
ω-3多不饱和脂肪酸对畜禽生理功能的影响及其应用的研究进展
2021-10-18 08:20薛永强马永喜
动物营养学报 2021年9期
薛永强 余 苗 马永喜 杨 博*
(1.华南理工大学生物科学与工程学院,广东省脂类科学与应用工程技术研究中心,广州 510006;2.广东省农业科学院动物科学研究所,农业部华南动物营养与饲料重点实验室,畜禽育种国家重点实验室,广东省畜禽育种与营养研究重点实验室,广州 510640;3.中国农业大学农业部饲料工业中心,北京 100193)
在动物营养领域中,脂肪的传统功效主要包括贮存和供应热能。近年来,人们逐步认识到脂肪在动物营养领域中的研究滞后及其功能的重要性。随着对脂肪营养研究的不断深入,脂肪酸尤其是多不饱和脂肪酸的生理功能及开发成为研究热点。脂肪酸是脂肪中最主要的成分,包括饱和脂肪酸和不饱和脂肪酸,而不饱和脂肪酸又分为单不饱和脂肪酸和多不饱和脂肪酸。多不饱和脂肪酸包括ω-3、ω-6、ω-7和ω-9系列多不饱和脂肪酸,其中ω-3多不饱和脂肪酸(ω-3 polyunsaturated fatty acids,ω-3 PUFAs)和ω-6多不饱和脂肪酸(ω-6 polyunsaturated fatty acids,ω-6 PUFAs)是人和动物体必不缺少的脂肪酸,当其缺乏时可引起生长迟缓、生殖障碍、免疫失调、皮肤损伤以及肾脏、肝脏、神经和视觉等方面的多种疾病[1]。
ω-3 PUFAs具有提高动物生产和繁殖性能、免疫调节、改善毛色及蛋壳外观等多种生理功能[2-4],主要包括α-亚麻酸(α-linolenic acid,C18∶3,ALA)、二十碳五烯酸(eicosapentaenoic acid,C20∶5,EPA)和二十二碳六烯酸(docosahexaenoic acid,C22∶6,DHA),其分子结构式见图1,广泛存在于鱼类、藻类生物及部分植物中。自然界中,基本所有油脂中都含有ω-6 PUFAs,而只有少数油脂含有ω-3 PUFAs,因此在动物养殖过程中容易出现ω-6 PUFAs过量而ω-3 PUFAs缺乏的现象,长期下去将造成动物机体代谢紊乱,影响动物机体的健康。本文从ω-3 PUFAs对畜禽繁殖性能、免疫功能、糖和脂代谢的影响及其在畜禽生产中的应用进行综述,以期为ω-3 PUFAs在动物营养领域中的应用提供参考。
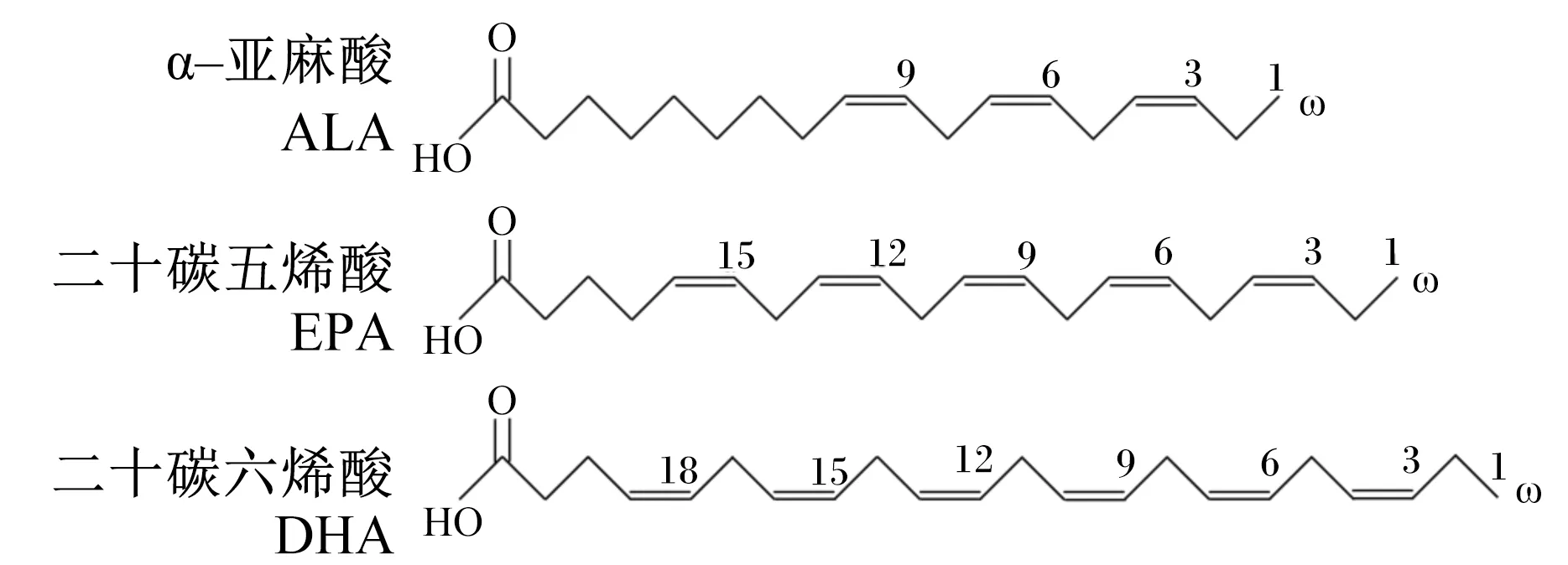
图1 几种ω-3 PUFAs的分子结构式Fig.1 Molecular structural formula of several ω-3 PUFAs
1 ω-3 PUFAs在机体内的合成途径
多不饱和脂肪酸在体内的合成途径如图2所示,ω-3 PUFAs和ω-6 PUFAs在体内的合成共用着同一套酶体系。ω-3 PUFAs和ω-6 PUFAs的起始脂肪酸分别是亚麻酸(C18∶3)和亚油酸(C18∶2),然后二者通过碳链延长、去饱和生成后续的系列脂肪酸,使碳链长度和不饱和度都增加。其中,ω-3 PUFAs中的EPA是白三烯素B5、前列腺素I3和血栓素A3等抗炎类激素的前体物质,而ω-6 PUFAs中的花生四烯酸则是血栓素A2、前列腺素A2和白三烯B4等促炎类激素的前体物质[5],两者起到相反的作用,ω-3 PUFAs与ω-6 PUFAs的比例决定了炎症反应和免疫应答的严重程度。由此可见,ω-3 PUFAs和ω-6 PUFAs在机体内发挥着不同的生理功能特点,甚至在某些方面具有相反的生理活性,因此ω-3 PUFAs和ω-6 PUFAs的平衡对动物的健康至关重要。

