
NGS-metabarcoding revealing novel foraminiferal diversity in the Western Pacific Magellan Seamount sediments*
2021-10-12 05:00JunfengSHIYanliLEIHaotianLITiegangLI
Junfeng SHI ,Yanli LEI ,Haotian LI ,Tiegang LI ,
1 Laboratory of Marine Organism Taxonomy and Phylogeny, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China
2 Key Laboratory of Marine Sedimentology and Environmental Geology, First Institute of Oceanography, Ministry of Natural Resources (MNR), Qingdao 266061, China
3 Clinical Research Center, Affi liated Hospital of Weifang Medical University, Weifang 261031, China
4 Southern Marine Science and Engineering Guangdong Laboratory (Zhuhai), Zhuhai 519082, China
5 Laboratory for Marine Biology and Biotechnology, Pilot National Laboratory for Marine Science and Technology (Qingdao),Qingdao 266237, China
6 University of Chinese Academy of Sciences, Beijing 100049, China
7 Center for Ocean Mega-Science, Chinese Academy of Sciences, Qingdao 266071, China
Abstract Seamount is a unique deep-sea ecosystem widely distributed in the world.Its biodiversity is vibrant due to its specific geographical and hydrological conditions.However,the diversity and features of foraminifera in such an environment have rarely been studied.We extracted environmental DNA (eDNA) in sediments and amplified the partial small subunit ribosomal DNA (SSU rDNA) of foraminifera to understand the foraminiferal diversity from four sites in Magellan Seamount (Western Pacific Ocean).Partial SSU rDNA sequencing was conducted and 912 979 foraminiferal reads were obtained and gathered into 266 operational taxonomic units (OTUs).In the available dataset,a high proportion of rare OTUs and low identity OTUs in each studied sample showed that the Magellan Seamount foraminiferal community might have a high genetic novelty.The relative abundance of foraminifera varied between replicates probably due to the existence of bias in amplification process and patchiness of the deep-sea floor.It showed that the Magellan Seamount has a relatively high benthic foraminiferal diversity characterized mainly by monothalamiids (76.37% of total reads) in association with rotaliids (19.03% of total reads),including planktic foraminiferal sequences(38.58% of rotaliids;7.36% of total reads).The remaining reads were assigned to miliolids (0.83% of total reads) and textulariids (0.66% of total reads),and 3.11% of total reads are unassigned to a specific family.The comparative analysis with foraminiferal assemblages from coastal and deep-sea environments indicated that seamounts could aggregate species from a nearby deep-sea.
Keyword:metabarcoding;Magellan Seamount;foraminifera;Western Pacific Ocean
1 INTRODUCTION
Foraminifera are commonly defined as marine testate protists among the most abundant,diverse,and widely distributed deep-sea microfauna.As significant components of deep-sea biotic communities,foraminifera play pivotal roles in ecological processes and the flow of energy and materials through ecosystems (Cornelius and Gooday,2004;Gooday and Jorissen,2012;Lei et al.,2014).Benthic foraminifera are sensitive to biotic and abiotic factors,and their abundance and test morphology allowed the recovery of paleoecological changes (Mojtahid et al.,2010;Bejaoui et al.,2019).Their assemblages can reveal information about salinity,temperature,food supply,oxygen,and bathymetry (Lei et al.,2017).Planktic foraminifera are considered as good evaluating indicators for open marine environments(Li and Keller,1998;Saïdi and Zaghbib-Turki,2016).Their community structures tend to be affected by abiotic factors and indicate the water hydrography(Vincent and Berger,1981;Zhang,1985;Naidu and Malmgren,1996;Li and Keller,1998;Saïdi and Zaghbib-Turki,2016;Jian et al.,2020).These reconstructions are established from the chemical composition and morphology of foraminiferal fossil test (Li and Keller,1998;Katz et al.,2010).
