
大叶金顶杜鹃繁育系统研究
2021-10-14 01:59余海清帅伟姜欣华刘燕云廖成云姬慧娟马文宝
四川林业科技 2021年5期
余海清,帅伟,姜欣华,刘燕云,廖成云,姬慧娟,马文宝*
1.甘孜藏族自治州林业科学研究所,四川康定 626001;
2.四川省林业调查规划院四川成都 610081;
3.四川省林业科学研究院,森林和湿地生态恢复与保育四川重点实验室,四川成都 610081
繁育系统是植物研究中较受关注的领域之一,植物的花部综合特征、花部各性器官、花开放式样、自交亲和程度及交配系统等都与其繁育系统密切相关[1]。植物繁育过程中与传粉者及传粉行为一起决定了植物后代的遗传组成和适合度[2],是种群有性生殖的纽带[3,4]。植物繁育系统已成为一个以“生殖”为核心,以探讨物种多样性发生历史、维持机理和保护策略为最终目的的综合交叉研究[5]。
杜鹃花是世界四大园林花卉之一,被称为“木本花卉之王”,中国杜鹃属植物有576种,川西南地区是杜鹃属的现代分布中心与多度中心之一[6]。杜鹃花属被认为是适应辐射的一个大属,在我国西南山地尤其是横断山区域特有种比例非常高,为探讨生态成种提供了一个很好的研究系统[7]。目前,国内外研究学者对不同亚属不同亚组的杜鹃花物种开展了繁育系统研究,如常绿杜鹃花亚属中云锦杜鹃花亚组的云锦杜鹃花[8]和大白杜鹃花[9]、长序杜鹃花亚组的R. ponticum[10,11]和R.aureum[12]、树形杜鹃花亚组的树形杜鹃R.arboreum[13]和露珠杜鹃花亚组马雄杜鹃花[14];杜鹃花亚属三花杜鹃亚组的绣叶杜鹃[15]、多鳞杜鹃[16]、有鳞大花组的大喇叭杜鹃花[17]和高山杜鹃组R. ferrugineum[18];异蕊杜鹃花亚属R.semibarbatum[19];羊踯躅亚属R.canadense[20]等4个亚属7亚组(组)的杜鹃花物种的繁育系统进行了研究,但是对于常绿杜鹃花亚属大理杜鹃花亚组植物以及在野外条件下原生种,特别是横断山区域特有杜鹃花物种的繁殖生物学方面研究相对较少[9,14,15,16]。
大叶金顶杜鹃R. prattii(康定杜鹃花)为常绿杜鹃花亚属Subg.Hymenanthes大理杜鹃花亚组Subsect.Taliensia的灌木或小乔木,中国特有种,产于四川西部、西南部和西北部[21],生于亚高山暗针叶林林缘、灌丛,分布海拔2800~4000 m,在叶形和花部形状等与金顶杜鹃区别明显,耿玉英已恢复其为种等级[6],大叶金顶杜鹃具有花朵大、花色艳丽、植株大型、群落优势明显等特点,是优良的杂交亲本种质资源。本研究以四川省甘孜州泸定县贡嘎山自然保护区内自然分布的大叶金顶杜鹃为研究对象,观察开花物候、花部综合特征和昆虫访花行为,统计了杂交指数、花粉-胚珠比,进行了人工控制授粉试验,为丰富常绿杜鹃花亚属大理杜鹃花亚组植物繁育系统的基础数据和四川省特有野生杜鹃花资源开发利用及新品种选育提供理论依据。
