
高温天数和吡虫啉互作对麦长管蚜世代内与世代间生态表型的影响
2021-10-20 06:42:14曹俊宇孙东宝
环境昆虫学报 2021年5期
曹俊宇,孙东宝,赵 飞
(1.吕梁学院生命科学系,山西吕梁 033000;2.山西农业大学植物保护学院,农业有害生物综合治理山西省重点实验室,山西太原 030031;3.中国农业科学院农业环境与可持续发展研究所,北京 100081;4.山西寿阳旱地农田生态系统野外科学观测研究站,山西寿阳 031700)
麦长管蚜Sitobionavenae是重要的全球麦类害虫之一,生活周期短、繁殖能力强,爆发频率较高,给麦类作物造成了严重的产量、质量损失(Honěketal., 2020; 姜玉英等, 2020)。目前防治麦蚜的主要手段仍然是化学农药,吡虫啉作为近几年发展较快的新烟碱类杀虫剂,广泛用于防治刺吸式口器的害虫(Yuanetal., 2020; 丁建朋等, 2020)。实际生产中,由于农药利用率的限制,环境中喷施的农药往往以低剂量药效发挥作用,因此,研究低剂量农药的效应更有现实意义。考虑到麦长管蚜的防治是在自然环境温度不断变化的条件下进行的,所以麦长管蚜在发生期间可能会同时受到高温和农药交互所带来影响。
麦长管蚜体型小,身体的比表面积大,与外界环境的热传导和热交流较快,对外界温度变化非常敏感(Angilletta, 2009)。通常一天当中的最高温可以代表一天的整体温度趋势,对生物的影响也较大,尤其是随着气候变暖的加剧,高温天发生更加频繁(Huetal., 2020; Reyetal., 2020),有时甚至会持续数天,因此生活在自然环境中的生物必将受到重复数天的高温天的影响。迄今为止,高温对生物存活、寿命、繁殖(Caoetal., 2018; Zhaoetal., 2019; Maetal., 2020)等生活史性状的研究已受到大量关注,但与农药结合对生物生态表型的考察则较为有限。
已有高温与农药互作研究表明,两胁迫互作会产生直接的即时生态效应。例如,高温天暴露会增加毒死蜱的毒性,显著降低异痣蟌Ischnuraelegans(Verheyenetal., 2019)和尖音库蚊Culexpipiens的存活率(Delnatetal., 2019);高温和农药吡虫啉结合会降低农药对西方蜜蜂Apismellifera致死率(Saleemetal., 2020)。同时还可产生延迟生态效应,早期高温和农药的处理会延缓处女丝蟌Lestessponsa的生长速率,延长发育时间(Sniegulaetal., 2017)、影响褐飞虱Nilaparvatalugens成虫的繁殖性状(Yuetal., 2012; Chengetal., 2014)。但这些高温农药互作研究仅考虑对世代内性状的影响,对跨代的影响却鲜有研究,这将会限制我们对高温农药互作生物影响的全面评估。