图2 多不饱和脂肪酸的合成途径Fig.2 Synthesis pathways of polyunsaturated fatty acids[6]
2 ω-3 PUFAs对畜禽繁殖性能的影响
脂肪对动物繁殖性能的研究早有报道,补充脂肪和能量对动物的生殖具有积极作用,在饲粮中添加脂肪可以提高母猪的繁殖性能等。脂肪酸作为脂肪的主要成分,与动物的健康、生产和繁殖之间密切关联,特别是ω-3 PUFAs与ω-6 PUFAs之间的比例[7]。脂肪对动物繁殖性能的研究更多的集中于脂肪与能量平衡对其繁殖性能的影响。随着对脂肪酸的深入研究,发现脂肪对动物繁殖性能的影响不是简单的调节能量平衡,可能对其具有直接作用[8]。ω-3 PUFAs对动物繁殖性能的影响具有以下几个方面:1)ω-3 PUFAs是合成类十二烷(前列腺素、凝血恶烷、环前列腺素和白三烯等)的前体物质,而类十二烷是一种与激素有类似作用的类激素。在这些类激素中,前列腺素中的前列腺素E2和前列腺素F2α在动物生殖系统中起着主要作用。ω-3 PUFAs可与胚胎发育中的细胞膜磷脂结合,对后代的脂质和类十二烷代谢具有独特的调节作用[9-10]。2)ω-3 PUFAs是一些重要生殖激素合成和代谢的影响因素,参与类固醇激素或性腺激素的生物合成,如糖皮质激素、孕酮、睾酮和雌二醇等。这些激素在动物生殖器官的生长、分化及动物生育能力等方面发挥着重要的作用。ω-3 PUFAs能够降低机体血浆胆固醇的含量,导致类固醇激素的减少,因为胆固醇是孕酮和雌二醇等激素的前体物质[11-12]。3)动物的精子的质膜表面含有很高浓度的ω-3 PUFAs,其中DHA的含量最高[13]。这些不饱和脂肪酸在促进精子成熟、维持膜的流动性、维持细胞功能及精卵结合等方面发挥着重要的作用。ω-3 PUFAs对雄性动物精子膜的完整性、存活率、精子活力、精子数量和密度等有着一定的影响[14-15]。4)ω-3 PUFAs与卵泡细胞的发育、卵母细胞的成熟与质量、早期胚胎质量等均有着密切的联系[16]。当在妊娠期和哺乳期饲喂母猪时,ω-3 PUFAs能够进入卵母细胞、胚胎和胎儿,有利于胎儿的早期发育[17]。由此可见,ω-3 PUFAs通过对一些生殖激素、类固醇激素、生殖器官及生殖细胞(精子和卵母细胞)等的影响,以此影响动物的繁殖性能。
Gokuldas等[18]研究发现,在母猪妊娠早期的饲粮中添加ω-3 PUFAs可以调节前列腺素生物合成途径中关键酶基因的表达,其中,显著上调了前列腺素E合成酶基因的表达,显著下调了前列腺素F合成酶和羰基还原酶1等关键酶基因的表达,对母猪的繁殖性能产生有益影响。Posser等[19]研究发现,给母猪补充富含ω-3 PUFAs的微藻类能够缩短断奶到再发情的间隔,并且有助于提高仔猪的初生重。Otte等[20]研究发现,ω-3 PUFAs能够降低初产母猪卵巢组织中红色素P450(cytochrome P450,CYP)中CYP11A1和CYP19A1的基因表达,说明血清胆固醇含量的降低可能减少类固醇激素生成,其中CYP19A1在卵泡细胞中的表达与卵泡优势机制有关,可能是卵母细胞质量和能力的标志,表明补充ω-3 PUFAs有助于提高雌性动物的繁殖性能。