Furthermore,the most part of living foraminifera ecological studies focused on shallow water environments (Lei et al.,2014,2017).Little is known about the biology and the diversity of living benthic foraminifera at the deep-sea environments as few works have been carried out on them (Lee and Anderson,1991;Lecroq et al.,2011;André et al.,2014,Angeles et al.,2020),especially in the seamounts where research data on foraminifera are very few (Ohkushi and Natori,2001;Shi et al.,2020).In addition,deep-sea sampling is challenging due to the vastness and remoteness of the deep seabed.Insuffi cient sampling limits to obtain a comprehensive assessment of foraminiferal diversity in the deep-sea(Lecroq et al.,2011;Goineau and Gooday,2019).
A seamount corresponds to an uplifted terrain with a height of more than 1 000 m (without reaching the sea surface) in the deep-sea (Epp and Smoot,1989).It is a unique habitat in the deep-sea,which is globally distributed in the oceans (Clark et al.,2010;Yesson et al.,2011).The seamount has a high biodiversity due to its specific geographical and hydrological characteristics (Taylor Column) (Parker and Tunnicliffe,1994;Stocks,2004a,b).All studies on foraminiferal diversity in seamount are based on their morphology (Nienstedt and Arnold,1988;Heinz et al.,2004;Gooday and Jorissen,2012).However,most deep-sea species are tiny,fragile,and challenging to identify (Lecroq et al.,2011).A large spectrum of deep-sea foraminifera belongs to early lineages characterized by simple,single-chambered(monothalamiids),organic-walled,or agglutinated tests and are poorly preserved in the fossil record(Gooday et al.,2001,2004;Lecroq et al.,2011).Thus,it is challenging and even impossible to decipher living foraminiferal diversity in seamounts based only on test morphology.In the past decade,molecular studies have revealed the diversity and geographic distribution of benthic foraminifera in deep-sea sediments (Pawlowski and Holzmann,2007;Lecroq et al.,2011;Lecroq,2014).However,most of these studies relied on clone libraries,limiting the number of sequences available for analysis (Lecroq et al.,2011).Next-generation sequencing (NGS) used for environmental rDNA sequencing reduced clone libraries’ limitations and made the decoding application possible (Lecroq et al.,2011;Frontalini et al.,2020).This approach made it possible to analyze the living foraminiferal diversity that is tiny,fragile,or rare to identify using classic methods in the deepsea comprising seamount habitats (Logares et al.,2014;Egge et al.,2015;Piredda et al.,2018;Shi et al.,2020).
Meanwhile,the total environmental DNA (eDNA)contains living cell and extracellular DNA(Pietramellara et al.,2009),it is characterized by mixing the genomic DNA of different biota present in the studied environment and has enormous potential to address fundamental research questions related to molecular biology,ecology,paleontology,and environmental sciences (Thomsen and Willerslev,2015).This provided an opportunity to study the sequence of foraminifera ex situ.
This study aims to (1) identify the taxonomic composition and diversity of foraminifera in the Magellan Seamount (Western Pacific Ocean)sediments,(2) reveal variations between different replications or sites,and (3) determine if there are foraminiferal sequences from other sources in sediments except in-situ benthic foraminiferal data.
2 MATERIAL AND METHOD
2.1 Location and sample collection
Twelve samples have been collected from four sites (with three sample replicates from each site)from the Magellan Seamount,which lies adjacent to the southeast of the Mariana Trench in the Western Pacific Ocean and comprises two concomitant top flat seamounts (Fig.1).To compare foraminiferal diversity from the four sites,sediments were collected at water depths ranging from 1 308 to 1 968 m using a pushcore operated by a remotely operated vehicle (ROV),Faxian(Discoveryin Chinese),equipped by the R/VKexue(Sciencein Chinese) in April 2018 (Fig.1).The ROV was only launched four times in four chosen sites as it was challenging to obtain the sediment samples from the Magellan Seamount.After collection,sediments were immediately frozen at-80 °C (sample information is presented in Table 1).