因此,本试验研究了高温事件34℃/3 h持续1 d、3 d和5 d与吡虫啉互作对麦长管蚜成蚜进行处理后,对母代存活、寿命、繁殖以及子代生长发育、繁殖、种群参数等生活史性状的影响,明确高温天数和农药互作对母代以及子代的生态表型影响,尤其是子代对母代胁迫经历的复杂响应,从而提高高温农药互作对生物种群动态预测的综合评估能力。
1 材料与方法
1.1 供试昆虫
为保证非遗传世代间效应分析(Andersenetal., 2005),实验室建立了麦长管蚜单克隆品系,该克隆系于2016年5月采自临汾郊区的冬麦田(35o55N, 111o16E),经室内饲养2年后用于实验。采用10~20 cm高的小麦幼苗在养虫笼内(34 cm×32 cm×34 cm)饲养。每周更换1次新鲜小麦幼苗。饲养条件:养虫室温度22℃±0.5℃,相对湿度50%~70%,光周期L∶D=16 h∶8 h。
1.2 亚致死农药剂量确定
95%吡虫啉原药(中农联合生物科技有限公司提供)。为了确定施药浓度,根据Shi等(2011)的浸叶法,方法略有改动,对刚羽化的成蚜进行毒力测定。将吡虫啉原药用比例为1∶9的吐温80(国药集团化学试剂有限公司)和丙酮分析纯作为溶剂制备成10 mg/mL的原液,再用蒸馏水稀释成5个系列浓度(0.5, 1.5, 50, 100, 300 μg/mL),以溶剂蒸馏水作为空白对照,剪取约5 cm长的麦苗将其浸到各个浓度梯度中大约 5~7 s,在空气中晾干,放入一个由滤纸保湿的培养皿中(直径90 mm),10片毒苗/皿,接入30头成蚜,24 h后记录存活蚜虫的个数,每处理重复3次。数据分析选用SPSS软件中的机值分析法。获得毒力回归方程:y=-1.917x + 0.823,选取亚致死浓度LC20= 20 μg/mL为农药处理浓度(表1),试验时,毒苗的制备同样采用浸叶法。
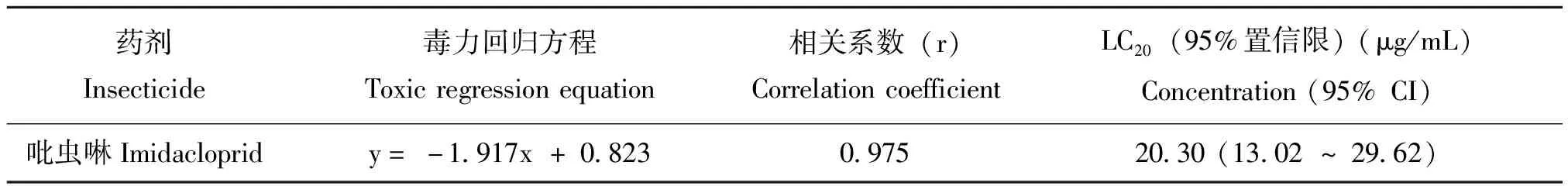
表1 吡虫啉对麦长管蚜无翅成蚜的毒力
1.3 高温天数和吡虫啉互作对麦长管蚜生态表型的影响
