
大熊猫主食竹叶围细菌多样性的季节性变化
2021-10-14 01:59田春洋洪明生龙珏洁谢建妹
四川林业科技 2021年5期
田春洋,洪明生,龙珏洁,谢建妹*
1.四川栗子坪国家级自然保护区管理局,四川雅安 625400;
2.西南野生动植物资源保护教育部重点实验室,西华师范大学生命科学学院,四川南充 637009
大熊猫(Ailuropoda melanoleuca)是中国的特有濒危物种,是典型的森林土著物种,加上它憨态可掬的外貌特征,受到了世界人民的喜爱。由于道路建设、森林采伐、竹子开花、采药以及放牧等干扰因素的影响,现存的1864只野生个体被分割于中国西南部的秦岭、岷山、邛崃、大相岭、小相岭和凉山等六大山系中[1-5]。竹子作为大熊猫主要的食物来源,是大熊猫保护的关键之一。四川省大熊猫栖息地内大熊猫可采食的竹类有7属32种,其中小相岭山系大熊猫栖息地内大熊猫可采食竹类3属7种,其中面积最大的为峨热竹(Arundinaria spanostachya),占该山系大熊猫栖息地内大熊猫取食竹总面积的38.08%[6]。由于道路建设等因素,该小相岭山系的大熊猫局域种群形成了孤立的小种群,易受到外来因素的干扰而灭绝[6]。
四川栗子坪国家级自然保护区位于小相岭山系,行政区域上位于石棉县南部,地理坐标为北纬28°51'02″—29°08'42″,东经102°10'33″—102°29'07″。整个保护区占地面积为47 885 km2。保护区内最高的海拔为4 551 m,最低的海拔为1 330 m,地势西南高、东北低。区内的年平均温度为11.7℃~14.4℃,年平均降水量为800~1250 mm。随着海拔的升高,区内植被类型从常绿阔叶林变为针叶林,在高海拔区域分布的植被类型有高山灌丛、草甸、流石滩等[3]。保护区内可供大熊猫取食的竹类有峨热竹、石棉玉山竹(Yushania lineolata)、丰实箭竹(Fargesia ferax)等,除冬季少量采食石棉玉山竹外,该区域的大熊猫全年主要采食峨热竹,该竹类主要分布在区内海拔约2800 m以上的区域[4]。
在植物叶部周围寄生或是附着有大量的微生物,这些微生物有真菌、细菌等[7]。在植物叶围微生物中,细菌的丰富度最高,并与植物叶面形成了一个微生物-植物相互作用的微生态系统[7]。叶面微生物不仅影响植物的生长,还能通过竞争养分和拮抗作用等来抑制植物表面病原菌的生长,对于防治植物病害有极大的作用,然而存在的致病菌也会导致植物患病[7−9]。例如,鞘脂杆菌属(Sphingobium)和鞘氨醇单胞菌(Sphingomonas)的一些物种被证明能降解很多有机污染物[10−11];甲基杆菌属(Methylobacterium)和假单胞杆菌属(Pseudomonas)的物种被报道有植物生长促进剂的潜能[12−14]。影响叶围微生物群落的因素非常多,就生物因素而言,有植物种类、基因型等,例如拟南芥单基因lacs2和pec1突变能改变其叶际微生物的组成[15−16]。就非生物因素而言,紫外压力、空气湿度、地理位置、生长季节等影响叶围微生物组成、多样性等,例如地理位置被证明是大田莴苣微生物的主要驱动因子[17]。因此,叶围微生物的组成、多样性等对于维护植物健康具有十分重要的作用,其影响因素也非常多。
