
饲料中添加黄连素对花鲈生长及脂肪代谢的影响
2021-10-18 09:39鲁康乐张春晓
动物营养学报 2021年9期
魏 宇 鲁康乐 王 玲 宋 凯 张春晓
(集美大学水产学院,厦门市饲料检测与安全评价重点实验室,厦门 361061)
高脂饲料是一种脂肪含量相对较高的高能饲料,可发挥“节约蛋白质”效应[1],以缓解鱼粉等蛋白质源的短缺,还可以减少养殖鱼类的氨氮排放,起到降低饲料成本和维持养殖环境的重要作用。特别是随着高密度集约化水产养殖业的发展,高脂饲料被广泛使用。然而高脂饲料的长期使用会加剧肝脏等部位的脂肪沉积,是诱发养殖鱼类脂肪肝的首要因素[2]。鱼类脂肪肝早期症状隐匿,随之发展会导致生长与饲料利用率下降,一旦遭受病原感染和环境应激极易发生群体死亡,给养殖业带来巨大的经济损失。因此,筛选适当的饲料添加剂对高脂饲料导致鱼类脂肪沉积进行调控非常有必要。
黄连素又称盐酸小檗碱,是从黄连、黄柏等植物中提取出来的生物碱,具有多种重要的生理功能,可有效调节脂肪代谢、改善脂肪肝症状和氧化应激等[3]。陈青青[4]研究发现,团头鲂高脂饲料中添加黄连素可以改善其免疫功能和抑制肝细胞凋亡,从而缓解高脂饲料导致的脂肪肝发生。黄连素来源广、成本低、副作用小、使用安全、易被吸收,是一种非常有潜力的用于调控脂肪沉积的物质,但其在海水鱼上的作用却未见报道。
花鲈又称海鲈、七星鲈,其肉质鲜美,具有较高的营养价值,是一种养殖量较高的海水鱼类,根据《中国渔业统计年鉴》的统计数据显示,2019年我国花鲈养殖产量已达18.02万t[5]。然而,花鲈的腹腔体积较大,使用配合饲料养殖的花鲈常导致脂肪在肝脏与腹腔大量沉积,易诱发脂肪肝,严重危害花鲈养殖的健康发展。据文献报道,花鲈对饲料脂肪的需求量为8%~10%[6];然而,实际生产中为了追求高产量与快速生长,花鲈饲料的脂肪含量普遍高达12%~14%。因此,本试验以花鲈为研究对象,研究了在高脂饲料下添加黄连素对花鲈生长、脂肪沉积与脂肪代谢的影响,旨在探讨黄连素调控脂肪沉积的作用。
1 材料与方法
1.1 试验饲料