Murphy等[21]研究发现,ω-3 PUFAs能够显著增加公猪精子的精液体积和总精子数量,并且改变了精浆的脂肪酸组成。Feng等[22]通过添加不同ω-3/ω-6 PUFAs(1.00∶18.39、1.00∶7.84、1.00∶5.04、1.00∶4.15和1.00∶2.32),结果显示,ω-3/ω-6 PUFAs为1.00∶4.15时能够显著提高公鸡血清中促性腺激素释放激素、促卵泡激素、促黄体激素和睾酮的含量,并显著提高这些激素相关受体基因的mRNA表达水平,表明ω-3 PUFAs可以提高公鸡的生殖性能。Asl等[4]在罗斯(Ross)肉鸡饲粮中添加不同ω-3/ω-6 PUFAs(分别为0.09、0.16和0.23),结果显示当比例为0.16时可以提高肉鸡的精液品质和繁殖性能,并且补充维生素E可以通过增强抗氧化能力来提高精子活力。以上结果均表明,ω-3 PUFAs对畜禽的繁殖性能有着显著的改善作用,但最佳的ω-3/ω-6 PUFAs还有待进一步的研究。
3 ω-3 PUFAs对畜禽免疫功能的影响
研究表明,ω-3 PUFAs具有良好的抗炎和抗氧化应激作用[23-24],能够调节脂类介质合成、活化内皮细胞、激活白细胞和细胞因子的释放等,缓解机体的过度炎症反应[25]。多不饱和脂肪酸是细胞膜磷脂的结构成分,可通过生成基于脂质的信号介质(如细胞膜上受体和免疫相关分子等)来调节细胞的免疫反应[26]。ω-3 PUFAs对免疫功能影响的可能机制有:1)ω-3 PUFAs可通过改变信号传递、免疫细胞因子的基因表达等来调节细胞因子的分泌,降低一些自身的炎症反应。ω-3 PUFAs还可提高免疫物质的活性,如免疫球蛋白(immunoglobulin,Ig)A、IgG、IgE和IgM等的活性[27],以此提高机体的免疫力。2)ω-3 PUFAs可抑制单核细胞、巨噬细胞和内皮细胞产生肿瘤坏死因子-α(tumor necrosis factor-α,TNF-α)、白细胞介素(interleukin,IL)-1β、IL-6和IL-8等炎症细胞因子。ω-3 PUFAs可抑制活化核转录因子-κB(nuclear factor-κB,NF-κB)信号通路,从而抑制NF-κB的表达,降低炎症反应;提高抗炎因子IL-4、IL-10及过氧化物酶体增殖物激活受体γ(peroxisome proliferator-activated receptor γ,PPARγ)的表达水平[28-30],从而缓解机体的炎症反应及提高免疫力。3)ω-3 PUFAs可抑制树突状细胞上细胞间黏附分子-1(intercellular cell adhesion molecule-1,ICAM-1)、淋巴细胞功能相关抗原-1(lymphocyte function-associated antigen-1,LFA-1)和CD80的表达,抑制辅助性T细胞的表达及细胞毒T细胞的激活[31],改善机体的炎症反应。4)ω-3 PUFAs还可抑制花生四烯酸的代谢,间接降低促炎类激素的生成,如降低花生四烯酸产生的类十二烷酸等介质的产生。5)ω-3 PUFAs中的EPA衍生而来的E类消退素(E-series resolvins,RvE)可在中性粒细胞、巨噬细胞、树突状细胞和T细胞中发挥强大的抗炎作用。由DHA衍生而来的D类消退素(D-series resolvins,RvD)和保护素(protectins)也同样具有良好的抗炎效果[5]。由此可以,ω-3 PUFAs可以通过多种途径来调节机体的免疫功能,调节机制如图3所示。