本研究以四川栗子坪国家级自然保护区内峨热竹竹叶为实验材料,通过室内微生物学分析、高通量扩增子测序技术等手段,探讨小相岭山系四川栗子坪国家级自然保护区内野生大熊猫主食竹峨热竹叶围细菌组成、多样性的季节性变化,以期为野生大熊猫食物资源的科学管理提供参考依据。
1 研究方法
1.1 样品收集和处理
在四川栗子坪国家级自然保护区峨热竹分部区域设置采样点,分别于2020年4月至2020年5月(春季)以及2020年10月(秋季)采集峨热竹竹叶,两个季节各采集样品16个。每个竹叶样品不低于200 g,并在4 h内运回保护站存于−20℃冰箱中,并在两天内进行叶围微生物的洗脱处理。
将竹叶样品用TE-buffer(10 mM Tris,1 mM EDTA,pH 7.5)进行洗脱、离心并收集菌泥[18−20],并参照说明书对菌泥用E.Z.N.A.TMSoil DNA Kit(Omega Norcross,GA,U.S.)进行基因组DNA提取。将符合DNA扩增纯度和浓度的样品送往上海美吉生物有限公司进行后续实验。
1.2 16Sr RNA序列的扩增和测序
首先进行DNA的样品质检,在进行DNA纯度检测时采用NanaDrop2000,采用琼脂糖凝胶电泳进行DNA完整性检测。检测结果显示DNA的纯度和DNA的完整度都符合后续扩增的要求。其次进行PCR的扩增,在进行扩增的过程中,为了兼顾扩增特异性,同时避免因测序的序列太长而造成其他有误的影响,选用细菌16SrRNA基因V3-V4高变区进行扩增(引物为338F:ACTCCTACGGGAGGCAGCAG;806R:GGACTACHVGGGTWTCTAAT)[19]。PCR产物经纯化后进行末端补齐,加尾和测序连接头后利用Illumina Hi Seq 2000平台进行测序。
1.3 数据分析
将所得原始测序序列使用Trimmomatic软件质控,使用FLASH软件进行拼接,根据序列首尾两端的barcode和引物区分样品,并调整序列方向,barcode允许的错配数为0,最大引物错配数为2。使用的UPARSE软件(version 7.1 http://drive5.com/uparse/),根据97%的相似度对序列进行OTU聚类,并在聚类的过程中去除单序列和嵌合体。利用RDPclassifier(http://rdp.cme.msu.edu/)对每条序列进行物种分类注释,比对16SrRNA基因数据库。去除可能被洗脱微生物时混入的竹叶组织叶绿体和线粒体污染的序列。
基于OTU百分比计算细菌微生物的组成和相对丰度,并使用秩和检验(wilcoxon rank-sum test)不同季节峨热竹叶围细菌在门水平和属水平上的差异性。利用Mothur软件计算样品的Alpha多样性,包括Sobs和Shannon等指数,使用秩和检验不同季节峨热竹叶围细菌Alpha多样性之间的差异性。通过QIIME(v1.80)计算峨热竹叶围微生物群落的Beta多样性,包括weighted Unifrac和unweighted Unifrac指数,并且使用R语言PCoA统计分析并作图,计算基于OTU水平的不同季节峨热竹叶围细菌weighted UniFrac distances和unweighted UniFrac distances距离下微生物结构的相似性和差异性,并使用ANOSIM分析检验其差异性水平[19,21-23]。
2 结果
2.1 峨热竹叶围细菌组成的季节性变化
