
56份人工合成六倍体小麦的遗传多样性分析
2021-10-23 06:21:18王秋云冯雯杰张巧玲张宪军刘淑珍鲍印广
麦类作物学报 2021年8期
王秋云,冯雯杰,张巧玲,赵 凯,张宪军,刘淑珍,鲍印广
(1.济宁市农业科学研究院,山东济宁 272000; 2.济宁市任城区农业农村局,山东济宁 272113; 3.山东农业大学农学院作物生物学国家重点实验室,山东泰安 271008)
小麦(TriticumaestivumL.)是地球上种植最广泛的粮食作物,占人类消耗总能量的五分之一[1]。随着人口数量不断增加,全球气候变暖,耕地面积和水资源逐渐减少,选育高产、优质、抗病虫小麦品种的需求更加迫切[2-3]。由于长期集中利用少数优质育种亲本,导致现有育成小麦品种的遗传基础日趋狭窄,遗传多样性逐渐缩小[4]。因此,引进、鉴定、评价、利用国内外优异种质资源,对于丰富小麦种质资源的多样性、拓宽小麦品种的遗传基础具有重要意义。
目前,生产中种植的普通小麦(TriticumaestivumL,AABBDD)是由四倍体二粒小麦(Triticumdicoccoides,AABB)与二倍体粗山羊草(Aegilopstauschii,DD)天然杂交而成[5]。由于长期的自然或人工选择作用,其遗传多样性逐渐降低[6-7]。通过人工杂交和染色体加倍重现二粒小麦与粗山羊草之间的杂交,可以引入新的基因资源,重获小麦起源和驯化过程中失去的许多有价值的遗传变异。
四倍体小麦和粗山羊草的遗传多样性丰富,含有抗病、高产、优质等许多优良基因,例如在粗山羊草中鉴定出抗秆锈病基因Sr33和Sr41、抗叶锈病基Lr21、Lr22a、Lr32、Lr39、Lr42和Lr43、抗条锈病基因Yr24和Yr28以及抗白粉病基因Pm2、Pm19、Pm34和Pm35等[8-10]。它们含有与普通小麦相同的染色体基组,是普通小麦的二级基因源[11]。利用四倍体小麦与粗山羊草杂交并经染色体加倍产生的合成六倍体小麦,含有双亲全部原始的遗传变异,对小麦遗传改良具有重要利用价值。