Fig.1 Location map of the studied sites of Magellan Seamount in the Western Pacific Ocean
Table 1 Summary of sampling sites,numbers of foraminiferal read and OTUs
2.2 eDNA extraction,amplification,and sequencing
The eDNA was extracted using DNeasy PowerSoil kit (QIAGEN,Germany) for sediment samples.Approximately 0.25 g of the sediment was required for each sample under the extraction process.Afterward,the eDNA extracts were stored at -20 °C.The foraminiferal 37f hypervariable region of SSU rDNA was amplified using a High-Fidelity Master Mix (TRANSGEN,China) by nested polymerase chain reaction (PCR).We used foraminiferal-specific primer pair,s14F3 (5ʹ-ACGCAMGTGTGAAACTTG-3ʹ) and s17 (5ʹ-CGGTCACGTTCGTTGC-3ʹ),and a long fragment (300–500 bp) was amplified in the first reaction.A shorter fragment (210–510 bp) was reamplified using the s14F1 (5ʹ-AAGGGCACCACAAGAACGC-3ʹ) and s17 primer pairs following Lejzerowicz et al.(2010).In the first amplification,34 cycles were programmed at 50 °C for annealing temperature;in the second amplification,the program was set at 24 cycles and 52 °C.The PCR system contained 12.5 μL of two times High-Fidelity Master Mix,0.5 μL of each primer (10 μmol/L),2 μL of eDNA template,and distillation-distillation H2O(ddH2O) was added to reach a final volume of 25 μL.
The PCR products were purified using the Gel Extraction Kit (OMEGA,USA).Sequencing was performed in Novogene Bioinformatics Technology Co.,Ltd.The sequence libraries were generated using the TruSeq®DNA PCR-Free Sample Preparation Kit for Illumina (NEB,USA) following the manufacturer’s recommendations.The libraries’ quality was conducted using the Agilent Bioanalyzer 2100 system.The library was sequenced using an Illumina HiSeq2500 platform,and 250-bp paired-end reads were generated.
2.3 Data analysis methods
Paired-end reads (raw reads) were split into samples based on unique barcodes and merged using FLASH (V1.2.7,http://ccb.jhu.edu/software/FLASH/;Magoč and Salzberg,2011).FastQC (http://www.bioinformatics.babraham.ac.uk/projects/fastqc/),an adopted application in quality control of NGS following Brown et al.(2017),was used to assess this dataset’s read quality.Low-quality reads (quality value of <20) were removed according to the QIIME(V 1.9.1,http://qiime.org/index.html) (Caporaso et al.,2010;Bokulich et al.,2013) after filtering,and singletons were excluded.OTUs that occurred in less than three samples were excluded to avoid bias due to the presence of extracellular DNA.As PCR errors often generated rare sequences,reads occurring in more than three samples and showing a minimal abundance of ten in all samples are obtained.
The filtered reads (clean reads) were generated to operational taxonomic units (OTUs) using the Unoise3(developed by Edgar (2016)) pipeline with an identified threshold of 100%.After the Unoise3 pipeline generation,each OTU—an artificially defined taxonomic unit based on sequence similarity used to distinguish taxonomic groups—represented a unique sequence.OTUs with less than 10 reads occurring in less than three samples were excluded from the analysis.Representative sequences were extracted from the OTU table and taxonomically assigned based on the PR2 database (Protist Ribosomal Reference Database;Guillou et al.,2012) using BLAST (Version 2.7.1);non-foraminifera (Rank 3 in PR2 database)OTUs were removed.OTUs were defined as abundant when the number of sequences exceeds 1% of all sequences in a sample.Rare OTUs were defined when the number of sequences was less than 0.1% of all sequences in a sample,as was the case for Pedrós-Alió(2006) and Fuhrman (2009).α-diversity (withinhabitat diversity) can reflect the richness and evenness of species within habitat.Thus,the Simpson,Shannon,and Margalef α-diversity indices were calculated in this work using QIIME v.1.91.