近年来,集约化模式养殖规模不断扩大,这给动物生产带来了各种应激,如早期断奶应激、运输和环境温湿度等,造成动物的炎症反应及生产性能下降。饲粮添加ω-3 PUFAs可以缓解动物的炎症反应。ω-3 PUFAs可以通过细胞和体液免疫反应来调节免疫系统,改善动物的抗氧化能力、脂质过氧化及免疫应答反应[32],有助于改善动物的生产性能。Huber等[3]研究发现,在低蛋白质饲粮中添加ω-3 PUFAs可以缓解断奶仔猪的炎症反应和应激反应,有助于提高饲料转化率。Duan等[33]研究发现,低ω-6/ω-3 PUFAs饲粮能够显著降低猪血清中促炎细胞因子IL-6和IL-1β的含量,并且显著降低肌肉组织和脂肪组织中的IL-6、IL-1β和TNF-α的mRNA表达水平,表明ω-3 PUFAs可以通过降低炎症因子的表达水平来降低机体的炎症反应。鱼油中含有丰富的ω-3 PUFAs,研究表明在饲粮中添加5%~7%的鱼油能够降低猪血浆中TNF-α、IL-1β和干扰素-γ(interferon-γ,IFN-γ)等细胞因子的含量[34-36],调节机体的炎症反应。在家禽中,Maroufyan等[37]研究发现,随着饲粮中ω-3 PUFAs的富集,肉鸡的脾脏重量、传染性法氏囊病毒和新城疫病毒的抗体滴度、血清IL-2和IFN-γ的含量不断升高,肉鸡的免疫应答增强。Ali等[38]研究发现,在鸡饲粮中添加50g/kg的鱼油,与对照组相比,其血清抗体和IgM和IgG含量显著升高,免疫功能得到提高。总体来说,ω-3 PUFAs可以通过增强免疫物质活性、降低炎症因子表达及提高抗氧化能力等来改善畜禽的免疫功能。

ω-3 PUFAs:ω-3多不饱和脂肪酸 ω-3 polyunsaturated fatty acids,IgA:免疫球蛋白A immunoglobulin A;IgG:免疫球蛋白G immunoglobulin G;IgE:免疫球蛋白E immunoglobulin E;IgM:免疫球蛋白M immunoglobulin M;NF-κB:核转录因子-κB nuclear factor-κB;TNF-α:肿瘤坏死因子-α tumor necrosis factor-α;IL-4:白细胞介素-4 interleukin-4;IL-10:白细胞介素-10 interleukin-10;PPARγ:过氧化物酶体增殖物激活受体γ peroxisome proliferator-activated receptor γ;ICAM-1:细胞间黏附分子-1 intercellular cell adhesion molecule-1;LFA-1:淋巴细胞功能相关抗原-1 lymphocyte function-associated antigen-1;EPA:二十碳五烯酸 eicosapentaenoic acid;DHA:二十二碳六烯酸 docosahexaenoic acid;RvE:E类消退素 E-series resolvins;RvD:D类消退素 D-series resolvins;Protectins:保护素。↑表示升高,↓表示降低。↑ mean increased, and ↓ mean decreased.