本试验选用鱼粉、豆粕、谷朊粉为主要蛋白质源,鱼油、豆油、卵磷脂为脂肪源,配制4种饲料,分别为低脂饲料(LFD组)、低脂饲料+100 mg/kg黄连素(LFD+BBR组)、高脂饲料(HFD组)和高脂饲料+100 mg/kg黄连素(HFD+BBR组),试验饲料组成及营养水平见表1。
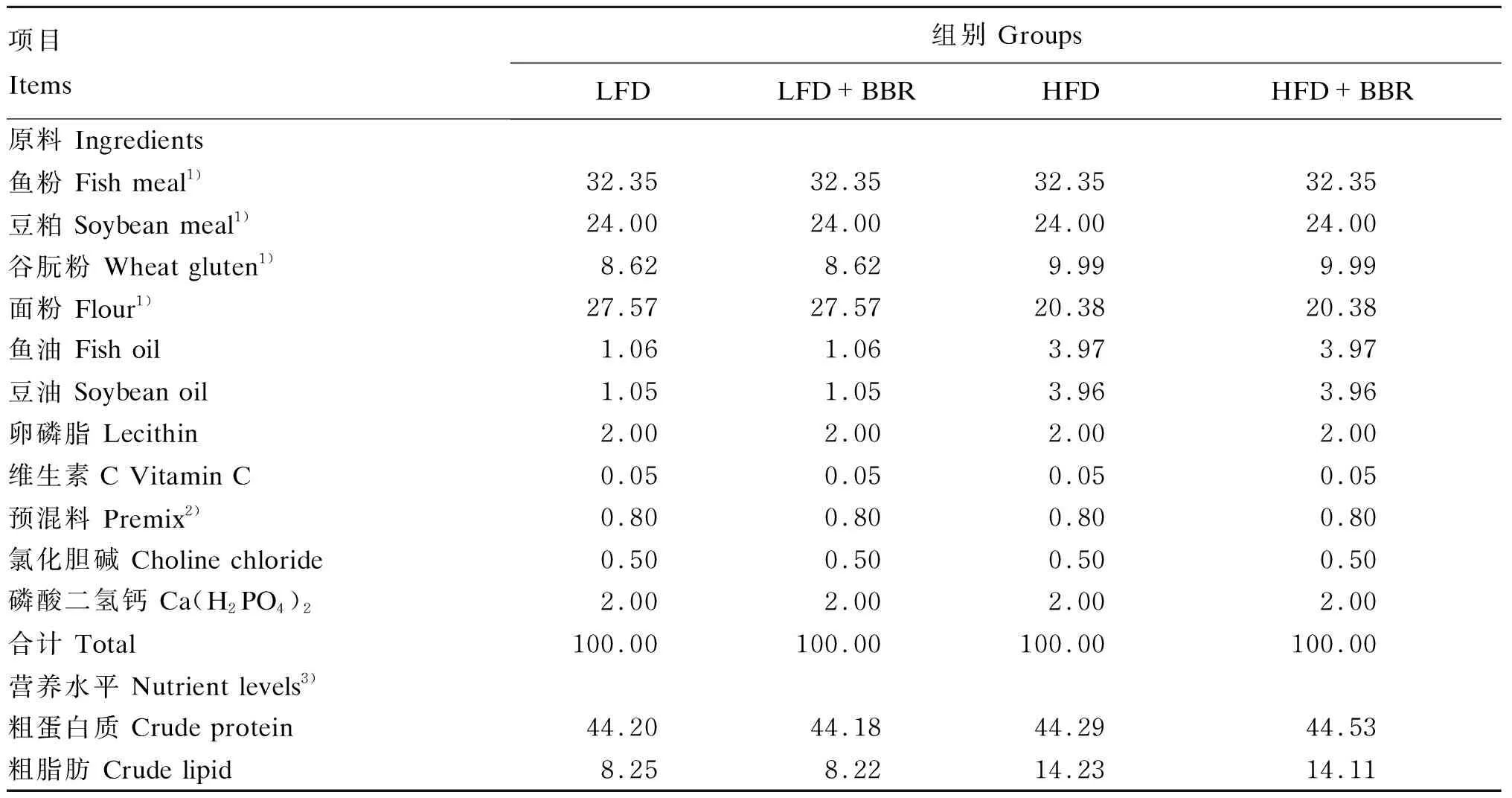
表1 试验饲料组成及营养水平(干物质基础)Table 1 Composition and nutrient levels of experimental diets (DM basis) %
3)营养水平均为实测值。Nutrient levels were measured values.
将各种原料粉粹后,按照表1称取各原料后,逐级、充分混匀,加适量的水进行制粒,制作了粒径分别为1.5和2.5 mm的2种颗粒饲料,将制粒后的饲料用烘箱烘干后,用自封袋保存于-20 ℃冰箱中待用。
1.2 试验鱼与饲养管理
试验所用花鲈鱼苗购于漳州诏安一家花鲈育苗场,养殖试验在集美大学水产学院龙舟池养殖实验场的室内循环系统中进行。将花鲈鱼苗运至实验场后,先暂养于养殖池中1个月,在此期间投喂商品饲料。待鱼苗适应试验环境后,开始分组进行养殖试验。
挑选360尾体质健壮、规格均一的花鲈[初始体重为(2.18±0.01) g],随机分到12个循环水养殖缸中,每缸30尾,用4种饲料投喂花鲈,每种饲料投喂3缸。养殖试验周期为8周,养殖试验前2周投喂1.5 mm粒径的饲料,养殖试验后6周投喂2.5 mm粒径的饲料。每天饱食投喂2次(08:30和17:30)。记录每缸鱼的摄食量,养殖试验期间保持以下条件:水温27~32 ℃,溶氧含量≥5 mg/L,盐度25.0~28.0 g/L,氨氮含量<0.1 mg/L,pH为7.5~8.5。
1.3 样品采集与分析
养殖试验结束后,将鱼饥饿24 h后用丁香酚麻醉将鱼麻醉(1∶10 000),捞出称量并计数。每缸随机取6尾对其进行尾静脉采血,离心分离血清后-80 ℃保存,用于检测血清生化指标;将鱼体置于冰上,用剪刀剪开鱼体腹部,随即取出内脏团,用镊子将肝脏和腹脂剥离,分别获得整个肝脏组织和腹腔脂肪组织(腹脂),并称量腹脂的重量以计算腹脂率,随后将肝脏和腹脂液氮速冻后于-80 ℃冰箱保存。将每缸剩余的鱼体称重后保存在-20 ℃冰箱中,用作分析鱼体全体组成。