3 RESULT
3.1 Overview of NGS dataset
The foraminiferal SSU rDNA dataset contained 1 059 636 raw reads.Based on the Illumina sequencing platform,an average of 98.20% reads in the analyzed samples had a high quality score (≥30).The preserved reads were compared with the PR2 database,and seven unique sequences with 1 516 reads were assigned to Excavata (amitochondriate,Protozoa without mitochondria,Rank 2 in PR2 database).These non-foraminiferal sequences were discarded from the dataset.After quality filtering,912 979 foraminiferal reads were retained and the percentage of filtered reads varied from 4.25% (FXD174.1) to 22.20% (FXD175.3).
The rarefaction curves (Fig.2a) show that all samples were fully sequenced as they tended to saturation.In this study,the numbers of effective reads in all samples were more than 60 000 (see Reads after filtered;Table 1).The reads were clustered into 266 distinct OTUs based on 100% sequence similarity.To avoid the threshold affecting clustering accuracy and achieve the best phenotype resolution,we defined each sequence as a separate OTU.The OTUs for each sample ranged from 107 to 176,with an average of 139.The details of the samples and foraminiferal reads used in our analysis are shown in Table 1.
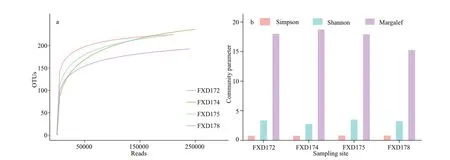
Fig.2 Comparison of α-diversity from the four studied sites
3.2 Taxonomic composition
The taxonomic assignment of the obtained 266 OTUs revealed 38 genera and 45 species among planktic and benthic foraminifers.The OTUs were assigned to a reference database with low similarity in most cases,and only 91 OTUs (34.21%,corresponding to 20.94% of the reads) reached a similarity of more than 97% (identity-value >0.97,see Supplementary Table S1) with targeted sequences in the PR2 database.The average identity-value of all detected OTUs was 93.59.The Sankey diagram (Fig.3a) showed the flow of NGS reads (left) and OTUs (right) from the four studied locations,which are assigned to different foraminiferal families (Rank 5 in PR2 database).Approximately,half of the OTUs (119 out of 266)were with less than 1 000 reads (Fig.3b &c).An OTUs with maximum numbers of reads were assigned toToxisarconsp.(74 346 reads),Elphidium frigidum(33 875 reads),Siphonapertasp.(7 524 reads),andTrochammia hadai(2 318 reads),becoming the most abundant species of monothalamiids,rotaliids,miliolids,and textulariids,respectively.The 15 most abundant OTUs approximately reached half of the reads.Only 13 OTUs occurred in all the studied samples.
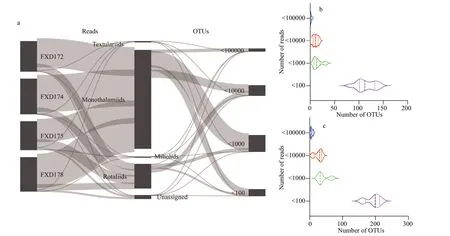
Fig.3 The relationship between OTUs and reads
Figure 4a shows the distribution of foraminifera in the analyzed samples,including all reads in sediments.Most OTUs were assigned to (Rank 5 in PR2 database)monothalamiids (single-chambered and soft-walled)(191 out of 266),and 76.37% of the OTU reads were distributed between the samples under study (697 226 out of 912 979).These monothalamiids correspond to more than 70% of the reads from eight samples(Fig.4b).Six OTUs with 28 392 reads were unassigned to a definite family (Rank 5 named foraminifera_XX;Supplementary Table S1),which represented 3.11%of all reads.The remaining 69 OTUs were assigned to rotaliids (58 OTUs),miliolids (2 OTUs),and textulariids (9 OTUs),representing 19.03%,0.83%,and 0.66% of the foraminiferal reads (including planktic and benthic foraminifera),respectively.