4 ω-3 PUFAs对畜禽糖、脂代谢的影响
长期饲喂高脂饲粮将导致动物机体糖、脂代谢紊乱以及胰岛素抵抗等,造成多种代谢性疾病[39]。脂肪酸作为脂质的主要组成成分,对糖、脂代谢有着重要的调节作用。增加ω-3 PUFAs的摄入量能够降低血糖、胰岛素、胰岛素抵抗指数、甘油三酯及低密度脂蛋白胆固醇等的含量,且能够提高脂联素等脂肪细胞因子的水平[39-40]。脂联素是脂肪细胞分泌的一种脂肪细胞因子,在改善机体糖脂代谢、炎症反应及氧化应激等方面起着重要的作用[41]。补充鱼油(含EPA和DHA),可以改善高脂诱导产生的肌肉组织和脂肪组织内脂质稳态的失衡[42]。ω-3 PUFAs对糖、脂代谢的调控机制:1)ω-3 PUFAs可促进脂肪酸的β-氧化,加强脂肪动员,增强产热,从而降低体脂的沉积;2)ω-3 PUFAs通过降低生脂酶如乙酰辅酶A羧化酶(acetyl CoA carboxylase,ACC)和提高激素敏感性脂肪酶(hormone-sensitive lipase,HSL)的活性[43],以及降低脂类生成相关基因和蛋白的表达水平[33],从而抑制脂质的合成,改善机体的脂质代谢;3)ω-3 PUFAs通过调节脂肪组织分泌的脂肪细胞因子如瘦素、抵抗素和脂联素等影响机体的糖、脂代谢[39];4)ω-3 PUFAs可能通过调节线粒体的功能和内质网应激来影响机体的胰岛素敏感性,调节机体的糖、脂代谢[44]。ω-3 PUFAs是一种重要的生物活性脂类,可通过对脂肪酸氧化、糖和脂质代谢相关酶、脂肪细胞因子及胰岛素敏感性等的调节影响机体的糖和脂代谢,在动物饲粮中添加适量的ω-3 PUFAs有利于机体正常的糖、脂代谢,其作为一种重要的营养素,对人和动物健康有着显著的有益作用。
Song等[45]研究表明,ω-3 PUFAs能够降低育肥猪脂肪组织中ACC基因的表达水平,并提高HSL基因的表达水平,并且降低血浆中胆固醇的含量,表明ω-3 PUFAs可以增强脂肪组织中游离脂肪酸的释放,调节储存在脂肪组织中脂肪酸的脂解和动员。Nong等[46]研究发现,随着饲粮中ω-3 PUFAs比例的增加(ω-6/ω-3 PUFAs为8∶1、5∶1和3∶1),猪皮下脂肪组织中的甘油三酯和总胆固醇含量显著降低,并显著上调HSL、肉毒碱棕榈酰转移酶1(carnitine palmitoyltransferase 1,CPT1)和脂肪酸结合蛋白4(fatty acid binding protein 4,FABP4)等基因的表达水平,但下调了脂肪甘油三酯脂肪酶(adipose triglyceride lipase,ATGL)基因的表达水平,表明ω-3 PUFAs能够增强脂解作用并影响脂肪组织中的脂肪酸代谢。Posser等[19]在妊娠85 d的母猪中研究发现,提供14和28 g/d的微藻(富含ω-3 PUFAs)可以显著降低母猪妊娠期血浆甘油三酯的含量,并提高了仔猪的初生重。Moreira等[47]研究同样发现,在后背母猪饲粮中添加鱼油,能够降低血清胆固醇的含量,降低的原因可能与卵母细胞瘦素染色增强有关。Neijat等[49]研究表明,饲粮添加ω-3 PUFAs(0.6%的亚麻籽油和藻类DHA)显著下调了蛋鸡肝脏中过氧化物酶体增殖物激活受体α(peroxisome proliferator-activated receptor α,PPARα)、肉毒碱棕榈酰转移酶1A(carnitine palmitoyl transferase 1A,CPT1A)、酰基辅酶A氧化酶1(acyl-CoA oxidase 1,ACOX1)和脂肪酸去饱和酶2(fatty acid desaturase 2,FADS2)等脂类代谢相关基因的表达水平,调节蛋鸡肝脏脂类代谢。麻丽坤等[49]研究发现,蛋鸡饲粮添加3%的鱼油可以显著降低血清总胆固醇的含量。上述结果均表明,ω-3 PUFAs能够调节畜禽脂质代谢,在饲粮中补充适量的ω-3 PUFAs可以改善动物的脂质代谢,提高动物机体免疫机能并产生有益作用。