血清总胆固醇(TC)和甘油三酯(TG)含量采用南京建成生物工程研究所的试剂盒,按照试剂盒说明书的操作步骤进行测定。
肝脏和腹脂的基因表达按照以下步骤进行:
1)总RNA提取与反转录。使用TRIzol试剂(TaKaRa)提取-80 ℃冻存的肝脏和腹脂组织总RNA,用NanoDrop ND-2000分光光度计测定提取的RNA浓度,并通过琼脂糖凝胶电泳确认RNA的完整性。然后使用Prime ScriptRT试剂盒(TaKaRa)将总RNA反转录为cDNA,-20 ℃冰箱保存待用。
2)实时荧光定量PCR。用Premier 5.0软件进行引物序列设计,引物合成工作由金唯智生物科技有限公司完成,引物序列如表2所示。加样上机,使用ChamQ Universal SYBR qPCR Master Mix试剂盒(南京诺唯赞)和QuantStudioTM6 Flex荧光定量PCR仪(ABI)进行实时荧光定量PCR,荧光定量PCR扩增体系为20 μL,包括10 μL ChamQ Universal SYBR qPCR Master Mix(2×)、0.4 μL PCR Forward Primer (10 μmol/L)、0.4 μL PCR Reverse Primer(10 μmol/L)、2.0 μL cDNA 模板、7.2 μL ddH2O。反应程序为95 ℃ 30 s;95 ℃ 5 s,60 ℃ 30 s,40个循环;95 ℃ 15 s,60 ℃ 62 s,95 ℃ 15 s。

表2 实时荧光定量PCR引物序列Table 2 Primer sequences used for RT-qPCR

续表2基因名称Gene names引物序列Primer sequence(5'—3')退火温度Tm/℃乙酰辅酶A氧化酶AOXF:ATACGTGGTCTGGGAGCCTAR:AGCTGGCTTGGCCTACATTC60脂肪甘油三酯脂肪酶ATGLF:CTTCCTCTCCGCAACAAGTCR:TGGTGCTGTCTGGAGTGTTC60线粒体E3泛素连接酶1Mul1F:GCTGCCGTGATACGAGTCATR:ACGTTGGACAAGGACTGGAC60自