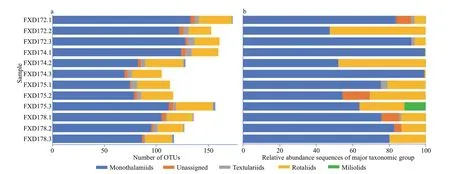
Fig.4 Relative abundances of the number of OTUs (a) and sequences (b) related to the major families of foraminifera detected by DNA sequencing in different sites
With calcareous tests,planktic foraminifera can be placed within the radiation of the rotaliids (Ujiié et al.,2008).In this dataset,23 OTUs (out of 266) were assigned to planktic foraminifera (globigerinacea,Rank 6 in PR2 database),representing 67 239 reads(7.36% of the total dataset;Supplementary Table S1).Planktic foraminiferal sequences were annotated to six species:Globigerinitauvula,Globigerinitaglutinata,Globigerinabulloides,Candeinanitida,Pulleniatinaobliquiloculata,andGlobigerinellasiphonifera.Among these species,Globigerinitauvulahad 14 OTUs and the most number of reads (53 346),representing approximately 79.34% of the planktic foraminiferal sequences.The number of OTUs ranged 14 (in FXD178)–23 (in FXD175),representing 7.25% (in FXD178)–10.31% (in FXD175) of each site’s total OTUs (Fig.5a).In the four sites,4 296 (in FXD172)–27 132 (in FXD175) planktic foraminiferal reads were retained,representing 1.99% (in FXD172)–13.32% (in FXD175) of the total sample sequence(Fig.5b).The absolute and relative abundance of planktic reads varied from site to site,but the number and proportion of OTUs were stable (Fig.5).
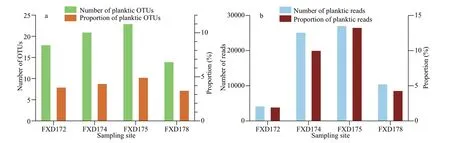
Fig.5 Occurrences of planktic foraminiferal eDNA in the abyssal sediments
3.3 Comparative analyses
The benthic foraminiferal diversity variation in sediments was observed among different sites and replicates.The number of OTUs among sites was the highest in FXD172 (on average of 164.7) and the lowest in FXD178 (on average of 128.7;Table 1).The rarefaction curves (Fig.2a) show that FXD174 has the highest value (Margalef value) and number of observed OTUs but the lowest Shannon-Wiener value.Furthermore,the values of the Margalef and Simpson indices have no remarkable changes among the four stations.The number of shared OTUs in all sites reached 131.Only 26.6% of all OTUs occurred in three replicates.Besides,the proportion of these OTUs in each site was 39.1% (FXD172),16.5%(FXD174),18.8% (FXD175),and 40% (FXD178)(Fig.6a–d).The average percentage of the shared OTUs in a site was about 32.8% (46.2% in FXD172,18.6% in FXD174,22.4% in FXD175,and 44% in FXD178).The number of exclusive OTUs in FXD174.3 and FXD175.3 exceeded the shared ones in all samples of one site (Fig.6b–c).To mine the dataset more deeply,we divided OTUs into abundant and rare.Moreover,statistical work was conducted separately in two families (Table 1).The number of abundant and rare OTUs varied between sites,whereas the difference between replicates among each site seems to be more significant.