孔令让等[12]利用四倍体小麦与粗山羊草杂交,通过幼胚拯救获得的Ps5-Y287双二倍体(AABBDD),其具有细胞学稳定、免疫白粉病、高抗条锈病和叶锈病等特点。由于合成六倍体小麦的基因组与普通小麦基因组同源,利用其与现代小麦品种杂交,可以通过同源重组的方式将人工合成六倍体小麦的优良基因导入普通小麦品种中[13-14]。以人工合成小麦为桥梁亲本的育种方式可以实现整个基因组不同位点的渗入,对于产量等多基因控制的数量性状的改良具有显著作用。因此,利用人工合成小麦作为育种桥梁材料进行小麦品种的遗传改良,越来越受到育种家的重视[15-16]。
本研究拟对引自CIMMY的56份人工合成六倍体小麦进行主要农艺性状鉴定,对其高分子量谷蛋白亚基的构成及基因组遗传多样性进行分析,为其在普通小麦品种遗传改良中的有效利用提供参考。
1 材料与方法
1.1 供试材料
供试材料为引自国际玉米与小麦改良中心(CIMMYT)的56份人工合成六倍体小麦,编号Syn1-Syn56(表1),由国家小麦改良中心山东(泰安)分中心引进并保存。对照为5个当地栽培品种:济宁12、济麦22、良星99、临麦4号、郑麦7698。
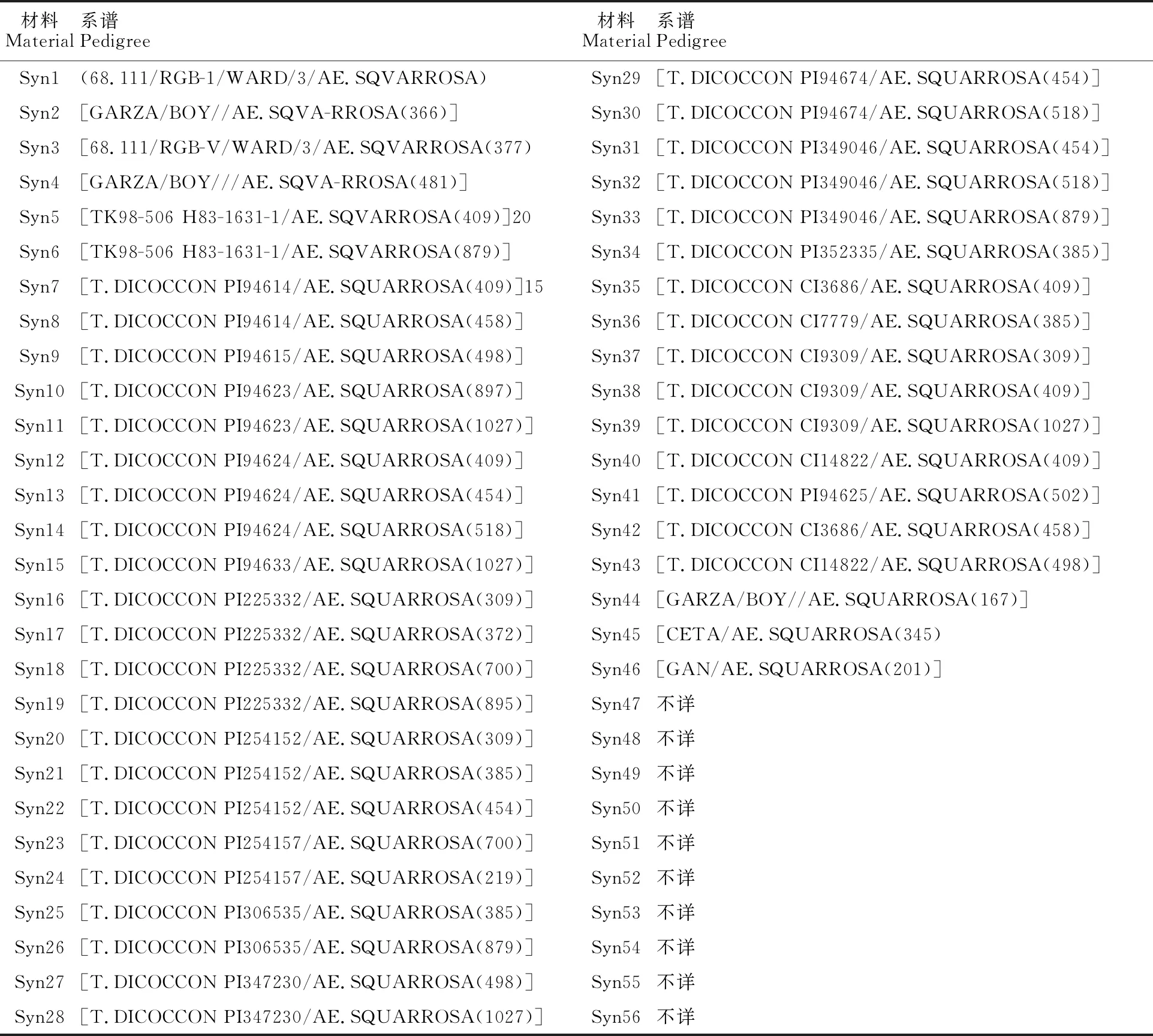
表1 56份供试人工合成六倍体小麦
1.2 试验方法
1.2.1 农艺性状调查
试验于2012 年10月至2019年6月在山东省济宁市农业科学研究院试验地进行,每个材料种植2行,行长3 m,株距30 cm,行距50 cm。其余管理按照正常田间管理进行。参照李立会、李秀全的《小麦种质资源描述规范和数据标准》[17],对供试材料的株高、穗长、单株穗数、小穗数、穗粒数、千粒重、籽粒颜色、籽粒形状、壳色、颖壳毛性等性状进行调查。Shannon-Wiener多样性指数(H`)采用张倩男等[18]方法计算。
1.2.2 白粉病抗性鉴定