噬相关基因5Atg5F:TCAGTCGCTGCCATTAGAGCR:TCTCGTCACCTGCGAAAACT60PTEN诱导激酶1PINK1F:CTGTGAAAGCCCGGTACACTR:TGATGTGGAACTTTGGGGCA60β-肌动蛋白β-actinF:CGCCGCACTGGTTGTTR:TTTGGGGTTCAGGGGG60
1.4 指标计算
增重率(WGR,%)=100×(Wt-W0)/W0;
特定生长率(SGR, %/d)=100×(lnWt-lnW0)/t;
饲料系数(FCR)=Wf/(Wt-W0);
蛋白质效率(PER)=(Wt-W0)/Wp;
腹脂率(%)=100×Wa/W。
式中:W0为初始鱼总重量(g);Wt为终末鱼总重量(g);W为单尾鱼重量(g);Wf为摄入的饲料总量(干物质基础,g);Wp为摄入饲料蛋白质总量(g);Wa为腹脂重(g);t为饲喂天数(d)。
1.5 数据处理
用SPSS 22.0分析软件进行双因素方差分析(two-way ANOVA),采用Tukey检验法进行多重比较,差异水平定为P<0.05。试验数据以平均值±标准差(mean±SD)来表示。
2 结果与分析
2.1 黄连素对花鲈生长及饲料利用的影响
黄连素对花鲈生长及饲料利用的影响如表3所示,脂肪水平和黄连素对花鲈的生长性能有显著影响(P<0.05),但无显著交互作用(P>0.05)。HFD组鱼体的末体重、增重率、特定生长率和饲料系数显著高于LFD组(P<0.05);在LFD或HFD组饲料中添加黄连素均显著降低了鱼体的末体重、增重率、特定生长率、饲料系数和蛋白质效率(P<0.05),说明黄连素对鱼体生长有一定的副作用。此外,各组间摄食量、肝体比和存活率均无显著差异(P>0.05)。
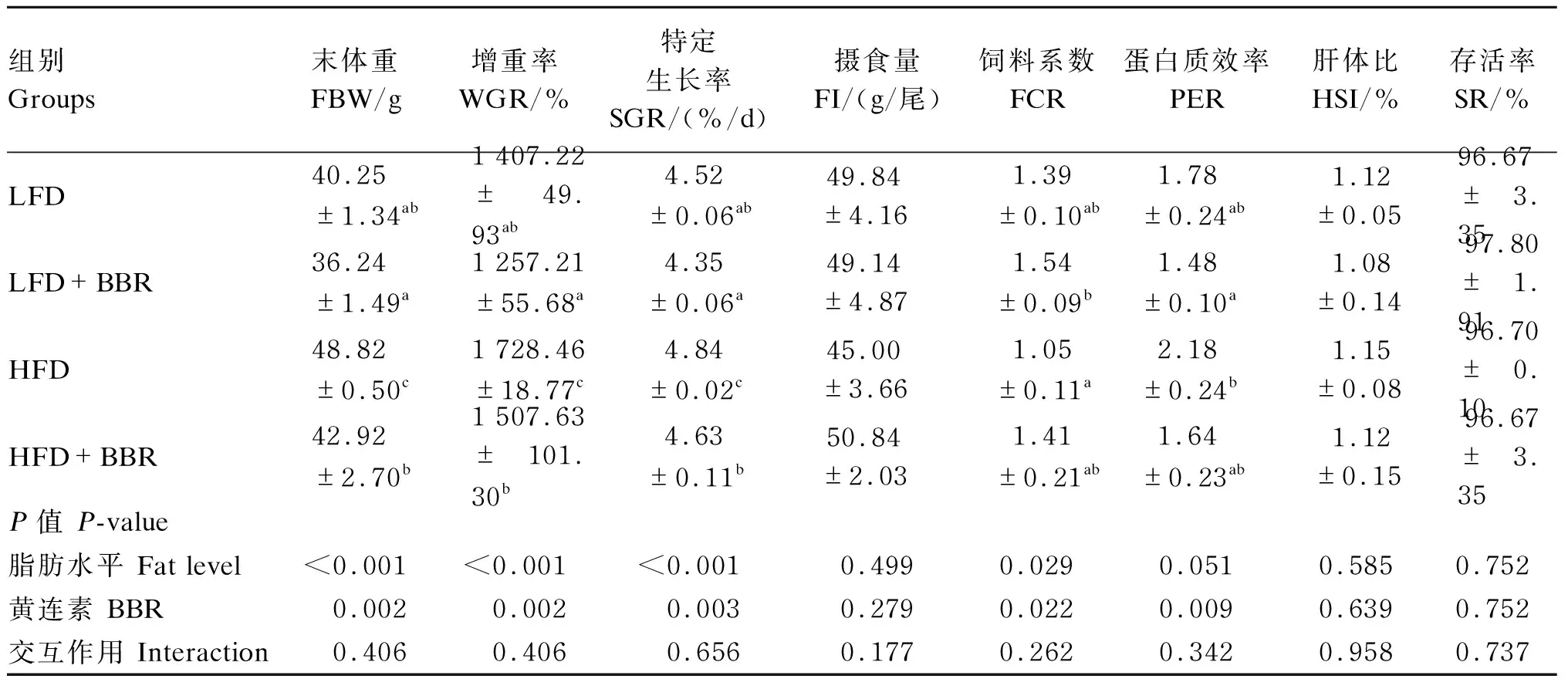
表3 黄连素对花鲈生长及饲料利用的影响Table 3 Effects of berberine on growth and feed utilization of L. maculatus
2.2 黄连素对花鲈脂肪沉积的影响