为了研究高温天数与农药吡虫啉互作对麦长管蚜世代内与世代间生活史性状的影响,其中高温事件34℃/3 h处理天数为1 d、3 d和5 d,农药吡虫啉(有无)处理时间为24 h相结合,共计6个处理。其中40头蚜虫/处理用于母代生活史性状调查,另80头蚜虫/处理作为备用仅用于子代的采集。
高温天数处理:将麦长管蚜成蚜单头接入插有新鲜无药麦苗的饲养管后,进行不同高温天数的处理,以24 h为周期,定点在34℃培养箱中进行3 h热处理,其余时间转移到22℃下继续饲养,处理天数为1 d、3 d、5 d。试验期间麦苗每3 d更换 1次。
高温+吡虫啉互作:将麦长管蚜成蚜单头接入插有浸毒麦苗的饲养管中,进行不同高温天数处理,且农药处理时间为24 h,结束后需将试虫转移至新鲜无药麦苗的饲养管中,接着完成余下高温天数的处理,直到处理全部结束。
世代内性状调查:试验期间,每天处理结束后,在第2天调查记录1次存活状态、产仔情况,并将计数后若蚜去除,直至供试蚜虫全部死亡,调查结束。存活率是各处理结束后,存活个体占处理总数的比例;成蚜寿命是从处理起始开始算起,直到死亡的时间;繁殖同样从处理起始开始记录,整个存活时期每头蚜虫所产后代的总数。
世代间性状调查:各处理结束后第二天调查时,随机采集30头子代/处理(共6个处理)新生若蚜置于常温22℃养虫室内,于每日上午8点记录死亡个体、蜕皮及产仔情况,并将蜕皮、死亡蚜虫及新生若蚜去除,直至供试蚜虫全部死亡。去除逃逸蚜虫,所有处理蚜虫均测定了以下指标:若虫死亡率、发育时间、成蚜寿命、繁殖。若虫死亡率是指活到成蚜的若虫占全部测试若虫的比例。发育时间是指新生若蚜发育至成蚜所需的时间。成蚜寿命是指从变成成蚜到蚜虫死亡的时间。成蚜繁殖是指每头成蚜所产后代的总数。
1.4 数据处理
对数据使用Shapiro-Wilks进行正态分布,发现试验中大部分数据不符合正态分布,因此高温天数间多重比较采用单因素Kruskal-Wallis进行分析,同一高温天数下施药和非施药差异显著性采用Mann-Whitney U检验检验。存活率显著性分析均采用列联表,非独立2×2以及2×3表法实现处理间的多重比较(Caoetal., 2018)。以上数据都运用SPSS version 20(SPSS Inc., Chicago, IL, USA)来进行分析。
种群参数(Zhuetal., 2018):净增长率R0=∑lx·mx,平均世代时间G=∑lx·mx·X/∑lx·mx,内禀增长率rm=ln(R0)/G,其中X为蚜虫的虫龄,lx为蚜虫在X时的成活率,mx代表蚜虫在X时所产的后代数量。在R语言中采用bootstrap程序计算每一种群参数的均值,并调用R语言中agricolae包中的sample函数和kruskal函数,采用Kruskal-Wallis法来比较各处理相应种群参数的差异显著性。