于2014-2016年在济宁市农科院试验地常年有小麦白粉病发病地块,进行白粉病抗性鉴定。在田间自然发病条件下,以当地感病小麦品种济宁12为对照,按照盛宝钦[19]提出的0~4级分级标准记载反应型,鉴定标准为:免疫(0)、近免疫(0;)、高抗(1)、中抗(2)、中感(3)及高感(4)。
1.2.3 高分子量谷蛋白亚基分析
采用张学勇等[20]的方法,以马奎斯(1,7+9,5+10)和中国春(Null,7+8,2+12)为对照,利用十二烷基硫酸钠聚丙烯酰胺凝胶电泳(SDS-PAGE)技术对供试材料的HMW-GS组成进行分析。
1.2.4 遗传多样性分析
采用酚-氯仿法提取植物幼嫩叶片的DNA[21]。采用涵盖小麦基因组21条染色体的 2 492对多态性引物[22]对3个合成六倍体小麦(Syn2、Syn44、Syn54)和5个小麦栽培品种(济宁12、济麦22、良星99、临麦4号、郑麦7698)进行了多态性分析,筛选出20对条带清晰,多态性好的分子标记进行遗传多样性分析。扩增反应在TakaRa PCR thermaL cycler上进行,采用降落PCR(Touchdown PCR),即94 ℃变性4 min; 94 ℃变性45 s,65 ℃复性50 s(每个循环降低 1 ℃),72 ℃延伸55 s,15个循环;94 ℃变性40 s,50 ℃复性40 s,72 ℃延伸40 s,30个循环;最后72 ℃延伸5 min;扩增结束后10 ℃保存。将PCR产物电泳结果进行统计分析,计算各引物的等位变异数和多态性信息量(PIC,polymorphic information content)。
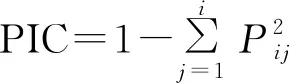
式中,Pij表示位点i的第j个等位变异出现的频率。PIC值的大小在0~1之间,0表示无多态性,1表示具有非常高的多态性。
1.3 数据处理
利用Excel 2007进行数据整理和分析。
2 结果与分析
2.1 主要农艺性状特点