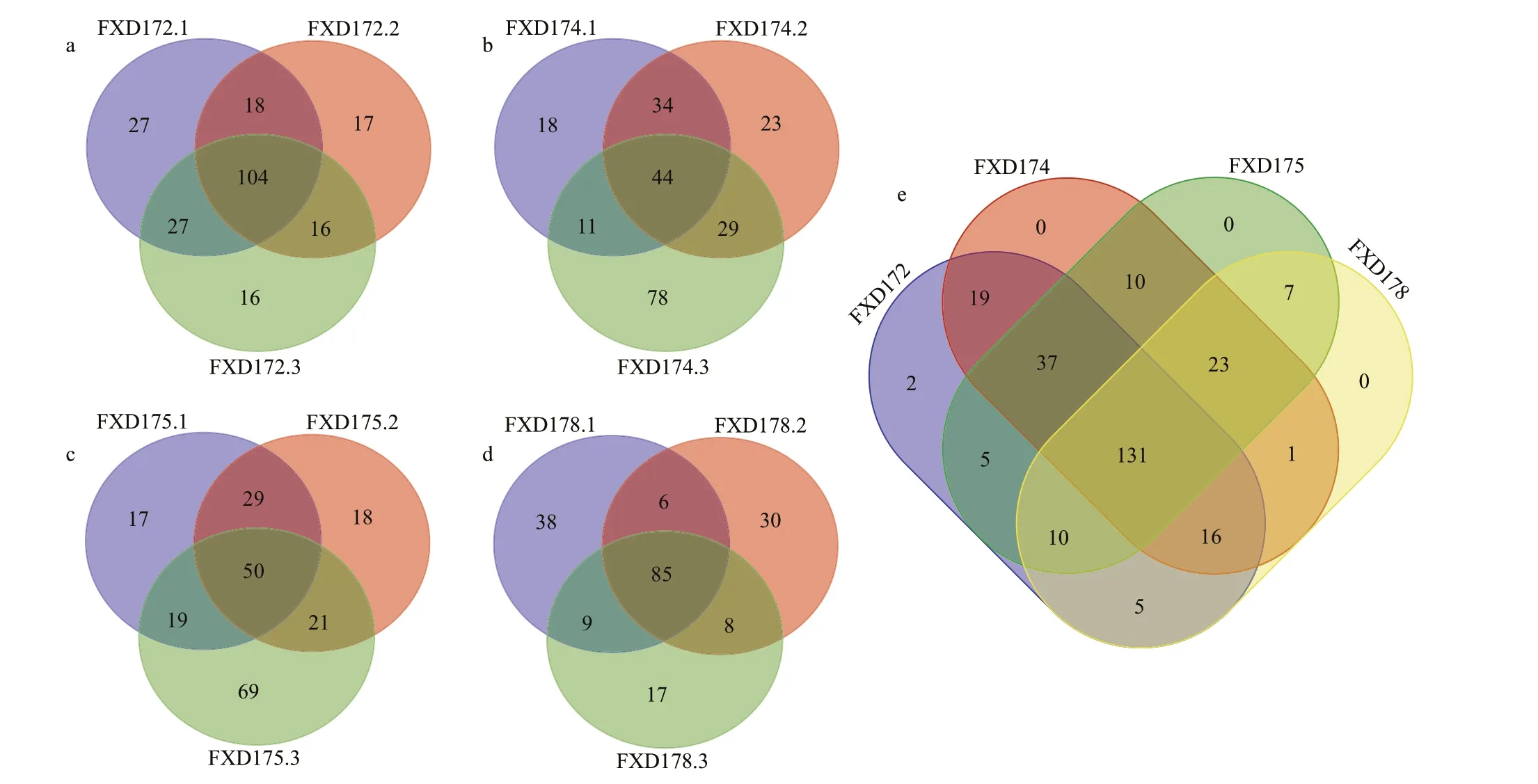
Fig.6 The number of unique and shared OTUs in different replications and sites
4 DISCUSSION
4.1 Deficiency of metabarcoding in foraminiferal research
In theory,the more sequences in a dataset,the better it reflects the sample’s actual composition.However,superfluous sequences meant a greater investment in the sequencing process,so we had to compromise between the sequencing data amount and sequencing costs.Approximately 20 000 effective reads per sample seemed suffi cient for a diversity study (Kozich et al.,2013),covering more than 95%of the OTUs (Lecroq et al.,2011).There is no rationale for defining the optimal size of environmental sequence datasets (Ni et al.,2013;Lejzerowicz et al.,2014).In this dataset,all the studied samples have more than 60 000 effective reads,which means the data are suffi cient for ecological research.
The PCR amplification process is biased but it influences the estimation of species relative abundances (Acinas et al.,2005;Clarke et al.,2014;Pawluczyk et al.,2015).Predictions of the species relative abundances often depend on the number of sequences.However,equal amounts of template DNA from different species did not yield equal proportions of amplicons (Elbrecht and Leese,2015;present study).The proportion of different families (including planktic and benthic foraminifera) among samples was similar regarding the number of OTUs (Fig.4a).In contrast,the proportion of different families among samples had extremely high variations regarding the relative abundance of sequences (Fig.4).This indicates that the relative abundance of the sequence was inaccurate for biomass evaluation.The effi ciency’s inequality can explain the result in amplifying multi-species mixed templates using the universal primer (Polz and Cavanaugh,2008;Piñol et al.,2015).