1 研究地点
研究地点位于四川省甘孜藏族自治州贡嘎山自然保护区雅家埂雅家情海附近,N29°51'57",E102°2'16",海拔3820 m。该区地处中国东部亚热带湿润季风区与青藏高原寒冷气候的过渡带,多年平均气温3.8℃,最冷月(1月)和最热月(7月)平均温度分别为−4.3℃和11.9℃,年平均降水量1940 mm,年平均蒸发量1578.6 mm,年平均空气相对湿度90.2%,土壤为山地暗棕壤,土层厚度40~80 cm,含沙量高、渗透性强,呈弱酸性。
该地区植被以寒温带灌丛为主,其生活型组成以小乔木和中小灌木为主,伴生种类主要为各种杜鹃花属植物,包括无柄杜鹃(R.watsonii)、陇蜀杜鹃(R. przewalskii)、栎叶杜鹃(R. phaeochrysum)、山光杜鹃(R.oreodoxa)、问客杜鹃(R.ambiguum)和毛蕊杜鹃(R.websterianum)等,其他木本和草本植物有峨眉冷杉(Abies fabri)、川滇高山栎(Quercus aquifolioides)、西康花楸(Sorbus prattii)和全缘叶绿绒蒿(Meconopsisintegrifolia)等。
2 试验方法
2.1 花部综合特征及开花动态观察
2019~2020年连续两年对野外居群的大叶金顶杜鹃R. prattii进行了群体花期观察;随机选取30株生长健壮、树形高大的大叶金顶杜鹃植株标记为研究样株,对个体和花序的花期观察,并记录相对应的开花时间。
随机标记花蕾期300朵,花开放后,每天取下20朵,每天对单花开花动态进行观测,记录花冠颜色、花粉散粉、花气味、花粉活力、柱头可授性、花冠长、花管长、内径、花开口宽、花丝长、花柱长、柱头和花药的相对位置。
2.2 大叶金顶杜鹃的花粉活力和柱头可授性
每天从采下20朵花中随机选3朵用来测定单花的花粉活力和柱头可授性。
2.2.1 花粉活力
花粉活力采用TCC法测定[22]。将花粉置于小型离心管中用TCC染液进行染色,置于40℃的恒温水中放置15~30 min后,将其染液置于载玻片上,在光学显微镜下观察。花粉被染成红色(活力较强)和淡红色(活力较弱)为有活力花粉,无色则为无活力的花粉。
花粉活力=(红色花粉数+淡红色花粉数)/观察花粉总数×100%。
2.2.2 柱头可授性