调查结果(表2)表明,56份供试材料株高平均为112.07 cm,变化范围为90.00~132.00 cm,变异系数最小,为8.46%;穗长平均为15.3 cm,变化范围是11.0~22.5 cm;单株穗数平均为 20.4个,变化范围为5.0~45.4个,变异系数最大,为43.54%;每穗小穗数平均为17.7个,变异范围是11.6~21.6个;穗粒数平均为35.56粒, 变异范围22.20~67.80粒;千粒重平均为36.37 g,变异范围为19.66~58.82 g,变异系数为27.90%。千粒重的多样性指数最高,单株穗数次之。
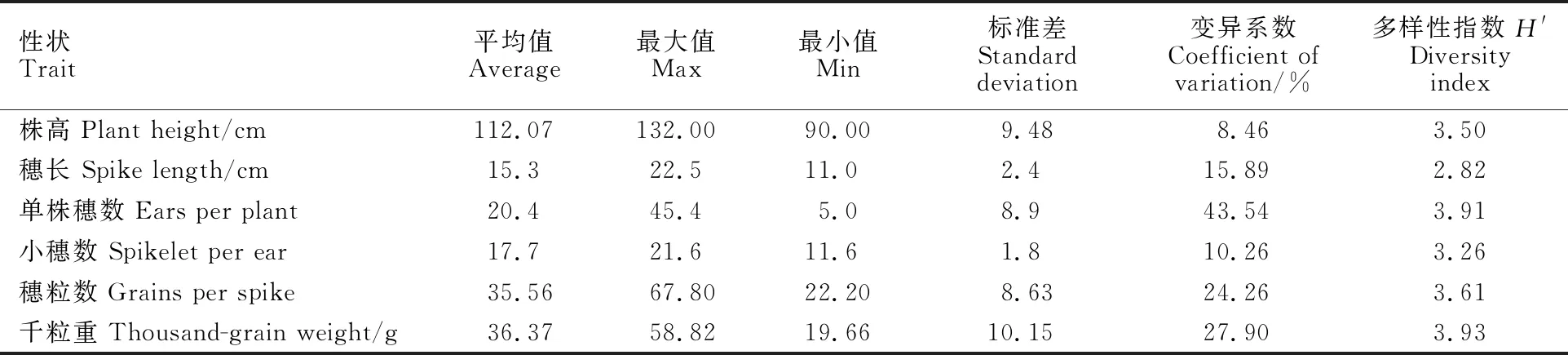
表2 供试材料主要农艺性状的变异特点
大部分材料的籽粒颜色为红色,籽粒形状呈长圆形,壳色多为白色,且颖壳多为无毛性。
调查中发现,供试材料具有繁茂性好、植株较高等特点。多数材料具有极强的分蘖成穗能力,如Syn1、Syn2、Syn56等材料的单株成穗数在10穗以上,Syn46的单株穗数平均达到达到45.4个;有的材料如Syn19、Syn30等穗粒数较多,平均达到50粒以上;Syn52、Syn50、Syn54等材料的千粒重较高,达50~60 g。
2.2 供试材料的白粉病抗性
以当地感病小麦品种济宁12为对照,在田间自然发病条件下,连续三年对供试材料白粉病抗性鉴定。结果表明,56份供试材料中Syn4、Syn23、Syn24、Syn32、Syn33、Syn40、Syn43、Syn54、Syn55共9份材料在田间表现为免疫或近免疫,占所有供试材料的16.1%;Syn8、Syn12、Syn15、Syn16、Syn17、Syn18、Syn22、Syn25、Syn26、Syn27、Syn29、Syn31、Syn35、Syn36、Syn39、Syn41、Syn42、Syn45、Syn47、Syn50、Syn51、Syn52、Syn53共23份材料表现高抗白粉病,占所有供试材料的41.1%;Syn1、Syn2、Syn3、Syn9、Syn13、Syn19、Syn20、Syn34、Syn44、Syn46、Syn48、Syn49、Syn56共13份材料表现中抗白粉病,占供材料的23.2%;有19.6%的材料表现中感小麦白粉病,主要是Syn5、Syn6、Syn7、Syn10、Syn11、Syn14、Syn21、Syn28、Syn30、Syn37、Syn38等。