径合成的化合物, 研究人员利用前体添加、基因失活、基因重组和化学酶法等组合生物合成技术获得了20多个salinosporamide A的结构类似物(肖吉等, 2012)。鞠建华等2016年研究发现了5-磷酸吡哆醛(pyridoxal 5'-phosphate, PLP)连接的转氨酶和一个新的异构酶在两个链霉菌中的非蛋白质氨基酸l-allo-Ile天然产物生物合成中的协同作用 (Li et al,2016)。
生物合成技术推动了海洋微生物天然产物领域的快速发展, 但也仍存在一些问题。例如, 如果微生物生物合成基因簇较大, 则会增加生物合成途径重构的复杂性, 得到的表达系统可能达不到预期效果。另外, 目前海洋微生物天然产物生物合成还处于初级发展阶段, 存在通量低、错误率高、自动化程度低、与传统工艺相比经济优势不明显等一些问题, 且目前对生物合成过程中的一些重要代谢途径和酶作用机制了解不足, 这些都限制了海洋微生物天然产物生物合成的广泛应用和快速发展。最后,合成生物学等技术在海洋微生物天然产物生物合成中的应用也不可避免的引起人们对其可能带来的生物安全风险产生担忧, 比如改造微生物的环境释放或泄漏对自然环境生态系统和人类健康方面的影响等, 相关的生物安全风险防控和监管措施值得进一步考虑(王丽苹 等, 2017; 杨菊 等, 2017; Gómez-Tatay et al, 2019)。
3 海洋微生物天然产物的活性评估
海洋微生物天然产物的活性评估主要是指测定其生物活性物质的抗氧化、抗病原细菌、抗真菌、抗病毒、抗肿瘤等的能力以及其生物酶、功能肽等生物活性, 从而筛选出有价值的活性物质。基本流程可概括为: 试验材料或样品采集➙选择合适方法分离、纯化和筛选目标微生物, 活性菌种的鉴定➙活性菌种的发酵以及化学分离➙活性物质的筛选➙活性物质某种抗性的活性测定➙抗肿瘤等活性成分鉴定➙单体化合物的活性验证。在某些情况下, 为了研究不同基团对化合物生物活性的影响, 需要进行如酰基化、羟基化、卤化和糖基化等合成后修饰。另外, 目前不仅仅是海洋微生物来源的天然产物,一般情况均是对新化合物进行的活性评价, 很少对已知化合物做系统的活性评价, 这种已知化合物活性评价不系统的情况下, 会漏掉重要活性的化合物,这种情况应该引起大家重视。
3.1 抗肿瘤活性评价
在海洋天然产物多种生物活性研究中, 海洋抗肿瘤活性物质的研究在海洋天然产物研究领域一直占据重要地位。抗肿瘤活性评价通常包括两个步骤:1) 初筛: 利用体外实验检测受试化合物对肿瘤细胞生长状态的影响。细胞活性测定是常用的方法,采用酚酞蓝细胞排染法、细胞计数法(Cell Counting Kit-8, CCK-8)、噻唑蓝[3-(4,5)-dimethylthiahiazo(-z-y1)-3,5-di-phenytetrazoliumromide, MTT]比色法、磺酰罗丹明B (Sulforhodamine B, SRB)比色法等方法测定化合物对肿瘤细胞生长抑制能力, 从而测定追踪抗肿瘤活性成分, 该方法能高效、准确的分离纯化抗肿瘤活性物质, 减少盲目性(刘光甫 等, 2016;谢春兰 等, 2017)。2) 确证实验: 利用实验性动物肿瘤模型观察体内抗肿瘤作用, 并通过细胞分子生物学方法探讨抗肿瘤作用机制。相对来说, 海洋微生物天然产物抗肿瘤活性的药理学研究大多仅停留在体外对肿瘤细胞毒性作用的初步筛选, 能够开展深入机制研究以及体内实验的还较少。
3.2 抗氧化活性评价