黄连素对花鲈脂肪沉积的影响如表4所示,与LFD组相比,HFD组花鲈的腹脂率、血清TG和TC的含量显著升高(P<0.05)。然而,饲料中添加黄连素显著降低了花鲈的腹脂率(P<0.05)。
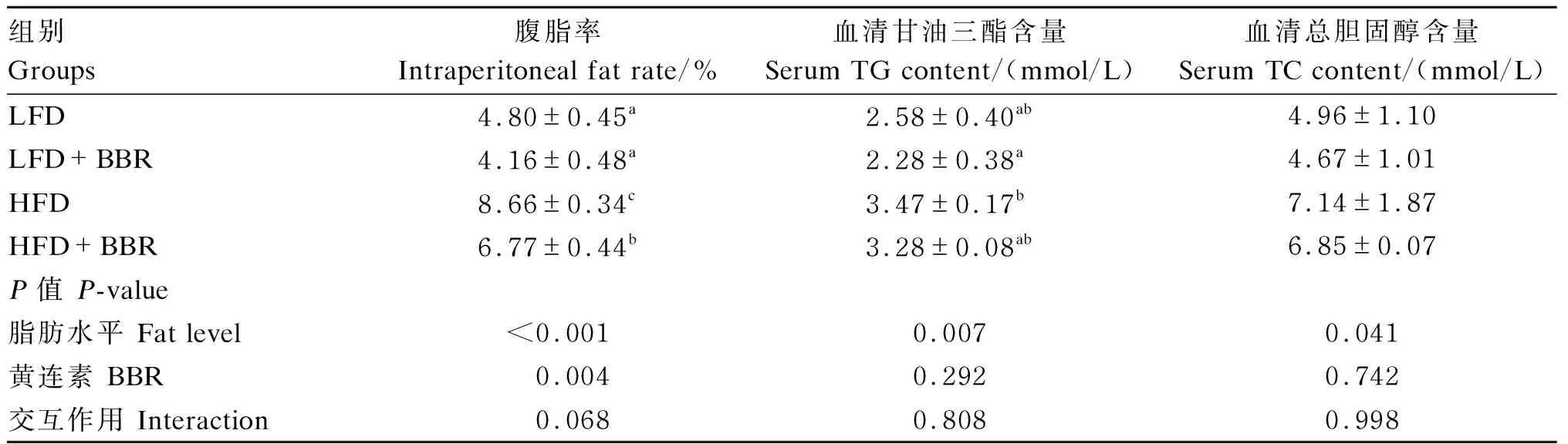
表4 黄连素对花鲈脂肪沉积的影响Table 4 Effects of berberine on fat deposition of L. maculatus
2.3 黄连素对花鲈肝脏基因表达的影响
黄连素对花鲈肝脏脂肪分解相关基因表达的影响如图1所示。肉碱棕榈酰转移酶 Ⅰ(CPTⅠ)、乙酰辅酶 A 氧化酶(AOX)、过氧化物酶体增殖受体 α(PPARα)和脂肪甘油三酯脂肪酶(ATGL)均是与脂肪分解相关的基因。与LFD组相比,HFD组AOX和PPARα基因相对表达量显著下调(P<0.05)、CPTⅠ基因相对表达量有下调的趋势但差异不显著(P>0.05)。与HFD组相比,饲料中添加黄连素后显著上调了CPTⅠ、AOX、PPARα和ATGL基因相对表达量(P<0.05)。
黄连素对花鲈肝细胞凋亡基因表达的影响如图2所示。与LFD组相比,HFD组的半胱氨酸天冬氨酸蛋白酶3(Caspase3)、半胱氨酸天冬氨酸蛋白酶9(Caspase9)和Bcl2 关联 X 蛋白(Bax)的基因相对表达量显著上调(P<0.05)。与HFD组相比,饲料中添加黄连素后显著下调了Caspase3和Bax基因相对表达量(P<0.05)。
2.4 黄连素对花鲈脂肪组织基因表达的影响
黄连素对花鲈脂肪组织凋亡基因表达的影响如图3所示。与LFD组相比,HFD组Bax的基因相对表达量显著下调(P<0.05)。饲料中添加黄连素后显著上调了Caspase9和Bax的基因相对表达量(P<0.05)。
黄连素对花鲈脂肪组织自噬基因表达的影响如图4所示。与LFD组相比,HFD组的PTEN 诱导激酶1(PINK1)、自噬相关基因5(Atg5)和线粒体E3 泛素连接酶1(Mul1)的基因相对表达量显著下调(P<0.05),饲料中添加黄连素后则上调了PINK1和Atg5的基因相对表达量(P<0.05)。
3 讨 论