综上,供试材料中有77%以上的材料对白粉病具有良好的抗性。
2.3 供试材料的高分子量谷蛋白亚基组成
从图1、表3和表4可以看出,供试份材料在编码高分子量麦谷蛋白亚基的Glu-A1、Glu-B1和Glu-D1这3个基因位点上表现出丰富的多态性,共有35种HMW-GS等位变异。其中,Glu-A1、 Glu-B1、 Glu-D1位点上分别有3、15、17种等位变异类型。从不同亚基的出现频率看,在Glu-A1位点上的3种变异类型(1,2*,null)中,null亚基频率最高,为51.8%,1亚基次之 (44.6%),2*亚基最少(3.6%)。Glu-B1位点上的亚基类型丰富,检测到15种亚基类型,其中7+8亚基出现频率最高,为25%;其次为17+18,为 12.5%;再次为13+16,为10.7%;还检测到6、7+8+9、8+20等稀有亚基类型。在Glu-D1位点的17种亚基类型中,2+12亚基最多,频率为30.4%;其次是优质亚基5+10,占12.5%;同时还检测出5+12亚基类型,占5.4%。

表3 供试材料的高分子量谷蛋白亚基组成
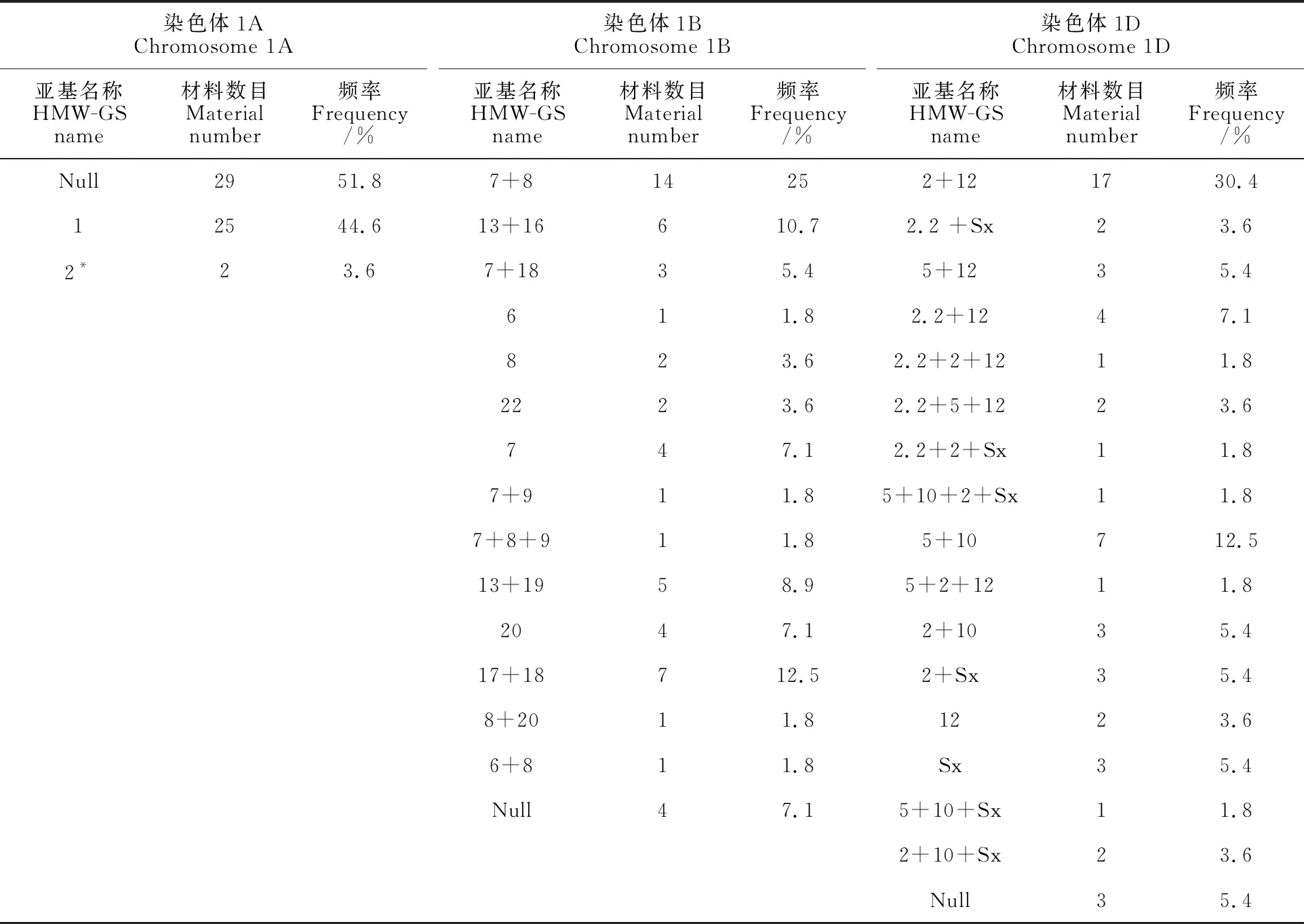
表4 供试材料的HMW-GS在不同位点的变异类型及其频率

1:Syn39; 2:Syn40; 3:Syn41; 4:Syn42; 5:Syn43; 6:Syn44; 7:Syn45; 8:Syn46; 9:Syn47; 10:Syn48; 11:Syn49;12:Syn50; 13:Syn51; 14:Syn52; 15:Syn53; 16:Syn54; 17:Syn55; 18:烟优361 Yanyou 361; 19:马奎斯 Makuis; 20:中国春 Chinese Spring.