抗氧化活性评价是指测定天然产物对自由基的清除能力或者对脂类物质氧化的抑制能力, 既要测定其体外抗氧化能力, 又要考虑其在生物体内的抗氧化效果。曾维才等(2013)和张雪等(2017)等系统介绍了天然产物抗氧化活性的常用评价方法, 可为海洋微生物天然产物抗氧化活性评价提供参考, 概括起来可包括: 1) 羟基自由基清除法, 主要有脱氧核糖法、邻二氮菲法等; 2) 超氧阴离子清除法, 主要有NBT (Nitro-blue tetrazolium, 氮蓝四唑)法、AP-TEMED (过硫酸铵/N, N, N', N'-四甲基乙二胺)体系法、邻苯三酚法等; 3) 基于电子转移途经的自由基清除法, 主要有ABTS [2,2'-azinobis-3-ethylbenzthiazoline-6-sulphonate, 2,2'-联氮-二(3-乙基-苯并噻唑-6-磺酸)二铵盐]自由基阳离子清除实验、DPPH (1,1-diphenyl-2-picrylhydrazine, 1,1-二苯基-2-三硝基苯肼)自由基清除实验等; 4) 基于氢原子转移途径的自由基清除法, 主要有氧自由基吸收能力(Oxygen Radical Absorbance Capacity, ORAC)测定法、化学发光(Photochemiluminescent, PCL)法、总自由基清除抗氧化能力(Total Peroxyl Radical -trapping Antioxidant Parameter, TRAP)法、DMPD (N,N-Dimethyl-p-phenylenediamine, N, N-二甲基-对苯二胺)法等; 5) 抑制自由基产生能力活性测定方法, 主要有络合金属离子检测方法, 如铁离子络合能力、铜离子络合能力、磷钼络合物法等; 6) 碘量滴定法的脂质氧化测定实验; 7) 细胞抗氧化活性评价方法,常用的有细胞抗氧化活性(cellular antioxidant activity, CAA)测定法、MTT法、ATP (adenosine triphosphate)测定法等; 8) 以小鼠等动物模型为基础的体内抗氧化评价方法。
3.3 抗菌活性评价
如前文所述, 抗菌活性是海洋微生物天然产物活性评价研究中较活跃的领域之一。K-B (Kirby-Bauer)纸片法是世界卫生组织规定的一般抗菌药物筛选标准方法, 该方法使用的初筛菌株通常包括金黄色葡萄球菌(G+)、藤八迭球菌(G+)、大肠杆菌(G-)、绿脓杆菌(G-)等。例如尚卓等(2012)采用滤纸片法分析了一株海洋放线菌唐德链霉菌(Streptomyces tendae)的抗菌活性, 该菌株对枯草芽孢杆菌和金黄色葡萄球菌具有显著抗菌活性。对于抗厌氧菌药物筛选, 目前尚无公认的标准方法, 多依据实际情况采用微量液体稀释法筛选抗变形链球菌药物, 或琼脂稀释法筛选抗幽门螺旋杆菌药物。季宇彬等(2019)建立并完善了一种改良双层琼脂板筛选法,通过共培养可从海洋真菌中快速筛选并发现抗菌活性菌株。
3.4 抗病毒活性评价
鉴于流感病毒的高变异性及耐药性和现有抗流感化学合成药物有限的瓶颈问题, 结构新颖、药理活性多样的海洋微生物次级代谢产物是寻找抗流感病毒新药的一条重要途径。20世纪50年代, 来源于海洋真菌——产黄顶孢霉(Acremonium chrysogenum)的头孢菌素C是第一个从海洋微生物中获得并成功开发为药物的化合物, 成为后来头孢菌素系列抗生素的先导化合物。细胞病变法(cytopathic effect, CPE)是常用的天然产物体外抗病毒活性测定方法。彭吉星(2015)采用CPE法对H1N1病毒感染狗肾上皮细胞进行了初步体外抗病毒活性筛选, 再通过高效液相色谱-紫外-质谱联用(High Performance Liquid Chromatography-UV-Mass Spectrometry, HPLC-UVMS)方法化学筛选和菌株种属的排重, 最终选取了5株具有较强抗甲型流感活性的海洋真菌菌株进行活性产物研究。