4.2 High genetic novelty of foraminifera in Magellan Seamount
The low identity-values of OTUs in our dataset may show a high genetic novelty as it was reported in other seamounts (Shi et al.,2020).The application of NGS contributed substantially to the detection of broader protist diversity.However,related research is also facing some challenges (Pawlowski et al.,2014).Many researchers have used a 97% sequence identity threshold to distinguish different species in the previous works (Ciardo et al.,2006;Westcott and Schloss,2017).This threshold was first proposed by Stackebrandt and Goebel (1994) when 16S rDNA sequences were considered suitable to estimate the species composition of a community.However,Edgar(2018) proved that the optimal identity threshold of a high-quality set of 16S rRNA sequences from known species was higher than 97%,and the optimal threshold was data-dependent.Previous studies have indicated that foraminifera have a high rDNA variation,leading to overestimated molecular diversity in environmental samples.Meanwhile,the database’s lack of reference sequences may also cause the pseudomorph of high genetic novelty.The extremely high genetic novelty recorded in our dataset may be a pseudomorph generated due to the foregoing circumstances or the embodiment of high biodiversity in the seamount area.
4.3 Variations of benthic foraminifera among samples and replicates
Variations of benthic foraminiferal relative abundances occurred in the four studied sites but were the most evident in the site FXD178 (Fig.4).The water depth in the four stations is approximately about 1 300–2 000 m and cannot be regarded as a controlling factor of these abundances’ changes.However,our results infer that heterogeneity of surface sediments can be responsible for the recorded variations in benthic foraminiferal relative abundances(Frontalini et al.,2020;Shi et al.,2020).FXD178 was in a relatively different habitat compared with other stations,and these habitat variations led to even moresignificant differences in diversity than that of water depths.
Not all significant families appeared in the three replications (related to the four sampled sites).For example,miliolids were absent in FXD172.2 and FXD172.3 but have been recorded in FXD172.1(Fig.4a).With a considerable number of rare OTUs(Table 1) in each sample,heterogeneity occurred in almost every replication.For example,FXD174.3 and FXD175.3 have more exclusive OTUs than the other two replications.This heterogeneity among replications can be explained by the surface sediments“patchiness”,which was characterized by vertical and horizontal distribution variations of microfauna in microhabitats even occurred on the centimeter scale and was widespread in the deep-sea (Jorissen et al.,2007;Lejzerowicz et al.,2014;Bejaoui et al.,2019).The heterogeneity of deep-sea microhabitats affected the diversity of replications,and this phenomenon appeared in the study of nematodes (Danovaro et al.,2013),ciliate (Zhao et al.,2017),and foraminifera(Lejzerowicz et al.,2014).
4.4 Planktic foraminiferal assemblages in sediments
In accordance with Morard et al.(2017) and Shi et al.(2020),we infer that the DNA of planktic foraminifera was preserved in the sediments.For example,the planktic foraminiferal reads accounted for 1.94% of the total reads in the seamount (Shi et al.,2020).An organism’s DNA can be adsorbed to solid particles (Herndl and Reinthaler,2013),such as marine snow,fecal pellets,and massive falls of phytoplankton material after surface blooms (Torti et al.,2015).The solid particles may agglomerate particles and enable the absorbed DNA to sink rapidly and become a record of extracellular DNA (Corinaldesi et al.,2011;Torti et al.,2015;Morard et al.,2017).The extracellular DNA could reconstruct planktic eukaryotic groups (Capo et al.,2015) and become a significant archive of present eukaryotic gene sequences (Corinaldesi et al.,2011).Planktic foraminiferal sequences in sediments provide an opportunity to understand the upper water column’s sedimentation process.Although the number of planktic sequences fluctuated considerably between different sites or samples,the number of OTUs kept the same order of magnitude.This enables to reconstruct species composition in the water column and determine the traceability of planktic sequences in sediments (Angeles et al.,2020).According to this track,we can determine the dynamic sedimentation pattern of plankton undercurrent.This will provide a basis for using sediment data to reconstruct the water environment column.