2 结果与分析
2.1 高温天数和吡虫啉互作对世代内生态表型的影响
2.1.1母代存活
不同高温天数处理(χ2=26.453, df=2,P<0.001)以及高温和吡虫啉互作(χ2=34.286, df=2,P<0.001)均对存活产生了显著影响,且随着高温天数的增加,存活率随之下降,仅高温处理下由67.5%下降为12.5%,与农药结合后由70%下降为10%(图1)。此外,吡虫啉的有无在高温持续1 d(χ2=0.058, df=1,P=0.809)、3 d(χ2=0.549, df=1,P=0.459)、5 d(χ2=0.125, df=1,P=0.723)后对存活均无显著影响(图1)。
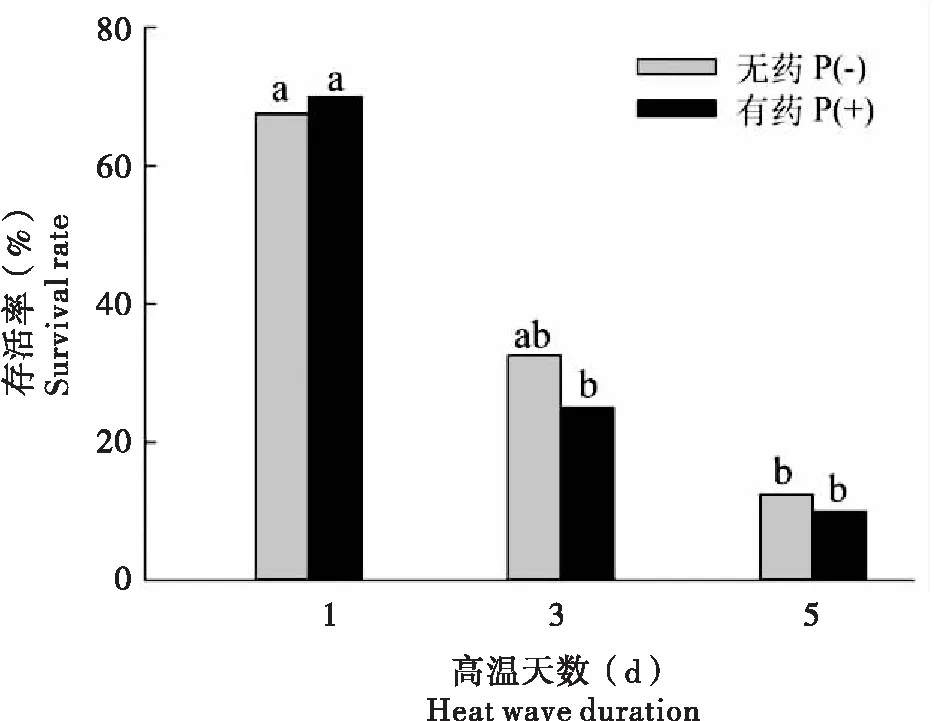
图1 高温天数和吡虫啉互作对母代麦长管蚜存活的影响
2.1.2母代寿命和繁殖
不同高温天数处理(χ2=1.739, df=2,P=0.419)以及高温和吡虫啉同时作用(χ2=0.352, df=2,P=0.838)均没有对母代寿命产生显著影响;且吡虫啉的存在,在高温持续1 d(U=750.000,P=0,623)、3 d(U=809.500,P=0.925)、5 d(U=709.000,P=0.367)后对寿命也均无显著影响(图2-A)。
与寿命相似,不同高温天数处理(χ2=1.125, df=2,P=0.570)以及高温和吡虫啉同时作用(χ2=2.411, df=2,P=0.300)均没有对母代繁殖产生显著影响;但吡虫啉的存在对繁殖却产生了负面影响,尤其是在高温持续3 d(U=595.500,P=0.032)和5 d(U=557.500,P=0.016)达到显著水平,分别降低了0.9±0.5头若蚜/成蚜和2.1±0.9头若蚜/成蚜(图2-B)。
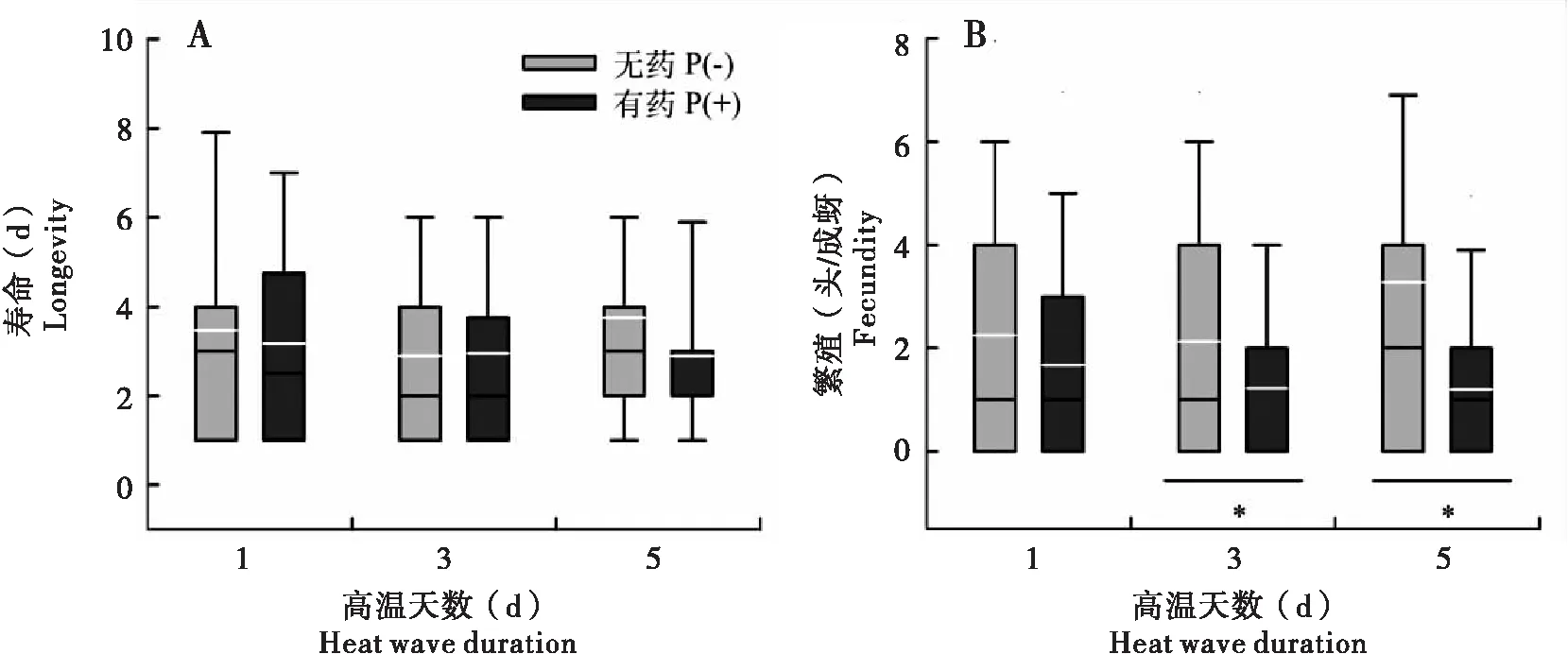
图2 高温天数和吡虫啉互作对母代麦长管蚜寿命和繁殖的影响
2.1.3母代种群参数
无论母代是否经农药处理,随着高温处理天数的增加,种群内禀增长率(rm)、净繁殖率(Ro)以及平均世代时间(G)都受到了抑制。且与单独高温处理相比,高温和吡虫啉的互作会对种群参数产生更加显著的负面影响(图3)。
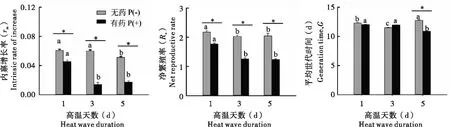
图3 高温天数和吡虫啉互作对母代麦长管蚜种群参数的影响