另外, 陈飞敏(2019)采用MTT实验来评价红树林真菌娃儿藤内生真菌(Diaporthesp.)SCSIO 41011与海生真菌CochlioboluslunatusSCSIO 41401的次级代谢产物的细胞毒性和不同亚型流感病毒的体外抗病毒活性筛选。采用蛋白质免疫印迹、间接免疫荧光和实时荧光定量PCR方法检测代谢产物Spirostaphylotrichin X对甲型流感病毒复制的影响, 并采用假病毒抑制实验、血凝抑制实验、神经氨酸酶抑制实验、时间过程分析实验和病毒RNA聚合酶抑制实验研究了其对流感病毒复制周期的阻断作用。
3.5 酶/酶抑制活性评价
酶活性的测定方法按反应时间通常可分为3种:1) 定时法: 又称中止反应法, 在酶反应一定时间后,通过加入强酸、强碱、蛋白沉淀剂等使反应完全停止, 通过测定酶反应开始后某一时间段内产物或底物浓度的总变化量来计算酶反应初速度; 2) 连续监测法: 又称动力学法或速率法、连续反应法。在酶反应过程中, 用仪器持续监测某一反应产物或底物浓度随时间的变化从而计算出酶反应初速度; 3) 平衡法: 又叫终点法, 通过测定酶反应开始至反应达到平衡时产物或底物浓度总变化量来计算酶活力。采用的监测方法有分光光度法、旋光法、荧光法、电化学方法、化学反应法等。例如董佩佩等(2018)利用分光光度计比色法对从海洋动物、海水、底泥等分离出的产中性蛋白酶、淀粉酶活性菌株进行了测定。酶抑制活性的测定主要是通过观察天然产物对酶活性的影响, 根据酶的特点, 酶的反应底物、产物都可以作为检测指标, 采用的方法与酶活性的测定方法类似。例如史倩等(2019)建立了基于高效液相色谱-紫外(HPLC-UV)和高效液相色谱-多级离子阱质谱分析(High Performance Liquid Chromatography-Ion Trap Mass Spectrometry / Mass Spectrometry,HPLC-ITMS/MS)的海洋天然产物体外抑制环氧合酶2 (Cyclooxygenase 2, COX-2)的活性评价方法。
4 海洋微生物天然产物成药性评价
药理活性和成药性是新药创制的两大要素, 然而大多数天然产物自然条件下不能完全满足制药要求, 需要进行结构改造, 因此新药研发的过程很大程度上就是成药性优化。目前海洋微生物天然产物的成药性研究还较少, 郭跃伟等(2016)开展了10种海洋生物来源候选药物的成药性评价, 其中5种是海洋微生物来源的天然产物, 分别是海洋微生物来源的生物碱HDN-1和WHJ-64、降二萜化合物wbg、变构甾体HDZ-137以及混源生物碱FGFC1。王媛媛(2012)对从海洋红树林根部海泥的真菌灰绿曲霉(Aspergillus glaucus) HB1-19中提取的一个结构新颖的蒽环内酯类化合物——灰绿霉素A(AspergiolideA, ASP-A)的抗肿瘤成药性进行了早期评价, 通过小鼠肝癌H22和人肝癌BEL-7402移植瘤模型对ASP-A的体内药效学进行评价, 采用HPLC方法对ASP-A进行了非临床药代动力学研究。曾志平(2019)运用基于结构的药物虚拟筛选技术, 从5969个海洋天然产物库中初步得到了10个能很好地靶向寨卡病毒非结构蛋白3 (Nonstructural protein-3, NS3)解旋酶的化合物, 并利用薛定谔软件套装中的药物吸收、分布、代谢、排泄及毒性预测模块(Absorption, Distribution, Metabolism, Excretion and Toxicity, ADMET)对这些化合物展开了初步的成药性评价, 认为可能成为潜在治疗寨卡病毒病的海洋天然产物。其中3个来自海洋微生物, 分别是2010年波兰科学家从海洋细菌——弧菌(Vibriosp.)DS40M4分离得到的Divanchrobactin, 以及2008年我国科学家从海洋真菌白曲霉中分离得到的芳香聚酮类化合物Aromatic polyketides和从青霉菌中分离得到的Dimeric Terrestrols。