采用联苯胺-过氧化氢法[23]检测柱头可授性。随机标记发育阶段一致的花,开花后每隔1 d取10朵花的花柱,在凹面载玻片中加入体积比为1%联苯胺∶3%过氧化氢∶水=4∶11∶22的反应液,将柱头浸入其中,观察柱头周围是否呈现蓝色并产生大量气泡,根据气泡的数量确定其柱头可授性的强弱。
2.3 大叶金顶杜鹃的繁育系统
大叶金顶杜鹃的繁育系统依据Cruden标准[24]、Dafni标准[23]和套袋试验结果进行综合判定。
2.3.1 花粉量及花粉-胚珠数(P/O)的测定
随机取将开放的花朵的花药5个置于干净的玻璃瓶中,在花药开裂后,加入15%的葡萄糖溶液2 mL,充分摇匀后,取10μL于显微镜下数出花粉粒数N,即可得每朵花的花粉量=200×N,同时取子房在体视显微镜下剖开,统计胚珠数,即可得P/O。以上重复测定10次[25]。
2.3.2 杂交指数估算
按照Dafni的标准进行花朵大小及开花行为的测量及繁育系统的评判。具体方法是:①花朵直径
2.3.3 套袋试验
对大叶金顶杜鹃植株分别进行套袋实验,A.不套袋、不去雄:自由传粉,用于检测自然条件下的传粉情况;B.不去雄套袋:自发的自花传粉,检测是否存在自花授粉;C.去雄授同株异花:开花前去雄,同株异花之间人工传粉,检测自交是否亲和;D.去雄授异株异花:人工异株异花传粉,用不同植株的花粉进行异花授粉后用牛皮纸套袋,检测杂交是否亲和;E.不套袋 去雄:自然条件下的异花传粉,自由传粉;F.套袋 去雄:开花前用牛皮纸套袋、去雄、不授粉,检测无融合生殖。
花期结束后,统计每个处理的坐果率。坐果率(%)=结实数/处理的花数×100%。
2.4 大叶金顶杜鹃的访花昆虫特性
对大叶金顶杜鹃种群的访花昆虫进行调查,随机标记已开放的花朵40朵,从9∶00—17∶00进行连续3 d的观察,记录其访花昆虫种类、访花行为和活动频率。
3 数据统计与分析
所有测量数据均用Excel 2007进行统计分析,测量结果均用平均值±标准差表示;使用SPSS 22.0软件对花冠长、花管长、内径、花开口宽等花部形态特征和坐果率等进行单因素方差分析。
4 试验结果
4.1 开花物候
大叶金顶杜鹃群体5月初始花至6月中下旬结束,花期持续48~55 d左右,单株花期18~32 d,花序水平花期13~16 d,单花开花持续时间为10~11 d。
4.2 花部特征及开花动态
大叶金顶杜鹃为伞形总状花序,花6~10朵,花冠钟形,粉红色,内面基部有深红色斑点,裂片5,花丝10~12,花药黑色,椭圆形孔裂式散粉,雌蕊1枚,雌蕊比花冠略短,高于雄蕊,柱头小,黄绿色(见图1)。