去除叶绿体和线粒体序列后,基于97%的相似度对序列进行聚类共获得1796个OTUs,其中,春季和秋季共有934个OUTs,春季独有101个OUTs,秋季独有761个OUTs。将相对丰度百分比小于1%的归类为others后,发现峨热竹叶围主要有变形菌门(Proteobacteria)、酸菌门(Acidobacteriota)、拟杆菌门(Bacteroidota)、放线菌门(Actinobacteriota)和扁平菌门(Planctomycetota)等5个菌门(见图1),其中变形菌门最为丰富。春季峨热竹叶围酸菌门细菌的相对丰度显著大于秋季峨热竹叶围酸菌门细菌相对丰度(见图1,P<0.01),而变形菌门细菌的相对丰度显著小于秋季变形菌门细菌的相对丰度(见图1,P<0.05)。
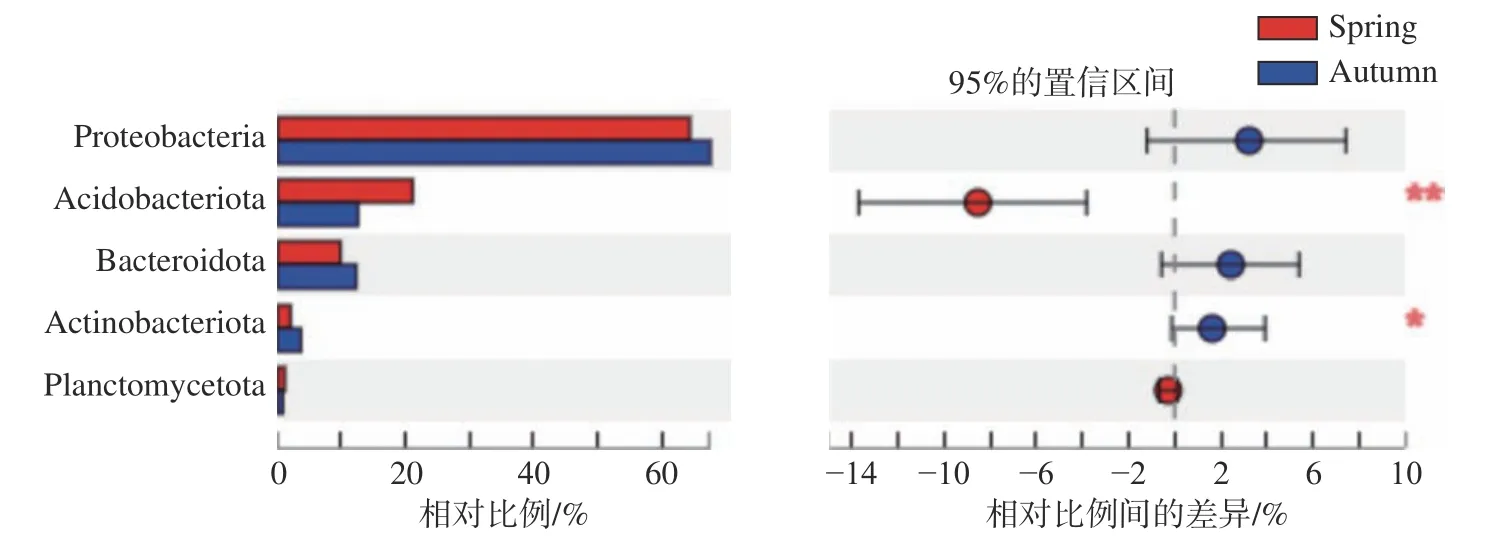
图1 不同季节峨热竹叶围细菌在门水平上的相对丰度差异性比较Fig.1 Community taxonomy of A. spanostachya phyllosphere bacteria on phylum level in different seasons (*<0.05;**<0.01)
在属水平,峨热竹叶围上主要分布有1174-901-12属、鞘氨醇单胞菌属(Sphingomonas)、Granulicella、嗜酸菌属(Acidiphilium)、未被分类的醋杆菌属(unclassified-f-acetobacteraceae)、薄层菌属(Hymenobacter)、Terriglobus、假单胞菌属(Pseudomonas)等属的细菌(见图2)。其中,春季峨热竹叶围的1174-901-12、Granulicella、unclassifiedf-acetobacteraceae、嗜酸菌属(Acidiphilium)、薄层菌属(Hymenobacter)、Endobacter、Bryocella等属细菌的相对丰度显著大于秋季(见图2)。而鞘氨醇单 胞 菌 属(Sphingomonas)、Mucilaginibacter、unclassified-f-Beijerinckiaceae、Massilia、norank-f-Acetobacteraceae、Pedobacter等属细菌则是春季的显著小于秋季(见图2)。
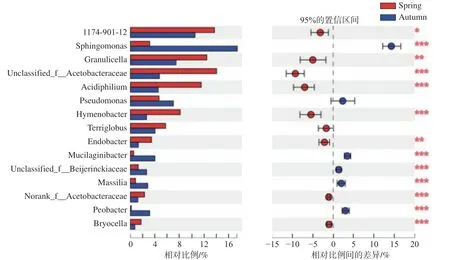
图2 不同季节峨热竹叶围细菌在属水平上的相对丰度及其差异性检验Fig.2 Community taxonomy of A.spanostachya phyllosphere bacteria on genus level in different seasons(*<0.05;**<0.01;***<0.001)
2.2 峨热竹叶围细菌α多样性的季节性变化
α多样性是指通过单样本的多样性分析反映样品内的微生物群落的丰富度和多样性,包括Sobs index、Shannon index等[15,21-22]。Sobs index表示样本中观察到的细菌数目,而Shannon index表示菌群多样性,包括了物种的数目和多度。使用QIIME软件分别对每个样品计算Sobs index和Shannon index。分析发现,峨热竹叶围细菌的Sobsindex和Shannon index均是秋季显著大于春季(见图3)。
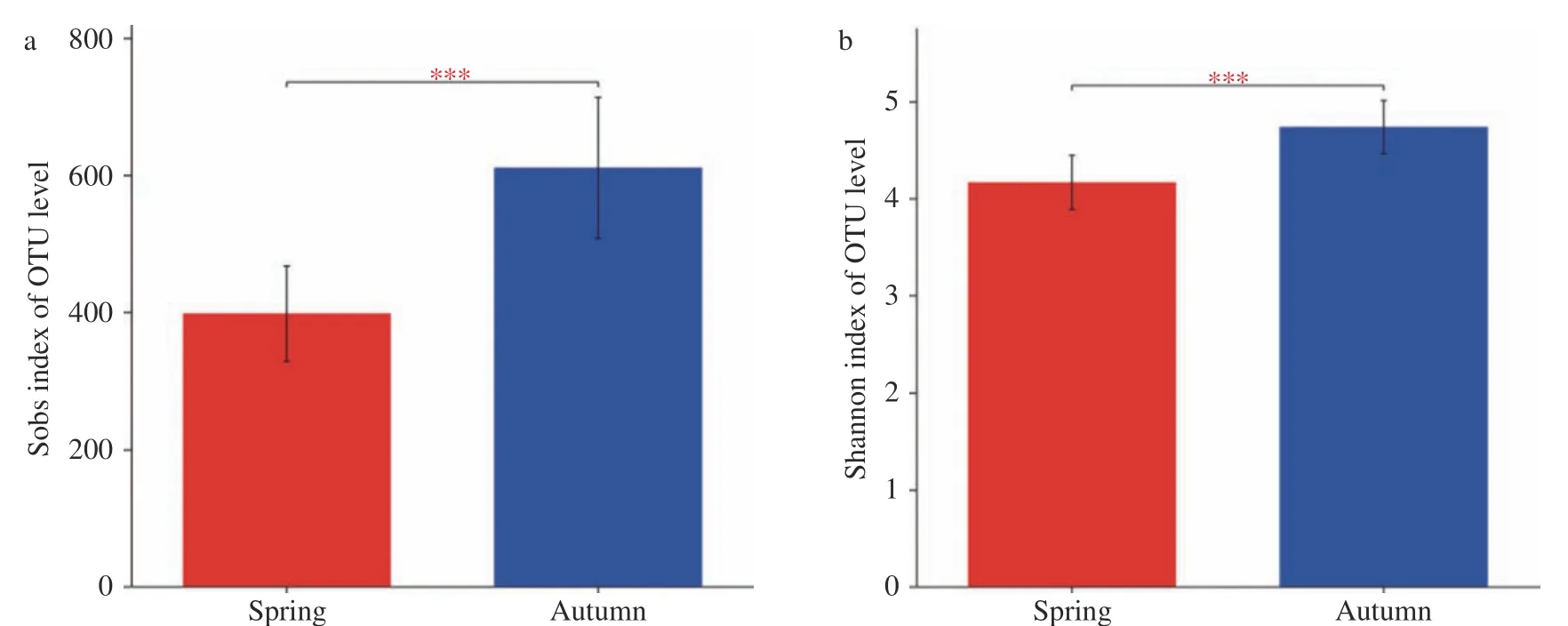