脂肪是鱼类重要营养素,饲料中脂肪不足会影响鱼体的生长,还会导致必需脂肪酸缺乏症[7-8]。适当的提高饲料中脂肪的含量有利于促进鱼体的生长,本试验结果也表明,将饲料中的脂肪含量提高到14%显著提高了花鲈的增重率。然而过高的脂肪水平也会对鱼体产生一些不利的影响,会出现脂肪过度沉积等现象[9-11]。Wang等[12]研究发现,高脂饲料会导致军曹鱼腹腔和肌肉脂肪沉积。凌仕诚[13]研究发现,过量的脂肪会导致黄颡鱼肠系膜脂肪沉积。本试验结果也证实,HFD组花鲈腹部脂肪沉积提高。
研究表明,黄连素可以降低机体脂肪沉积[14]。为了缓解高脂饲料导致的鱼体脂肪过度沉积,本试验探究饲料中添加黄连素对花鲈生长及脂肪沉积的影响。由生长性能的数据看出,相较于HFD组,添加黄连素显著降低了花鲈的增重率。各组的摄食量变化不大,因此,黄连素降低花鲈增重率的原因并非是通过影响摄食,可能是通过促进分解代谢、增加能量消耗而导致的。
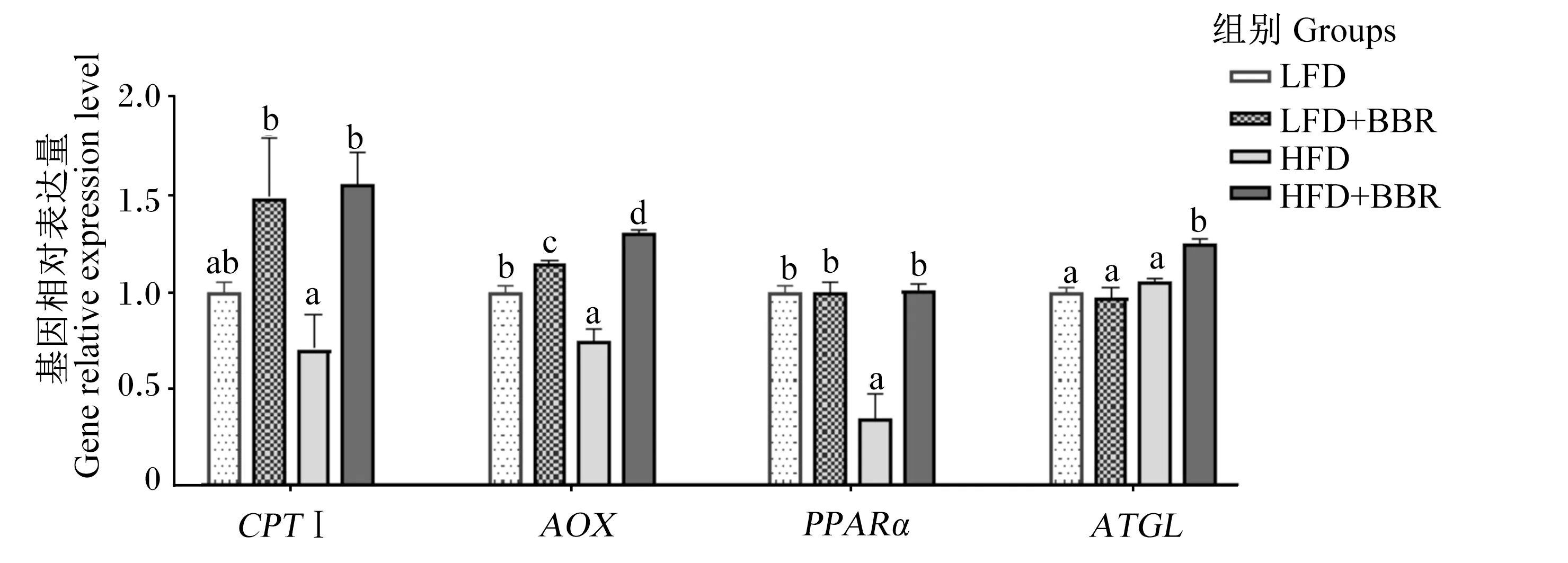
CPTⅠ:肉碱棕榈酰转移酶Ⅰ carnitine palmitoyl transferase Ⅰ;AOX:乙酰辅酶A氧化酶 acetyl-coa oxidase;PPARα:过氧化物酶体增殖受体α peroxisome proliferators activated receptor alpha;ATGL:脂肪甘油三酯脂肪酶 adipose triglyceride lipase。
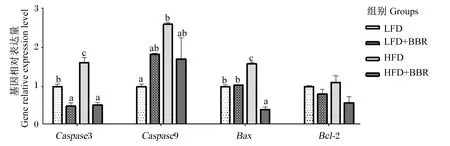
Caspase3:半胱氨酸天冬氨酸蛋白酶3 cysteinyl aspartate specific proteinase 3;Caspase9:半胱氨酸天冬氨酸蛋白酶9 cysteinyl aspartate specific proteinase 9;Bax:Bcl2关联X蛋白 Bcl2-associated X;Bcl-2:B细胞淋巴瘤-2 B-cell lymphoma-2。下图同 the same as below。
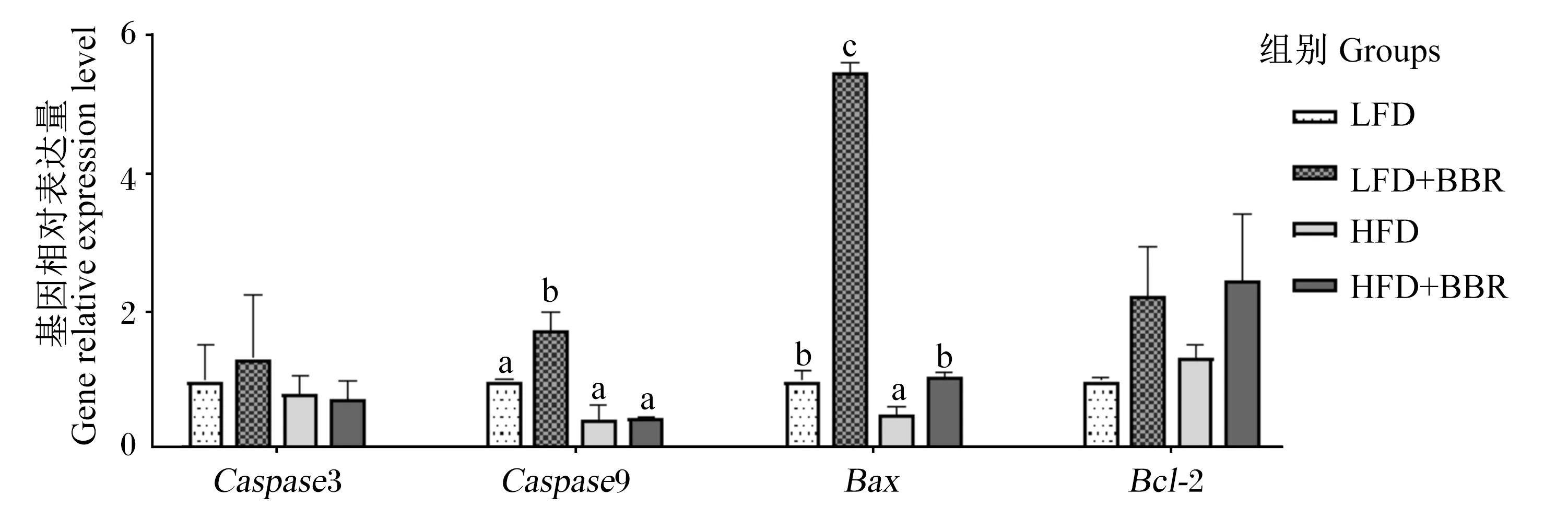
图3 腹脂中凋亡相关基因的相对表达Fig.3 Relative expression of apoptosis related genes in abdominal fat