2.2 高温天数和吡虫啉互作对世代间生态表型的影响
2.2.1子代存活和发育时间
母代经历单独高温处理对子代若蚜存活率的影响差异不显著(χ2=0.111, df=2,P=0.946),但高温和农药同时作用(χ2=5.425, df=2,P=0.066)却对该性状产生临界显著影响;且与单独高温处理相比,吡虫啉的存在均会增加子代若蚜的死亡率,在高温持续1 d达到了显著临界水平(χ2=3.455, df=1,P=0.055),存活率下降了23.3%,在高温持续3 d后达到了显著水平(χ2=6.696, df=1,P=0.010),存活率下降了33.3%(图4-A)。
母代经历不同高温天数(χ2=2.055, df=2,P=0.358)以及高温和吡虫啉同时作用(χ2=0.213, df=2,P=0.899)均没有对子代发育时间产生影响;且母代是否经吡虫啉处理在高温持续1 d(U=198.500,P=0.304)、3 d(U=129.000,P=0.611)、5 d(U=232.000,P=0.745)对子代发育均无影响(图4-B)。
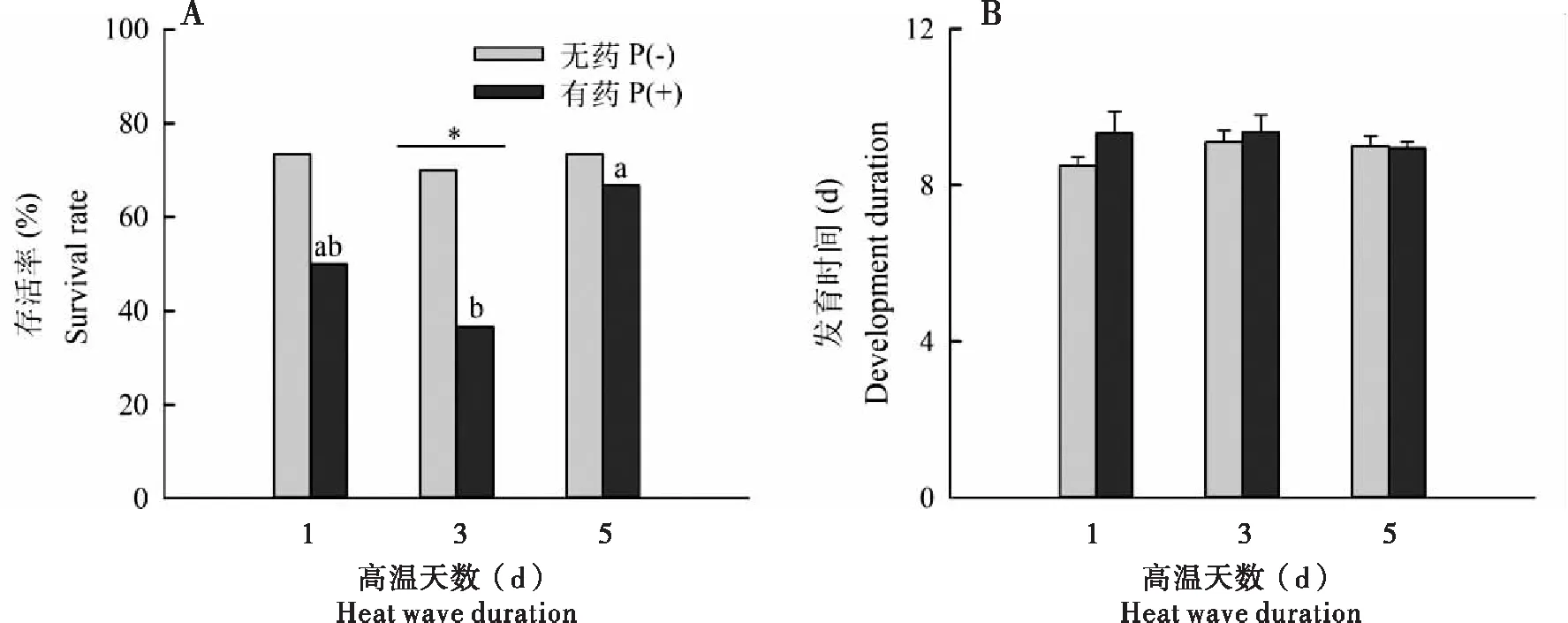
图4 母代经历高温天数和吡虫啉互作对子代麦长管蚜存活、发育的影响
2.2.2子代成蚜寿命和繁殖
母代经历不同高温天数处理(χ2=2.640, df=2,P=0.267)以及高温和吡虫啉互作(χ2=2.010, df=2,P=0.366)均对子代成蚜寿命的影响差异不显著;且与母代单独经历高温处理相比(1 d: 13.3±1.8 d; 3 d: 10.9±1.3 d; 5 d: 14.0±1.2 d),高温和吡虫啉的互作均会延长子代成蚜寿命(1 d: 17.2±0.7 d; 3 d: 15.7±1.6 d; 5 d: 15.3±1.1 d),尤其是在高温持续3 d(图5-A)。
与寿命相似,母代经历高温处理(χ2=2.976, df=2,P=0.226)以及高温和吡虫啉同时作用(χ2=4.079, df=2,P=0.130)均没有对子代繁殖产生显著影响;尽管与母代单独经历高温处理相比,母代经吡虫啉处理在高温持续1 d(U=195.000,P=0.366)、3 d(U=124.500,P=0.725)、5 d(U=366.500,P=0.421)对子代繁殖均无显著影响,但是均有轻微的刺激作用(图5-B)。

图5 母代经历高温天数和吡虫啉互作对子代麦长管蚜寿命和繁殖的影响
2.2.3子代种群参数