此外,还在10份材料中检测到一种新的亚基,暂命名为Sx。该亚基的分子量比目前已知分子量最小的12亚基还小,而且单独表达或与2、5、12等亚基呈双亚基、三亚基、四亚基组合的形式出现,如5+10+Sx、2+10+Sx、2+2+Sx和 5+10+2+Sx等。
上述结果表明,供试材料Glu-1位点上的亚基变异类型非常丰富,除含有多种优质亚基或亚基组合外,还含有新的亚基类型,因此在小麦加工品质性状的遗传改良中具有重要的应用价值。
2.4 遗传多样性
选择田间农艺性状较好的材料Syn2、Syn44和Syn54(图2)与5个当地栽培品种济宁12、济麦22、良星99、临麦4号、郑麦7698,进行了全基因组遗传多态性分析。用筛选出的20对条带清晰、多态性较好的引物(表5)进行扩增分析,结果(图2)表明,Xgwm312在供试材料中可扩增出3种等位变异,在栽培品种中可以扩增出2种等位变异。20对引物在供试材料中共检测到94个等位变异。平均每对引物在3个人工合成小麦中能检测到2.9个等位变异,PIC值为0.65;在5个栽培品种中检测到1.8个等位变异,PIC值为 0.28。说明人工合成小麦的遗传多样性高于栽培品种。

A:Syn54灌浆期; B:Syn54成熟期;C和D中:1.济宁12,2.Syn2,3.Syn44,4.Syn54,5.良星99,6.临麦4号,7.济麦22,8.郑麦7698。
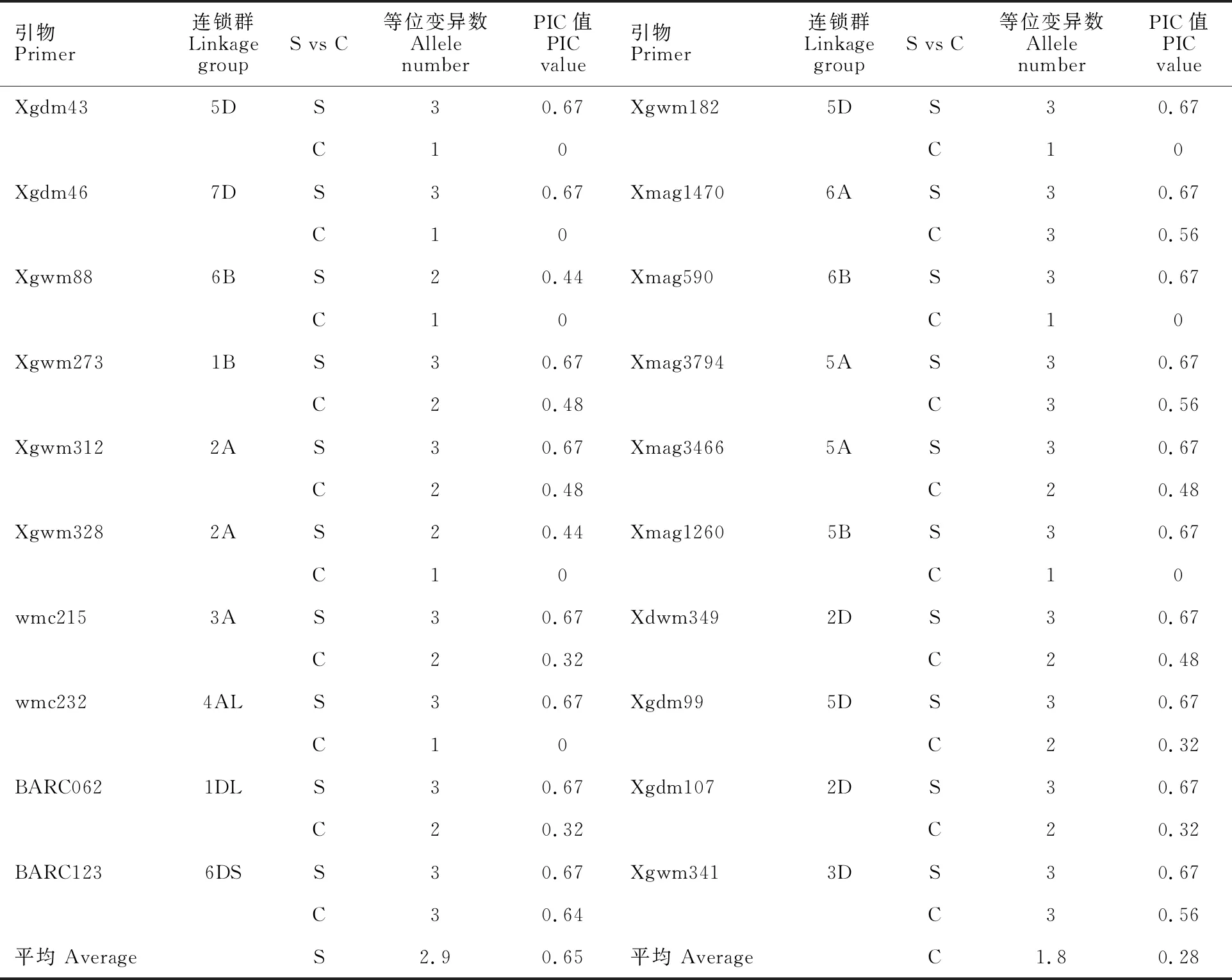
表5 SSR引物在部分供试材料中的扩增结果
3 讨 论
由于长期集中利用少数骨干亲本材料,导致现代育成小麦品种的遗传相似性增加,遗传多样性减少,降低了小麦品种的适应性,产量水平也难有新的突破。如何拓宽小麦的遗传基础是当前广大小麦育种家急待解决的重要课题。
利用四倍体小麦和粗山羊草杂交获得的六倍体人工合成小麦,具有丰富的遗传多样性,含有许多在小麦育种中具有利用价值的优异基因,它已作为拓宽普通栽培小麦品种遗传多样性的新种质资源,日益受到育种家的重视。叶亚青[23]对9个人工合成小麦的S1代形态学研究表明,栽培二粒小麦与粗山羊草的人工合成小麦遗传了粗山羊草的高分蘖特征,但是植株的高度则具有栽培二粒小麦的特点。陈国跃[24]分别以97份引自国际玉米小麦改良中心(CIMMYT)人工合成六倍体小麦和中国农科院品资所的16份人工合成六倍体小麦为材料,对其进行了抗条锈病和白粉病鉴定:证明有48份材料对条锈病生理小种条中31、条中32和水源14表现抗性,54份材料对小麦白粉病菌15号生理小种具有抗性,其中16份材料表现为免疫或近免疫。本研究对引自CYMMIT的56份人工合成小麦材料进行了农艺性状鉴定,结果表明,供试材料的性状变异主要集中在千粒重和单株穗数等产量性状上,整体穗粒数较多,分蘖成穗多。