图3 基于OTU水平的不同季节峨热竹叶围细菌α多样性(Sobs index、Shannon index)差异性比较Fig.3 Alpha diversity (a:Sobsindex; b:Shannon index)of A. spanostachya phyllosphere bacteria on OTU level in different seasons(***<0.001)
2.3 峨热竹叶围细菌β多样性的季节性变化
PCoA分析(Principal coordinates analysis),即主坐标分析,是一种非约束性的数据降维分析方法,可用来研究样本群落组成的相似性或差异性,是β多样性分析中常用的一种方法[15,21-22]。基于OTU水平的不同季节峨热竹叶围细菌主坐标分析发现,春季和秋季峨热竹叶围细菌在weighted-UniFrac distance和unweighted-UniFrac distance的群落结构上具有显著差异性(weighted-UniFrac distance,R=0.74,P<0.001;unweighted-UniFrac distance,R=0.78,P<0.001),其第1主成分贡献率分别达到了44.86%和23.33%(见图4)。

图4 基于OTU水平的不同季节峨热竹叶围细菌主坐标分析(PCoA)。Fig.4 Principal coordinates analysis(PCoA analysis)of A. spanostachya phyllosphere bacteria on OTU level in different seasons
3 讨论
本研究中,在春季峨热竹叶围菌群独有101个OUT,占序列总数的5.62%;在秋季独有761个OUT,占序列总数的42.37%。秋季的细菌丰度远高于春季,这可能是温度变化等因素造成的,此外,叶围营养不足等状况也与细菌菌群的多样性有关[9,18]。
同一竹种在不同季节中的优势菌存在差异,本研究发现在所有样品中,除去others共检测到5个菌门,分别是变形菌门(Proteobacteria)、酸菌门(Acidobacteriota)、拟杆菌门(Bacteroidota)、放线菌门(Actinobacteriota)和扁平菌门(Planctomycetota),其中丰度最大的优势菌门是变形菌门(Proteobacteria)。在属水平上,春季和秋季的菌群组成有一些差异,除去others后,春季有19个属,其中占比超过5%的属有1174-901-12属、Granulicella属、未被分类的醋杆菌属(unclassified-f-acetobacteraceae)、嗜酸菌属(Acidiphilium)、薄层菌属(Hymenobacter)、Terriglobus属。优势菌属为1174-901-12属,独有属为Bryocella属和Tundrisphaera属。秋季除去others后还剩20个属,其中占比超过5%的属有1174-901-12属、鞘氨醇单胞菌属(Sphingomonas)、Granulicella属、假单胞菌属(Pseudomonas)。优势菌属是鞘氨醇单胞菌属(Sphingomonas),独有属是土地杆菌属(Pedobacter)。以上结果与前人对竹种叶部微生物的研究结果相符[24];具有降解有机物的鞘氨醇单胞菌(Sphingomonas)和具有植物促进剂的潜能的假单胞杆菌属(Pseudomonas)等植物有益微生物在秋季峨热竹叶围的显著增加,有利于该竹种的健康和生长[10-14]。造成不同季节中优势菌存在差异的原因可能是在不同的生长状态和生理时期下,竹种叶围微生物会有不同的适应性,这会直接导致微生物的种群组成和优势菌在季节上的差异。