4.5 Characteristics of foraminiferal communities
The dominance of monothalamiids is consistent with previously published results based on morphology (Gooday,2002;Gooday et al.,2004) and molecular biology (Lecroq et al.,2011;Lejzerowicz et al.,2014;Shi et al.,2020).Some classifications inferred from eDNA data (i.e.,monothalamiidsclade-E in Supplementary Table S1) are not the most abundant family in our dataset,different from results of Lejzerowicz et al.(2014).The composition of foraminiferal assemblages in the seamounts had significant proximity to the ordinary deep-sea and coastal area characterized by abundant monothalamiids and rotaliids taxa as recorded by Lecroq et al.(2011),Pawlowski et al.(2011),and Li et al.(2020).
As for the diversity of foraminifera,Magellan Seamount had a relatively high Shannon-Wiener index (>3;Fig.2b) compared with adjacent abyssal plains (<2.6;Shi et al.,2020).In the typical deepsea,the Shannon-Wiener index of foraminifera(including planktic and benthic) exhibited a lower value (approximately 2;Corliss et al.,2009).In the coastal Jiaozhou Bay,the Shannon-Wiener index of foraminifera was higher (superior to 5;Li et al.,2020)than the Shannon-Wiener index recorded in the studied Magellan Seamount.From the coastal shallow water area to the ordinary deep-sea and then to the seamounts,the diversity of foraminifera first declined and then rose.Nevertheless,the composition of foraminiferal assemblages did not change and still be dominated by monothalamiids and rotaliids.The foraminiferal communities in the seamounts seem more similar to the distant coastal area than the nearby deep-sea and seamount sections.Furthermore,the specific hydrologic conditions (Taylor Column)(Parker and Tunnicliffe,1994) of the seamounts favored species gathering.Thus,we speculate that the foraminiferal community in the deep-sea was from the foraminiferal species diffused from the coastal area.The foraminiferal planktic larvae from the deepsea aggregated around the seamounts,which caused the high foraminiferal diversity in seamounts and similar species composition to the deep-sea and coastal areas.
5 CONCLUSION
Magellan Seamount may have a high genetic novelty due to the high proportion of rare and low identity OTUs.The relative abundance of foraminiferal families varied between replicates.The planktic foraminiferal sequences were detected in the sedimentary environment DNA dataset.The monothalamiids and rotaliids were the prominent foraminiferal families in the Magellan Seamount.
After a comparative analysis of the foraminiferal communities in the coastal area and deep-sea,we speculate that seamounts could aggregate species.
6 DATA AVAILABILITY STATEMENT
Sequence data that support the findings of this study were submitted to the Sequence Read Archive under accession number PRJNA675069.
7 ACKNOWLEDGMENT
The data and samples were collected onboard R/VKexue.We are also very grateful to Dr.Ezzedine SAÏDI for his useful comments and suggestions.
 Journal of Oceanology and Limnology2021年5期
Journal of Oceanology and Limnology2021年5期
- Journal of Oceanology and Limnology的其它文章
- Screening of stable internal reference genes by quantitative real-time PCR in humpback grouper Cromileptes altivelis*
- Morphology and multifractal features of a guyot in specific topographic vicinity in the Caroline Ridge,West Pacific*
- Geochemical characteristics and geological implication of ferromanganese crust from CM6 Seamount of the Caroline Ridge in the Western Pacific*
- Deep-sea coral evidence for dissolved mercury evolution in the deep North Pacific Ocean over the last 700 years*
- Physical oceanography of the Caroline M4 seamount in the tropical Western Pacific Ocean in summer 2017*
- Characteristics and biogeochemical effects of oxygen minimum zones in typical seamount areas,Tropical Western Pacific*