无论母代是否经农药处理,随着高温处理天数的增加,子代种群内禀增长率(rm)和净繁殖率(Ro)均是先下降后上升,平均世代时间(G)逐渐缩短。且与母代单独经历高温处理相比,吡虫啉的存在均会对子代种群参数产生不同程度的负面影响(图6)。

图6 母代经历高温天数和吡虫啉互作对子代麦长管蚜种群参数的影响
3 结论与讨论
在世代内,对存活和种群参数的调查发现,无论吡虫啉是否存在,随着高温天数的增加,存活率和种群增长率也相应减少。说明随着高温暴露时间的增加,会降低昆虫的存活,而减缓种群的增长速度,这与多数研究结果相同,如黑脉金斑蝶Danausplexippus不同龄期幼虫的存活率随着重复高温持续天数的增加而逐渐降低(Yorketal., 2002);麦无网长管蚜Metopolophiumdirhodum成蚜在高温持续天数增加的重复高温暴露下死亡率逐渐增加(Maetal., 2013);白纹伊蚊Aedesalbopictus成虫暴露在极端高温天越长,造成的死亡率越大,对种群产生的负面影响也越大(Jiaetal., 2019)。而对寿命和繁殖而言,无论吡虫啉是否存在,高温天数的增加对寿命、繁殖均无显著影响。该结果与Ma等(2004)发现麦无网长管蚜成蚜随着高温天数的增加,繁殖量和寿命同样随着减少的结果以及Hercus等(2003)研究表明,重复高温天数从1 d增加到4 d,黑腹果蝇Drosophilamelanogaster寿命的延长效果逐渐增强均不一致。这种现象可能是由于与高温的选择有关,高温34oC/3 h在持续1 d后产生的胁迫已使得麦长管蚜的寿命、繁殖降到了极限,随着高温天数的增加,已没有下降的空间,否则只能死亡,而随着高温天数的增加死亡率相应增加也充分说明了这一点。此外,相较单独高温处理,高温和吡虫啉的互作尽管没有对世代内存活、寿命产生显著影响,但却对繁殖产生较显著的负面影响,这说明高温天数和农药互作会提高胁迫的程度,恶化单独高温处理所产生的延迟效应。这与大多数研究报道高温和农药互作会产生协同影响相一致(Janssensetal., 2017 ; Zhenetal., 2019; Theysetal., 2020)。这种高温和农药毒性之间存在的正相关性的原因主要是高温增加了生物对于农药的摄入量(Holmstrupetal., 2010)。