陈青青[4]在团头鲂上的研究表明黄连素具有促进脂肪分解代谢的作用,与本试验所得到的高脂饲料中添加黄连素后花鲈脂肪沉积显著降低结果相似。因此,本研究探讨了黄连素对花鲈肝脏脂肪代谢的影响。结果表明,高脂饲料会下调AOX和PPARa的基因相对表达量,而添加黄连素之后显著上调了CPTⅠ、AOX、PPARa和ATGL的基因相对表达量。CPTⅠ是脂肪酸β-氧化过程中的限速酶,对脂肪代谢具有重要的调控作用,其表达水平升高有助于脂肪分解代谢,降低脂肪沉积[15-16]。过氧化物酶体增殖物激活受体家族成员PPARα是CPTⅠ的上游调控因子[17],PPARα可通过调控CPTⅠ来调节脂质代谢[18]。这与在团头鲂[19]中的研究结果一致,说明黄连素可能通过上调PPARα基因相对表达量,从而调节CPTⅠ的表达加快脂肪氧化分解,最终缓解了脂肪沉积。AOX是过氧化物酶体β-氧化的限速酶,有研究表明,AOX活性降低导致过氧化物酶体β-氧化速率下降[20];ATGL是甘油三酯水解的限速酶,是催化甘油三酯第1步水解的重要酶,在脂肪分解中发挥着重要的作用[21]。在本试验中,黄连素的添加可能是通过提高CPTⅠ、PPARα、AOX的活性,从而提高脂肪酸的氧化速率,并且促进了TG的降解,从而缓解脂肪的沉积。这也提示了黄连素可能会增加机体对能量的消耗,而使得体重降低。
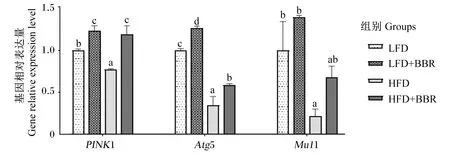
PINK1:PTEN诱导激酶1 PTEN induced putative kinase 1;Atg5:自噬相关基因5 autophagy related gene 5; Mul1:线粒体E3泛素连接酶1 mitochondrial E3 ubiquitin ligase 1。
肝脏脂肪的过度沉积往往会损伤肝细胞,常导致肝细胞凋亡[22-23]。凋亡主要包括外源性的死亡受体途径和内源性的线粒体途径,Caspase等蛋白水解酶是细胞凋亡过程中的重要效应因子,Caspase可在线粒体途径中被激活而诱发凋亡,因此Caspase被称之为促凋亡因子[24]。本试验结果可知,高脂饲料组花鲈肝脏Caspase3、Caspase9和Bax的基因相对表达量上调,说明HFD组肝细胞的凋亡被激活,肝细胞凋亡增多。而添加黄连素后,花鲈肝脏Caspase3和Bax的基因相对表达量显著下调,说明黄连素缓解了高脂饲料导致的花鲈肝细胞凋亡,起到了保护肝细胞的作用。