单株穗数在10个以上的占到全部材料的91%,高于平均值20.44个的有25份,Syn46的单株穗数达到45.4个。这表明六倍体人工合成小麦具有较高的分蘖成穗潜能,在小麦单株穗数的遗传改良中具有较大的应用价值。在抗白粉病方面,Syn4、Syn23、Syn24等9份材料在田间表现免疫或近免疫,占所有供试材料的16.1%,Syn8、Syn12、Syn15等23份材料表现高抗白粉病,占41.1%,Syn1、Syn2、Syn3等13份材料表现中抗白粉病,占比23.2%。说明多数供试材料对白粉病具有良好的抗性,在小麦抗白粉病育种方面具有利用价值。
对供试材料的HMW-GS组成进行分析,共检测到35种HMW-GS等位变异。其中,Glu-A1位点上有3 种等位变异类型;Glu-B1 位点有15种等位变异类型,Glu-D1 位点检测到17种等位变异类型。鉴定出优质亚基17+18、13+16、2*、5+10等亚基和重组的亚基类型:5+12。同时还发现了一些新的未知亚基或亚基组合。这与陈国跃[24]对人工合成六倍体小麦高分子量麦谷蛋白亚基研究结果相似,但发现的亚基变异类型更多,并出现了四亚基组合类型5+10+2+Sx等,表明供试材料在小麦加工品质性状的遗传改良中可能具有重要利用价值。其中新亚基对小麦加工品质性状的影响有待进一步研究。
微卫星DNA标记具有操作方便、成本低廉等特点,已经被大量地应用到包括小麦族在内的许多物种的遗传多样性评价研究中[25-27]。Lelley等[28]利用SSR标记技术,对粗山羊草和普通小麦的遗传多样性分析结果表明,在每个SSR位点上所检测到的等位变异都比在栽培小麦品种中的多[28]。本研究用20对SSR分子标记进行遗传多样性分析,结果显示,供试的人工合成小麦中扩增出更多的等位变异类型。因此,人工合成小麦具有较高的遗传多样性。
本研究结果为供试材料的后续研究和有效利用提供了参考,奠定了基础。
猜你喜欢
作物学报(2022年6期)2022-04-08 01:26:24
今日农业(2021年9期)2021-11-26 07:41:24
国际医学放射学杂志(2021年5期)2021-10-22 07:26:20
世界最新医学信息文摘(2020年68期)2020-12-25 11:55:27
麦类作物学报(2018年4期)2018-05-11 09:34:08
第一财经(2017年36期)2017-09-25 06:17:11
西藏科技(2016年8期)2016-09-26 09:00:21
吉林大学学报(医学版)(2015年4期)2015-12-17 07:48:13
现代农业(2015年5期)2015-02-28 18:40:49
新疆农垦科技(2014年1期)2014-02-28 19:19:01