根据大叶金顶杜鹃单花开花周期、花部特征动态和开花动态,将单花开放进程分为花蕾期、初花期、盛花期和凋谢期4个时期(见图1和表1)。花蕾期至凋谢期花瓣颜色由粉红色逐渐变淡至白色;花蕾期花粉便开始线性散粉,柱头弯曲具有少量黏液,初花期和盛花期花药大量线性散粉,柱头直立分泌大量黏液,凋谢期花药散粉和柱头分泌黏液逐渐减少;花蕾期时开始分泌花蜜,随着开花进程蜜量逐渐较少,整个过程无气味(见表1)。

图1 大叶金顶杜鹃开花周期Fig.1 Flowering stageof Rhododendron prattii
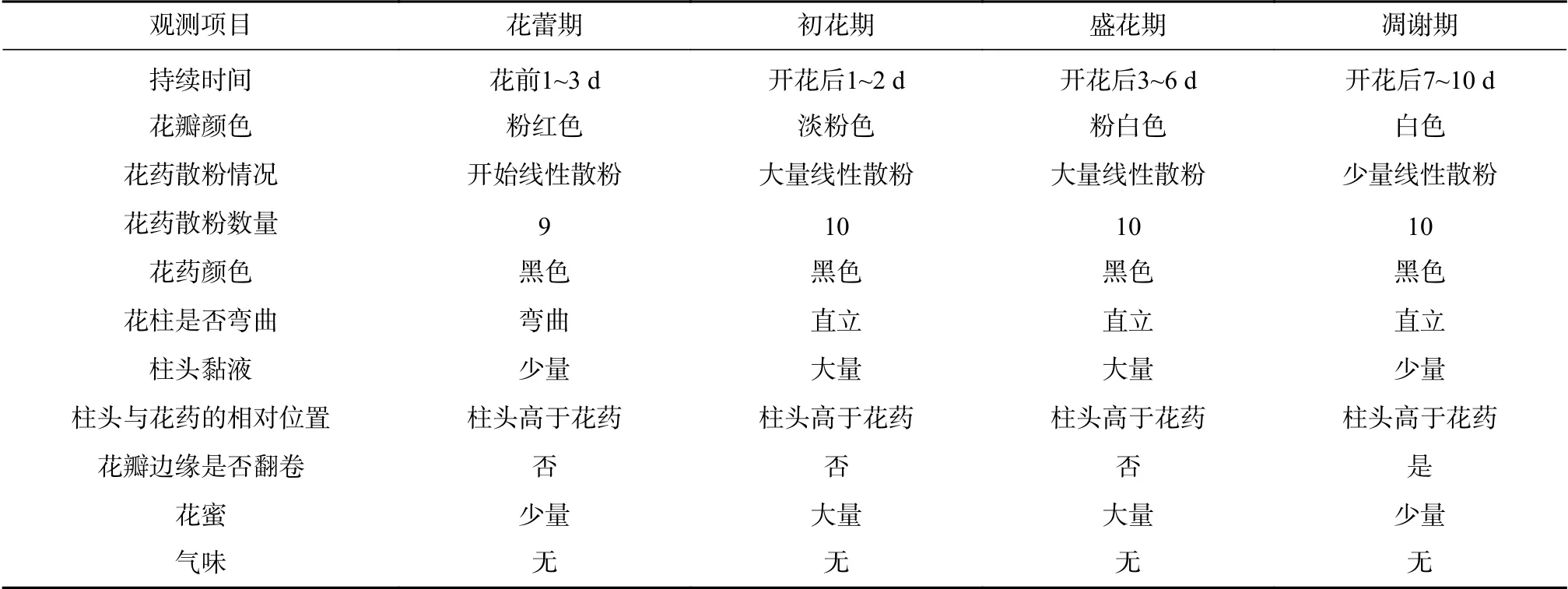
表1 大叶金顶杜鹃单花的开花动态Tab.1 Flowering dynamicsof singleflower of R. prattii
随着大叶金顶杜鹃蕾膨大开花,其花柱、花丝、子房均持续生长,花蕾期与初花期、盛花期和凋谢期的最短花丝和最长花丝表现出显著差异;不同时期间花柱长无差异;柱头始终位于花药群上方,花药和柱头空间最短距离下降趋势明显,花蕾期和凋谢期的花药和柱头空间最短距离呈现出显著差异,子房长显著增长(见表2)。
4.3 花粉活力和柱头可授性
大叶金顶杜鹃花粉活力在自然条件下持续时间为10 d左右。大叶金顶杜鹃蕾期花粉活力为58%,初花期花粉活力维持在60%左右;盛花期花粉活力最高,达95%以上;随后缓慢下降,至凋谢期(第8 d)花粉活力仍保持58%,第9 d快速下降,至第10 d时花粉活力仍还有15%(见图2),与花部动态中花粉散粉情况保持一致(见表2)。
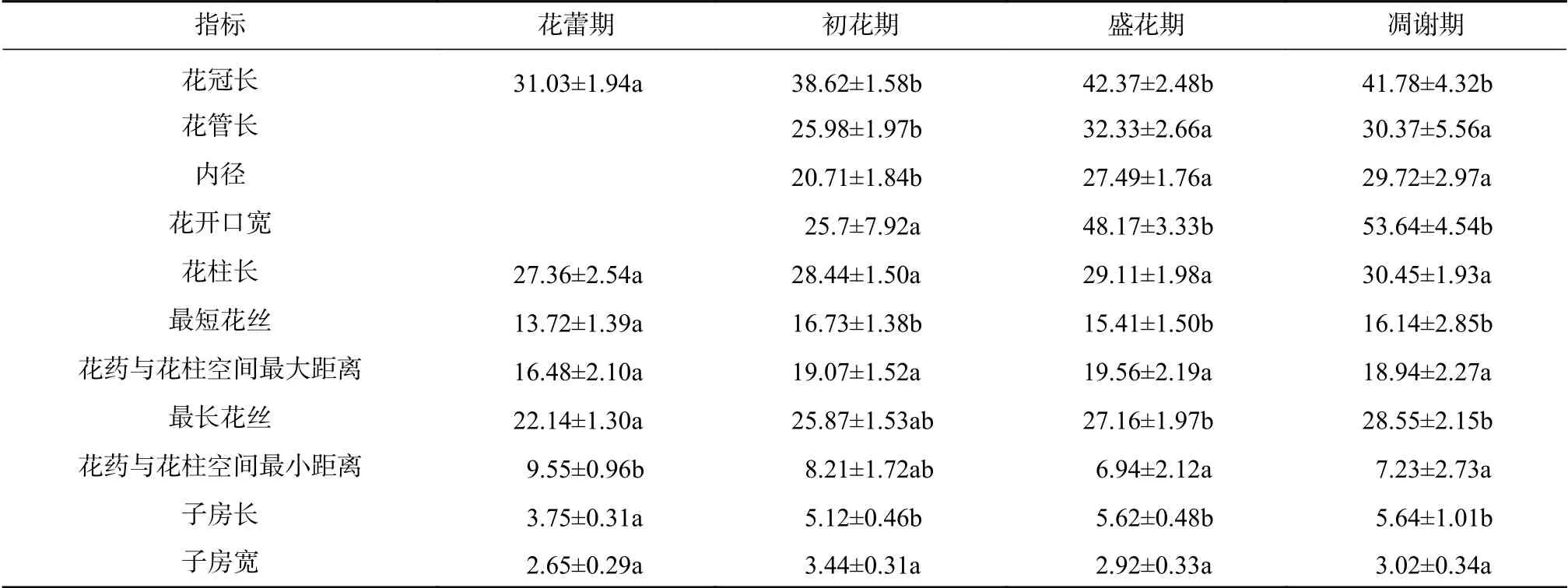
表2 大叶金顶杜鹃的花部综合特征动态变化Tab.2 Dynamic changesof flower comprehensive characteristicsof R. prattii
大叶金顶杜鹃在花蕾期花柱头周围便有气泡出现,可授性相对较弱;盛花期和凋谢期(第7 d和第8 d)柱头周围呈现蓝色并产生大量气泡,可授性最强;凋谢期第9 d开始柱头萎蔫,但仍具有可授性(见图2)。
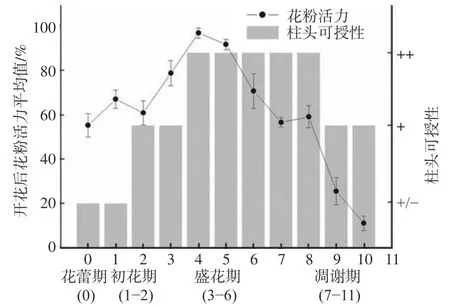
图2 大叶金顶杜鹃粉活力和柱头可授性日变化Fig.2 Diurnal variation of pollen activity and stigma receptivity of R. prattii
4.4 繁育系统
4.4.1 花粉量及花粉-胚珠数(P/O)
根据Cruden的标准,大叶金顶杜鹃繁育系统属于兼性异交(见表3),意味着异交程度不断上升。

表3 大叶金顶杜鹃的花粉-胚珠比(平均值±标准差)Tab.3 Pollen-ovuleratio estimate of R. prattii (Mean ± S.D.)
4.4.2 杂交指数