5 ω-3 PUFAs在畜禽生产中的应用
在过去几十年里,饲粮中应用的油脂主要是大豆油、玉米油等,而这些油脂中含有大量的ω-6 PUFAs,造成ω-6 PUFAs和ω-3 PUFAs的失衡。ω-3 PUFAs与心血管疾病、抑郁症、肥胖及癌症等多方面密切关联[50-53],高含量的ω-3 PUFAs和低ω-6/ω-3 PUFAs更有利于人体健康,富含ω-3 PUFAs的畜产品也受到更多消费者的青睐。饲粮中的ω-3 PUFAs是畜产品沉积ω-3 PUFAs的主要方式[54],因此通过对饲粮ω-3 PUFAs的含量调控改善畜产品品质也是现阶段的研究热点。
5.1 改善畜禽的生产性能
亚麻籽油、鱼油和微藻类等功能性原料富含ω-3 PUFAs,在畜禽饲粮中添加适量的多不饱和脂肪酸有助于改善畜禽的生产性能。ω-3 PUFAs对畜禽生产性能的影响见表1。

表1 ω-3 PUFAs对畜禽生产性能的影响Table 1 Effects of ω-3 PUFAs on livestock and poultry performance

续表1试验动物Experimental animals生长阶段Growth stage试验设计Experimental design试验结果Experimental results参考文献References母猪 Sows妊娠期45 d至哺乳期21 d分7组:棕榈油组、0.5%亚麻油组、2.0%亚麻油组、0.5%鱼油组、2.0%鱼油组、0.5%亚麻油+0.5%鱼油组和0.5%亚麻油+1.0%鱼油组饲喂亚麻籽油的母猪比饲喂鱼油或棕榈油的母猪多出1.3和1.5头活产仔猪,但对仔猪的初生重和窝重无影响Tanghe等[61]北京油鸡 Beijing Youchicken1日龄添加0%、0.12%、0.42%、1.00%和1.97%亚麻油使ω-6/ω-3 PUFAs分别为30∶1、20∶1、10∶1、5∶1和2.5∶1,为期13周相比ω-6/ω-3 PUFAs为30∶1组,ω-6/ω-3 PUFAs为5∶1组的平均日增重提高10.22%,料重比降低12.21%,各亚麻油添加组的死淘数均降低Qi等[62]罗斯肉鸡 Ross broilers1日龄向日葵油组(ω-3/ω-6 PUFAs≈0)、向日葵油+大豆油组(向日葵油∶大豆油=1∶1)和大豆油组,为期35 d相比向日葵油组,大豆油组的平均日增重提高6.70%,料重比下降6.22%Jalali等[63]北京油鸡 Beijing Youchicken10周龄对照组(Ⅰ组)、9%亚麻籽组(Ⅱ组)和9%亚麻籽+200 mg/kg维生素E组(Ⅲ组),为期8周相比于Ⅰ组,Ⅱ组和Ⅲ组的阶段日增重分别提高11.56%和20.27%,阶段日采食量分别提高5.95%和8.36%,料重比分别下降5.20%和10.41%。赵丹阳等[64]
一般而言,饲粮中添加富含ω-3 PUFAs原料对畜禽的生产性能均具有改善作用。然而也有研究发现,补充ω-3 PUFAs对畜禽的生产性能没有显著的影响[2,65],造成这样的原因可能与饲粮组成、饲喂周期、多不饱和脂肪酸的类型、ω-3 PUFAs含量或ω-6/ω-3 PUFAs有关。因此,今后的研究应更多集中于对ω-6/ω-3 PUFAs以确定畜禽不同阶段、不同品种对多不饱和脂肪酸的具体需求量。值得注意的是ω-3 PUFAs易被氧化,产生过氧化物、氢氧化物、醛和酮等有害物质,对动物造成不同程度的危害。通常通过饲粮添加维生素E、硒等来提高其抗氧化能力,因此,如何有效地控制ω-3 PUFAs氧化稳定性也是当前研究中亟待解决的问题。