有学者在研究不同竹种的营养成分对大熊猫肠道影响时发现,不同情况下大熊猫肠道中均表现为变形菌门(Proteobacteria)、酸菌门(Acidobacteriota)、扁平菌门(Planctomycetes)和厚壁菌门(Firmicutes)占主导,其中变形菌门丰富度最高[25],大量对大熊猫、小熊猫的肠道菌群研究对结果表明,变形菌门(Proteobacteria)、厚壁菌门(Firmicutes)和拟杆菌门(Bacteroidetes)是大熊猫、小熊猫胃肠道的优势门[20,26-28],变形菌门最为丰富的结果与我们的研究结果类似。
从香农指数和Sobs指数的分析结果中可以看出,峨热竹的细菌菌群在春季和秋季之间存在非常明显的差异,并且是秋季中细菌菌群的多样性和丰富度远远高于春季中细菌菌群的多样性和丰富度,这进一步说明,季节性变化导致了细菌菌群出现了显著差异。这与前人对四川省6种竹叶部微生物的研究结果一致,该研究结果也证明竹种微生物类群的丰富度指数在秋季达到最高值[29]。在四川地区春季的气候特征分别表现为低温多雨,这样的特征不利于微生物在叶表面附着,而秋季拥有更为高温潮湿的气候条件,这便使得大多数微生物可以附着在叶表面,并且进行大量繁殖等生命活动,因此叶表面的微生物丰富度指数在秋季高于春季。基于PCoA分析的结果表明峨热竹春季的细菌群落和秋季的细菌群落有着明显的不同,再次说明了细菌菌群在不同季节间具有显著差异[9,18,25]。
保护大熊猫主食竹已经成为保护大熊猫的一种重要手段,四川栗子坪国家级自然保护区中峨热竹又是大熊猫的主要采食竹,因此对峨热竹对科学保护是该山系大熊猫保护对重要内容。本研究表明,峨热竹叶围细菌菌群中变形菌门(Proteobacteria)占优势,该门细菌也是大熊猫肠道微生物中的优势菌群,因此该门细菌优势度的变化可能对该山系大熊猫肠道健康产生潜在影响。另外,对植物有益的鞘氨醇单胞菌属(Sphingomonas)和假单胞杆菌属(Pseudomonas)细菌在秋季显著增加且为主要菌群,然而在春季并不是优势菌群,因此在其他季节(包括春季)检测该类菌群的分布和活性有利于了解不同季节峨热竹叶围有益微生物的状况。以上研究结果有助于了解不同季节小相岭山系峨热竹的叶围细菌菌群的差异性,为大熊猫主食竹资源的保护和科学管理提供参考依据。
猜你喜欢
中国农学通报(2022年14期)2022-06-01
油气田环境保护(2022年2期)2022-05-09
小哥白尼(野生动物)(2021年7期)2021-11-20
山东畜牧兽医(2021年5期)2021-06-07
少儿美术(2020年9期)2020-11-05
幼儿教育·父母孩子版(2020年8期)2020-03-04
娃娃乐园·综合智能(2019年10期)2019-12-07
中国沼气(2019年1期)2019-04-13
陶山(2019年1期)2019-03-29
学苑创造·A版(2018年7期)2018-08-07