大叶金顶杜鹃杂交指数表明(见表4),其杂交指数(OCI)为4,故其繁育系统为异交,部分自交亲和,需要传粉者。

表4 大叶金顶杜鹃杂交指数观测结果Tab.4 Hybridization index of R. prattii
4.4.3 套袋试验
从大叶金顶杜鹃植株套袋结果发现,人工异株异花传粉坐果率为97.5%,说明大叶金顶杜鹃异交率最高,与其他套袋实验坐果率呈现出显著差异;在自然条件下的传粉其坐果率为71.5%,自然条件下异株异花和同株异花的坐果率为54.7%;不去雄自花花粉授粉主动自交坐果率为23.1%,授同株异花花粉其坐果率为55.5%,自然状态下、同株异花和自然条件下异株异花和同株异花的坐果率不存在显著差异,但与自花授粉的坐果率相比差异显著,说明大叶金顶杜鹃异交,自交亲和,需要传粉者;大叶金顶杜鹃不存在无融合生殖现象(见图3)。
4.5 昆虫访花特性
大叶金顶杜鹃的访花媒介主要为双翅目食蚜蝇类(成虫)和蝇类(成虫)、膜翅目的叶峰类(成虫),蛛形纲(蜘蛛)、象甲类(成虫)等。整个花期过程中,花蕾期和凋谢期昆虫数量相对较少,盛花期是访花昆虫的活动高峰时期。观察中发现双翅目的食蚜蝇类(成虫)和蝇类(成虫)在中午11:00—13:00其访花频率最高,有效传粉昆虫主要以食蚜蝇类(成虫)为主,蝇类主要针对凋谢期的花朵进行传粉,与柱头不发生有效接触,不是有效的传粉昆虫。
鞘翅目象甲类的访花行为表现为在大叶金顶杜鹃的花蕾上打洞,进入蜜腺部位,从基部开始啃食花丝、花柱,导致花败育。叶蜂类会在大叶金顶杜鹃中停留,但未观察到其携带花粉。蜘蛛类常在大叶金顶杜鹃中爬行或在花丛中织网,未见其觅食花蜜或花粉(见图4)。
5 结论与讨论
在繁育系统的演化中,与之相适应的重要特点是花部特征及花生物学特性的协调进化[8]。大叶金顶杜鹃具有异交(虫媒传粉)花部综合特征,花大、色彩艳丽,能够吸引昆虫为其进行传粉,食蚜蝇类(成虫)是其的有效传粉昆虫。大叶金顶杜鹃的雌、雄蕊具有明显的空间异位,在4个时期中柱头始终处于雄蕊上方(见表1),是典型的雌雄异位植物,这种空间布置可避免自花授粉,是植物避免自交的一种机制[26]。
但与此同时,由于其分布海拔较高,天气变化无常导致其缺乏足够有效的传粉昆虫(见图4),其也具备了自交的花部综合特征。大叶金顶杜鹃单花期长10~12 d,其在花蕾期和开花后8 d内均保持较高活力(58%以上),说明了该物种雄性适合度较高,生殖持续能力较强。与此同时,其柱头从花蕾期就有少量分泌黏液,从盛花期第4 d到凋谢期期第8 d柱头具有较强可授性,二者成熟时间同步性高(见图2),为其自交提供了有利条件。