在世代间,对子代性状而言,母代经历不同高温天数处理,对子代存活、发育、繁殖和寿命均没有显著影响,说明高温单独暴露对麦长管蚜生活史性状的影响并没有延续到子代。但是当高温和农药互作后,却对子代的存活产生显著负面影响,且随着高温处理天数的增加,对存活的负面影响表现先加重后减缓的趋势,这主要是由于随着高温处理天数的增加,对母代的损伤逐渐增大,造成母代死亡率逐渐增大,但考虑到高温持续1 d后造成的损伤较小,同时高温持续5 d后较强选择作用,致使存活下的母代个体适合度相差不大,但持续3 d就可能会存在适合度较差的母代个体,因此产生的后代质量差异也较大,相较其他处理对若蚜存活产生的负面影响较强。这在子代发育以及种群内禀增长率中也有不同程度的体现。此外,母代经历高温和吡虫啉两胁迫的互作对子代存活和发育均产生了不同程度的负面影响,这再次证明高温和农药互作加强了胁迫程度(Wangetal., 2007 ; Janssensetal., 2017)。但是这种负面影响并没有一直延续,甚至在子代成蚜的寿命、繁殖产生了正面刺激作用。这说明母代的胁迫经历,对子代的负面影响在前期的发育过程中通过补偿效应得到了恢复(Stoksetal., 2006; Dmitriewetal., 2007),并产生了一定毒物兴奋效应(Liangetal., 2014 ; Wangetal., 2016),刺激生长,因此相较母代种群增长速率的显著下降,子代种群增长速率却得到很大的缓解。
总之,麦长管蚜经历高温和吡虫啉的互作后显著抑制了母代种群的增长,但却对子代的负面影响仅限于发育、存活,甚至对子代成蚜性状产生了刺激作用,阐明世代内和世代间生物对于胁迫的复杂响应,表明仅仅根据一代的影响来预测胁迫效果尤其是双重胁迫对生物所产生的影响并不合理,暗示了跨代效应对于生物胁迫响应的重要性。同时结果也为气候变暖和农药双重胁迫下麦长管蚜种群动态的预测、防控提供一定的理论指导。
猜你喜欢
云南化工(2021年9期)2021-12-21 07:43:42
河北果树(2021年4期)2021-12-02 01:14:40
四川蚕业(2021年4期)2021-03-08 02:59:56
世界热带农业信息(2018年3期)2018-09-26 07:56:20
爱你(2018年24期)2018-08-16 01:20:42
爱你·阳光少年(2018年8期)2018-05-14 09:01:18
中国照明(2016年4期)2016-05-17 06:16:18
广西林业科学(2016年2期)2016-03-20 05:53:21
广西林业科学(2016年1期)2016-03-20 05:32:58
广西林业科学(2016年4期)2016-03-16 05:44:47