5.2 提高畜禽的产品品质
近年来,随着人们生活质量的不断提高,人们对肉质及相关畜产品品质的需求也愈来愈高,开发富含ω-3 PUFAs的产品,有助于改善人类健康和生活水平。饲粮中的脂肪酸组成在调节畜禽产品的营养品质方面起着重要的作用,尤其是多不饱和脂肪酸。Nong等[46]研究发现,低ω-6/ω-3 PUFAs(8∶1、5∶1和3∶1)显著增加了中国地方猪皮下脂肪组织中ω-3 PUFAs含量(3.04% vs. 3.97% vs. 5.43%),并且显著降低了皮下脂肪组织中ω-6/ω-3 PUFAs(5.06 vs. 3.68 vs. 3.02),表明低ω-6/ω-3 PUFAs可以调节猪肉的品质,并促进ω-3 PUFAs在肉中的沉积。Leikus等[56]研究表明,饲粮添加亚麻籽油显著提高猪腰髂肋肌肉中的ALA、EPA和总ω-3 PUFAs的含量,对改善猪肉中不饱和脂肪酸和饱和脂肪酸比例和ω-6/ω-3 PUFAs具有积极的影响。Song等[45]研究发现,饲粮添加亚麻籽油(ω-6/ω-3 PUFAs分别为18∶1、4∶1和2∶1)可显著增加猪背最长肌中的ALA(0.91% vs. 3.51% vs. 4.38%)和总不饱和脂肪酸(14.54% vs. 17.75% vs. 19.13%)含量,降低ω-6/ω-3 PUFAs(15.81 vs. 4.06 vs. 3.41),并且降低猪肉的蒸煮损失,改善肉品质。Khatibjoo等[2]研究发现,饲粮中ω-6/ω-3 PUFAs与蛋黄中ω-3 PUFAs含量呈显著正相关,低ω-6/ω-3 PUFAs可以显著增加蛋黄中的DHA、EPA和总ω-3 PUFAs的含量。在蛋鸡饲粮中添加花生油、鱼油、红花油、亚麻籽、鱼粉或藻类等油脂,可提高鸡蛋中ω-3 PUFAs的含量[32]。Qi等[62]研究发现,饲粮中ω-3 PUFAs替代ω-6 PUFAs对鸡的皮下脂肪、肌内脂肪含量及肉品质均有着显著的改善作用。且禽肉中的ω-3 PUFAs含量可以通过增加饲粮中ω-3 PUFAs的含量来改善[66]。由此可见,通过对饲粮中ω-3 PUFAs的组成和含量的调节来生产富含ω-3 PUFAs的产品,对生产具有特定营养功能的动物产品有着重要的意义。
6 小 结
ω-3 PUFAs与动物的繁殖性能、免疫功能及机体代谢之间存在着密切的关系,与动物健康和动物产品品质相关,在人和动物体内发挥着重要的生理功能。研究发现,在畜禽饲粮中添加适量的ω-3 PUFAs可以提高畜禽的繁殖性能,改善畜禽的免疫功能,调节糖和脂代谢,改善畜禽产品品质。如何有效控制ω-3 PUFAs氧化稳定性,在畜禽的不同生长阶段、不同品种对饲粮ω-3PUFAs含量及ω-6/ω-3 PUFAs比例的研究还相对缺乏。探究ω-3 PUFAs对畜禽生理功能影响的作用机制将成为今后的研究趋势。加强ω-3 PUFAs在动物饲粮中的应用,在开发特定的动物产品方面有很大的应用价值,具有重要的理论和现实意义。
猜你喜欢
动物营养学报(2022年8期)2022-09-01
中国饲料(2021年17期)2021-11-02
中国生殖健康(2020年5期)2021-01-18
中成药(2018年12期)2018-12-29
中国生殖健康(2018年5期)2018-11-06
天然产物研究与开发(2018年9期)2018-10-08
中学科技(2016年5期)2016-05-12
合成化学(2015年10期)2016-01-17
动物营养学报(2015年9期)2016-01-07
中国洗涤用品工业(2015年8期)2015-02-28