5 小结与讨论
由于当前新药研发的瓶颈以及海洋科技和天然产物研究的发展, 近10年来国内外在海洋微生物来源天然产物研究方面发展迅速。但与海洋其他来源天然产物相比, 目前对海洋微生物天然产物的研究仍明显较少。据国际海洋微生物普查(International Census of Marine Microbes, ICoMM)计划研究人员保守估计, 海洋微生物至少有2000万种, 丰度总量可达1030数量级(Qiu, 2019; 刘宇航 等, 2020), 目前研究和鉴别过的海洋微生物种类微乎其微, 其中用于人类药物开发的更是少之又少。海洋微生物天然产物的发展缓慢主要是受研究技术方法的限制,因此提高海洋微生物分离技术、优化海洋微生物培养技术、发展海洋微生物高通量筛选技术、加强基因编辑、高通量测序、宏基因组、生物信息学等新一代生物技术的应用, 在海洋微生物获取及其天然产物资源开发中至关重要。
从来源类型来看, 目前海洋微生物天然产物的来源多集中在海洋真菌(海洋青霉属和曲霉属最多)、海洋放线菌(链霉菌属最多), 其他海洋微生物菌属研究较少, 应拓展海洋微生物天然产物的新来源, 充分开发海洋微生物天然产物资源。技术方法上, 需要加强发展海洋微生物天然产物半合成和全合成技术, 加强海洋微生物天然产物次级代谢产物代谢途径研究, 降低合成过程中的盲目性, 优化次级代谢产物合成和发酵工艺。发展海洋微生物天然产物活性先导化合物生物合成技术, 阐明海洋微生物天然产物生物合成基因簇和合成途径, 研制新的发酵工艺或酶系统, 发展组合生物合成等技术在海洋微生物天然产物大规模生产中的应用。
从研究内容来看, 国内外海洋微生物天然产物的研究论文大多集中在海洋微生物的分离、培养、天然产物活性测定等方面, 大多处于抗肿瘤、抗菌、抗病毒等天然产物活性的初步评价, 研究内容模式套路化和程式化, 理论研究水平不高。相对于海洋微生物来源天然产物的药理活性评价来说, 天然产物的成药性评价方面的研究工作更少。在今后研究中, 应加强海洋微生物天然产物生物活性评价研究,建立新的、有效的药物筛选方法, 提高可成药性海洋微生物天然产物筛选效率, 以及动物模型实验和作用机制的深入研究。
从研究国家来看, 我国在海洋微生物天然产物领域论文量虽然领先, 但仍需认识到与世界发达国家在海洋微生物天然产物及其药物研发体制、技术、产品、产业化等层面上的差距。另外, 国内外海洋微生物天然产物研究领域均存在理论研究与转化严重脱节, 新发现的海洋微生物天然产物逐年增多,但可发展成药物的很少, 与临床疾病治疗的实际结合也有待加强。我国应建立国家层面、符合国际规范的海洋微生物天然产物及药物研发创新体系, 加大投资力度, 培育发展海洋微生物天然产物及药物相关新兴战略性产业, 鼓励产学研合作, 促进医药企业与高校、科研院所联合进行深层次的海洋微生物天然产物药物研发, 提升我国海洋微生物天然产物开发能力。
猜你喜欢
学苑创造·A版(2022年5期)2022-05-19
食品安全导刊(2021年21期)2021-08-30
现代临床医学(2021年2期)2021-03-29
英语文摘(2020年7期)2020-09-21
大自然探索(2019年1期)2019-01-24
今日财富(2017年32期)2017-10-19
大自然探索(2017年3期)2017-04-07
方圆(2016年15期)2016-09-14
方圆(2016年4期)2016-03-07
对联(2011年10期)2011-09-18
