
刺鱼骨盆退化的数学建模研究*
2021-10-10 09:39:10扬州大学胡澳菲嵇佳城
区域治理 2021年36期
扬州大学 胡澳菲,嵇佳城
一、引言
刺鱼类属于硬骨鱼纲,主要生活在北半球的寒带到温带,常见于亚欧大陆和北美山脉的西部和北部,淡水和海水中均有它们的存在。刺鱼体型较小,长约15公分,其重要特征是背部在背鳍前方有一行刺,腹鳍各具有一锐刺。然而,淡水刺鱼在骨盆形态结构上表现出了惊人的差异性。刺鱼骨盆带是在躯干上腹侧和后侧的一对鳍,早期研究表明它们在游泳中起到的作用很有限,但最近的研究表明它们有助于保持平衡。近期研究发现,大约8%的现存刺鱼已经完全失去骨盆,且在不同栖息地间,刺鱼骨盆的缺失程度不完全一致。
研究发现,刺鱼的骨盆退化主要是由Pitx1基因缺失突变、捕食者的缺失及大型水生生物捕食的增加、钙磷等离子浓度的限制三个方面引起。例如,幼年刺鱼更易受到大型无脊椎动物的捕食,这是由于幼年刺鱼骨盆发育尚未完善,骨盆对于捕食者的抵御作用还不够。因此,骨盆的缺失更易受到捕食者的侵蚀,而当缺少相应捕食者时,刺鱼的防御能力就会降低,这也更易引起骨盆的退化。此外,研究还发现刺鱼骨盆的完整度与钙离子浓度呈正相关,钙离子浓度越高,刺鱼骨盆越完整。
本文重点研究捕食者的捕食强度与刺鱼骨盆分布之间的关系,理清捕食者的存在究竟是如何影响刺鱼的骨盆退化。为此,本文构建了基于基因遗传的捕食者与被捕食者种群模型,通过将模拟数据与挪威22个湖泊中刺鱼骨盆分布数据进行对比,探索刺鱼骨盆退化的生物机理。
二、材料与方法
(一)数据约集
通过约集挪威22个湖泊中刺鱼种群密度以及骨盆信息,将骨盆表型分为三类:(Ⅰ)骨盆缺失:骨盆所有成分都缺失;(Ⅱ)骨盆残留:部分骨盆的成分缺失;(Ⅲ)骨盆完整:所有骨盆的成分都完整存在(即前突、升支、后突、骨盆脊柱)。进一步根据骨盆完整度,将其量化为0到8分:其中0分表示完全丧失骨盆,8分表示具有完整骨盆,1至7分表示骨盆不同的退化程度,其中得分越低,退化程度越明显(图1)。

图1 刺鱼骨盆性状得分图
(二)捕食与被捕食者种群模型
Klepaker等人对刺鱼骨盆退化的研究发现,骨盆缩小常由Pitx1基因的缺失突变引起,这在一定程度上说明了刺鱼骨盆退化与基因之间有着密不可分的联系,故本文从基因角度构建模型。一个基因在细胞有丝分裂时有两个基因位点,称为等位基因,分别来自父和母。我们假设由8个值组成的性状差异取决于两个等位基因上可以填充的8个基因位点,令每个基因位点取值为0或1。如果8个基因位点都填满了1,那么结果将是8个1的和,并将导致表型“8”,即代表此种刺鱼骨盆完整。因为等位基因上的基因位点取值只可能是0或1,所以得分情况共有28种可能组合,即共有256种可能的基因组合,但只有9种得分可能性,即0分到8分之间的可能性。

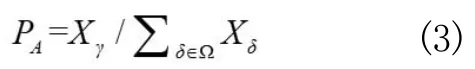
其中,Xã表示其中之一的刺鱼种群密度,表示所有刺鱼种群密度之和,而基因重组产生的概率PR计算公式为:
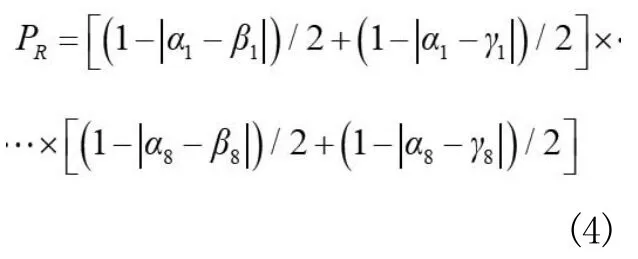
参数选取及数据对比如下:
(1)对于出生率,考虑到刺鱼的骨盆越多,其所需用来生产下一代的能量等也越多,故假设随着骨盆的退化,刺鱼的出生率也会随之增加,因此;(2)对于捕食率,考虑到捕食者对于骨盆越完整的刺鱼捕食难度越大,故假设随着刺鱼骨盆的退化,捕食者对于刺鱼的捕食率也会随之增加,因此;(3)对于死亡率,骨盆越完整,所需提供给骨盆生长的能量与养分就越多,故而其供给自己生存的养分会减少,进而增高了死亡率,因此。在确定了参数表达式的基础上,通过循环取值的方式,在保证不取到负值的情况下,将刺鱼的出生率及被捕食率从0.1到1的各个情况都取遍,得到了共计720种取值情况。
为与挪威湖泊中刺鱼骨盆分布对比,本文引入以下公式来比较差异性,即:

其中fi,i= 1,2,…, 8表示模型所得刺鱼在第i组参数下刺鱼各个性状所对应的种群密度,gi,i=1,2,…,8表示挪威实地考察所得刺鱼在第i个湖泊中各个性状所对应的种群密度。
三、结果与分析
在所给的不同参数组合下,本文模拟刺鱼种群密度分布,并与挪威22个湖泊所对应的种群密度分布对比,计算其密度分布差异性。最后从中挑选出了差异性最小的22组数据,同时展示了具有代表性的三组具有不同骨盆性状的刺鱼密度分布图(图2)。
图2显示,在模型计算并挑选出的22组刺鱼种群密度分布情况中,有17组能够和与之对应的挪威实地湖泊中刺鱼的种群密度分布情况相近,差异性皆保持在0.15以下,甚至有几组能够实现高度一致。虽然无从知晓挪威实地湖泊中各个性状刺鱼的出生率、死亡率和被捕食率,但由结果与挪威实地湖泊中刺鱼的密度分布情况的匹配程度非常高可知,模型计算中所用到的各个性状刺鱼的出生率、死亡率和被捕食率间接反映了与之对应的挪威当地该湖泊的生态环境,尤其是湖泊1-17中捕食者的捕食强度是导致刺鱼骨盆退化的重要因素。对于剩下5组刺鱼种群密度分布情况,其与挪威实地湖泊中的数据的差异性较大。原因有可能是除了捕食者的因素外,还应该存在着其他的因素影响刺鱼的骨盆退化。

图2 22个湖泊密度分布差异性及三组刺鱼密度分布图
四、讨论
本文通过建立捕食者与被捕食者种群模型来探索影响刺鱼骨盆退化的因素。从模型结果来看,大部分结果与挪威实地数据有很高的匹配度。下面以湖泊1,11,19为例,重点讨论这三个湖泊中的具体情况。
对于湖泊1,几乎只有表现型得分为8的刺鱼存活,究其原因,该湖泊中该表现型刺鱼的被捕食率很低,而其他基因型的刺鱼因为受到捕食者的攻击,无法存活。对于湖泊11,发现虽然每种表现型的刺鱼的出生率都较低,但各表现型刺鱼基本都能存活下来,究其原因,该湖泊中各表现型刺鱼的被捕食率相较于其他湖泊相对较小,受到捕食者的攻击较少,这也正是该湖泊中刺鱼能够存活下去的原因之一。对于湖泊19,由于该湖泊中各个基因型的刺鱼被捕食率都很低,而出生率除了表现型得分为0和1的刺鱼,都远远高于被捕食率,这也就是该条湖泊中除了表现型得分为0和1的刺鱼,其他基因型的刺鱼都能存活下去的原因之一。但整体来看,该湖泊的匹配度并不理想,因此可以推测,该湖泊中还存在着其他因素共同影响刺鱼的骨盆退化。
五、结语
本文通过构建基于基因遗传的捕食者与被捕食者种群模型,研究了影响刺鱼骨盆退化的潜在因素,重点研究捕食者的捕食强度与刺鱼骨盆分布之间的关系。从模型所得结果与挪威实地数据的差异性对比来看,该模型下的结果与实地数据做到了大部分相类似,这也就验证了从基因角度入手是切实可行的。同时,通过匹配程度较好的17组数据,反推出了与之对应的湖泊中刺鱼的生态结构,分析得到湖泊中捕食者的捕食强度是导致刺鱼骨盆退化的重要因素,这也使得本文的模型具有了一定的现实意义。对于部分匹配程度不是很好的结果,说明除了捕食者的因素外,还应该存在着其他的因素影响刺鱼的骨盆退化。因此,本文虽已找到并证实了影响刺鱼种群分布的部分因素,但还有许多因素需要进一步的探索和研究。
猜你喜欢
农村百事通(2023年7期)2023-08-05 09:18:56
云南大学学报(自然科学版)(2021年1期)2021-02-05 08:04:04
科学养鱼(2020年8期)2020-12-20 14:29:50
当代水产(2019年8期)2019-10-12 08:57:48
新课程·下旬(2018年9期)2018-11-14 12:49:12
太原师范学院学报(自然科学版)(2018年2期)2018-08-17 12:21:50
东华大学学报(自然科学版)(2018年1期)2018-06-29 03:35:24
渔业致富指南(2016年12期)2016-11-11 07:07:22
中外文摘(2016年13期)2016-08-29 08:53:27
中学生理科应试(2016年7期)2016-05-14 15:39:39