根据花粉-胚珠比(见表3)、杂交指数(见表4)、传粉昆虫观察(见图4)以及套袋实验结果(见图3)来看,其异株异花传粉坐果率达到97.5%,显著高于其他套袋结果,与绣叶杜鹃的同株异花的坐果率(36.3%)[15]相比,大叶金顶杜鹃花同株异花和自花授粉的坐果率分别为55.5%和23.1%,表现出异交,并具有较高自交亲和性。同时,大叶金顶杜鹃的盛花期花药孔裂散出线状的花粉,凋谢期花冠翻卷并呈下垂状,从基部与花托分开,花瓣(连同雄蕊)从子房前移到花柱,使原来略为上翘的花柱与柱头下垂,部分花粉粘附到柱头上,借助花瓣脱落实现花后授粉(自花授粉),可能是对传粉媒介短缺的一种适应,是其对恶劣生境的一种繁殖保障机制。这个结果也和同为常绿杜鹃花亚属的R. ponticum[10,11]、R.aureum[12],R.arboreum[13]以及杜鹃花亚属的绣叶杜鹃花[15]、马雄杜鹃花[14]的研究结果一致。因此,我们认为大叶金顶杜鹃花的繁育系统是异交,自交亲和,需传粉者。
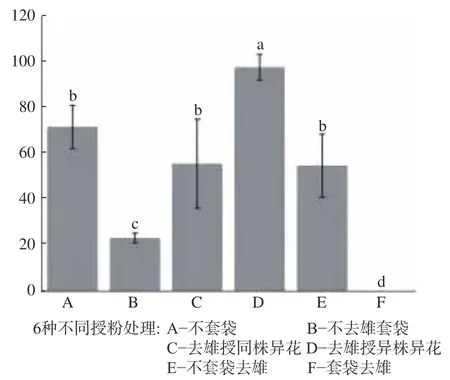
图3 大叶金顶杜鹃坐果率统计结果Fig.3 Statistical resultsof seed setting rateof R. prattii

图4 传粉媒介对大叶金顶杜鹃的访花行为Fig.4 Flower visiting behavior of pollination media to R. prattii
尽管植物的自交在占据新生境,克服高山传粉媒介匮乏,增强植物种群的局部适应性,获得父母本优良性状等方面具有一定的优势,但R.aureum由于在末花期自交率增加,其坐果率下降,种子产量减少,种子败育率增大[12]。云锦杜鹃长期自交引起近交衰退,导致后代适应性下降,云锦杜鹃种群的明显衰退现象[8]。我们也发现大叶金顶杜鹃种群有明显衰退现象,是否在凋谢期的补充自交造成了其种子适合度下降还有待于进一步研究。同时,大叶金顶杜鹃异株异交套袋结果高达97.5%,为杜鹃花杂交育种工作提供了良好杜鹃花的母本,项目组通过与低海拔分布的大白杜鹃等进行杂交育种,已获得部分杂交种幼苗,可为低海拔地区的杜鹃花新品种创制奠定基础。
猜你喜欢
中国科学探险(2022年8期)2022-12-31
安徽农业科学(2022年18期)2022-10-13
新疆农垦科技(2022年1期)2022-03-10
宝藏(2021年1期)2021-03-10
新疆大学学报(自然科学版)(中英文)(2020年2期)2020-07-25
大科技·百科新说(2018年3期)2018-04-11
百姓生活(2018年1期)2018-01-15
通俗小说报(2009年11期)2009-12-23