由本研究可知,黄连素有效降低了花鲈腹脂的含量。因此,本试验还探讨了黄连素降低腹脂的分子机制。结果表明,高脂饲料可以显著降低脂肪细胞的凋亡和自噬相关基因的表达。据有关文献可知,脂肪细胞凋亡后导致脂肪细胞的总数减少了,从而导致脂肪沉积的减少[25-26]。此外,高脂饲料可以显著降低脂肪细胞自噬相关基因的表达。细胞自噬是一种真核生物中高度保守的代谢过程,可降解脂类等生物大分子物质以供细胞重新利用[27]。因此,自噬深度参与脂肪细胞的细胞分化与能量代谢,若自噬被抑制会导致脂肪的沉积,被激活则脂肪沉积会减少。在本试验研究中,HFD组添加黄连素后提高了PINK1和Atg5基因相对表达量。PINK1被认为是线粒体转运的分子开关,参与线粒体自噬调控过程[28];Atg5是形成自噬复合体的关键基因,影响细胞自噬活性,对细胞的自噬调控有着重要作用。有研究表明,敲除或沉默Atg5可导致脂肪沉积的发生[29]。因此,高脂饲料抑制了花鲈脂肪细胞的自噬,而黄连素的添加激活了脂肪细胞的自噬,最终缓解了脂肪的沉积。
4 结 论
综上所述,高脂饲料会导致花鲈的脂肪过度沉积并损伤肝细胞,添加黄连素通过促进脂肪细胞的凋亡与自噬而缓解脂肪沉积,且黄连素还可以上调肝脏脂肪分解基因的表达,从而促进了脂肪分解;但黄连素会降低花鲈的生长速度和饲料转化率。
猜你喜欢
中华养生保健(2020年5期)2020-11-16
沉积与特提斯地质(2019年1期)2019-07-16
沉积与特提斯地质(2018年4期)2018-07-16
益寿宝典(2017年34期)2017-02-26
中老年健康(2016年10期)2016-11-19
中国运动医学杂志(2016年3期)2016-07-10
南方文学(2016年4期)2016-06-12
中国塑料(2015年11期)2015-10-14
医学研究杂志(2015年12期)2015-06-10
食品工业科技(2014年5期)2014-03-11
