
IgM分泌细胞在不同年龄家兔胃肠道黏膜中的分布
2021-10-09 09:35刘晓霞曹婷婷李沛轩张睿刘丽萍王雪艳王雯慧
畜牧与兽医 2021年10期
刘晓霞,曹婷婷,李沛轩,张睿,刘丽萍,王雪艳,王雯慧
(甘肃农业大学动物医学院,甘肃 兰州 730070)
黏膜不仅是机体内外环境直接交流的场所,也是机体抵御外界病原微生物的主要屏障。机体绝大部分的感染都是经黏膜入侵引起的,而胃肠道黏膜被认为是机体内最大的免疫器官,每天接触大量的外来抗原,要想识别并清除这些外来成分,就需要黏膜免疫系统发挥相关功能,维持胃肠道的动态平衡。其中,IgM作为黏膜抗体对胃肠道黏膜免疫有着重要作用,它是抵御外源性抗原的第一道防线。
IgM是唯一一种所有脊椎动物中都存在的抗体类别[1],由浆细胞产生并释放,主要以分泌型IgM(sIgM)的形式混入黏膜分泌物中,发挥免疫学作用。sIgM有多种免疫效应功能[2-4],与IgA和IgG相比,能有效地激活补体,发挥补体依赖的细胞毒作用(CDC),同时释放细胞因子、趋化因子或炎症介质,通过与其Fc受体(Fcα/μR和FcμR)的相互作用,参与内吞、介导抗感染免疫。IgM在IgA缺乏时也可以代替其成为黏膜免疫系统的主导性抗体,研究表明,在患有选择性IgA缺陷的人群中,约有一半的人没有症状,就是因为胃肠道黏膜中SIgM的释放增加了[5]。此外,IgM在抗癌免疫反应中也有一定作用,能够有效抑制癌瘤细胞生长,同时杀伤癌瘤细胞,如天然IgM能将捕获的病毒颗粒运送到次级淋巴器官,从而加强获得性免疫应答的速度和强度[6]。
IgM类抗体也可以作为一种诊断工具,用来判断机体是否存在近期感染,对感染监测以及高发地区流行病学调查具有重要意义。例如在这次波及全球的新冠疫情中,就将病毒特异性IgM抗体和IgG抗体阳性作为确诊标准之一,尤其是通过化学发光法检测IgM适用于新型冠状病毒(SARS-CoV-2)检测的各个阶段[7]。另外,高滴度抗体血浆也被应用于治疗重症SARS-CoV-2患者[8]。IgM抗体已经在多个方面发挥出了自己独特的作用,受到了越来越多的人重视。
目前在动物上针对IgM的研究集中于外源性物质(饲料、药物、细菌等)以及各种疾病对其表达和分布的影响,例如,杨红洋等[9]对鸡免疫新城疫疫苗后,探究中药复方多糖对鸡血清中IgM含量的影响;王晓珊[10]研究中药复方“紫黄散”对新城疫病毒(NDV)感染鸡后免疫球蛋白含量及黏膜免疫组织淋巴细胞数量的改变。而关于正常动物IgM分泌细胞的研究相对较少,尤其针对家兔胃肠道的更为少见。本试验以家兔为实验动物,研究IgM分泌细胞在不同年龄家兔胃肠道黏膜的分布,以期为防治家兔胃肠道疾病及新型口服疫苗的开发等提供一些免疫学理论基础。
1 材料与方法
1.1 实验动物
30只日本大耳兔购于兰州兽医研究所实验动物中心,将30只日本大耳兔分为3组,即幼年组(2月龄)、青年组(5~7月龄)、老年组(36月龄),每组10只,雌雄各半。
1.2 主要试剂和仪器
山羊抗兔IgM多克隆抗体(一抗),购于Abcam公司;生物素标记驴抗山羊IgG多克隆抗体(二抗),购于北京博奥森生物技术有限公司;浓缩型DAB试剂盒,购于北京中杉金桥生物技术有限公司;切片机,购于德国Leica公司;DP-71显微照相系统,购于日本Olympus公司。
1.3 样品的采集和制备
1.3.1 组织样品制备
耳缘静注戊巴比妥钠(0.1 mL/kg)致死家兔,打开腹腔采集胃底腺区、幽门腺区、十二指肠、空肠、回肠、盲肠、结肠、直肠等部位组织样品,并立即用10%中性福尔马林溶液固定样品。
1.3.2 切片制作
将固定好的样品按部位取材,胃组织样品大小约为1 cm×1 cm×0.4 cm,肠道组织样品长约为0.5 cm,流水过夜后将组织块分别置于75%、80%、95%、100%梯度酒精中脱水,按常规方法制作石蜡切片,保存备用。
1.4 免疫组化染色
采用间接ABC免疫组织化学染色法。脱蜡至水洗;3% H2O2溶液室温作用15 min,纯水洗2 min×3次;37℃,0.1%胰蛋白酶作用40 min,纯水洗2 min×3次;37 ℃,5% BSA封闭40 min;滴加一抗(1∶500)4 ℃孵育18 h(不超过24 h),PBS洗5 min×4次;滴加二抗(1∶1 000)37 ℃孵育40 min,PBS洗5 min×4次;室温避光进行DAB显色;苏木精轻度复染。之后进行脱水,透明,封片,光学显微镜观察。阴性对照组除用PBS代替一抗孵育外,其余步骤与试验组一致。
1.5 组织学观察及数据处理
每份样品切片随机选择5张,使用Olympus DP-71显微镜观察切片,每张切片随机选择5个视野,统计阳性IgM分泌细胞数量,细胞数量以平均每高倍视野细胞个数(个/HPF)来表示。使用IBM SPSS 22.0软件进行分析统计,结果表示为“平均值±标准差”,并分别进行组内比较(单因素方差分析)和组间比较(独立性t检验),采用ORIGINPRO2018绘图。
2 结果与分析
2.1 胃中IgM分泌细胞的分布特点
经观察发现,未加一抗的胃底腺区(图1A)和幽门腺区(1D)染色阴性;呈阳性的IgM分泌细胞大多为圆形或椭圆形,细胞核呈圆形,位于细胞的一端,有的则在细胞中央,少数核有凹陷,且核不着色,阳性细胞为明显的胞浆染色,根据细胞形态可初步判断IgM分泌细胞为浆细胞。IgM分泌细胞在青年组家兔胃底腺区和幽门腺区的分布略有不同,在胃底腺区中散在分布于胃黏膜固有层(图1B和1C),而在幽门腺区中则主要集中在腺体周围的固有层,少数存在于胃小凹周围的黏膜固有层(图1E和1F)。幼年组和老年组家兔胃中IgM分泌细胞的分布规律与青年组相似。
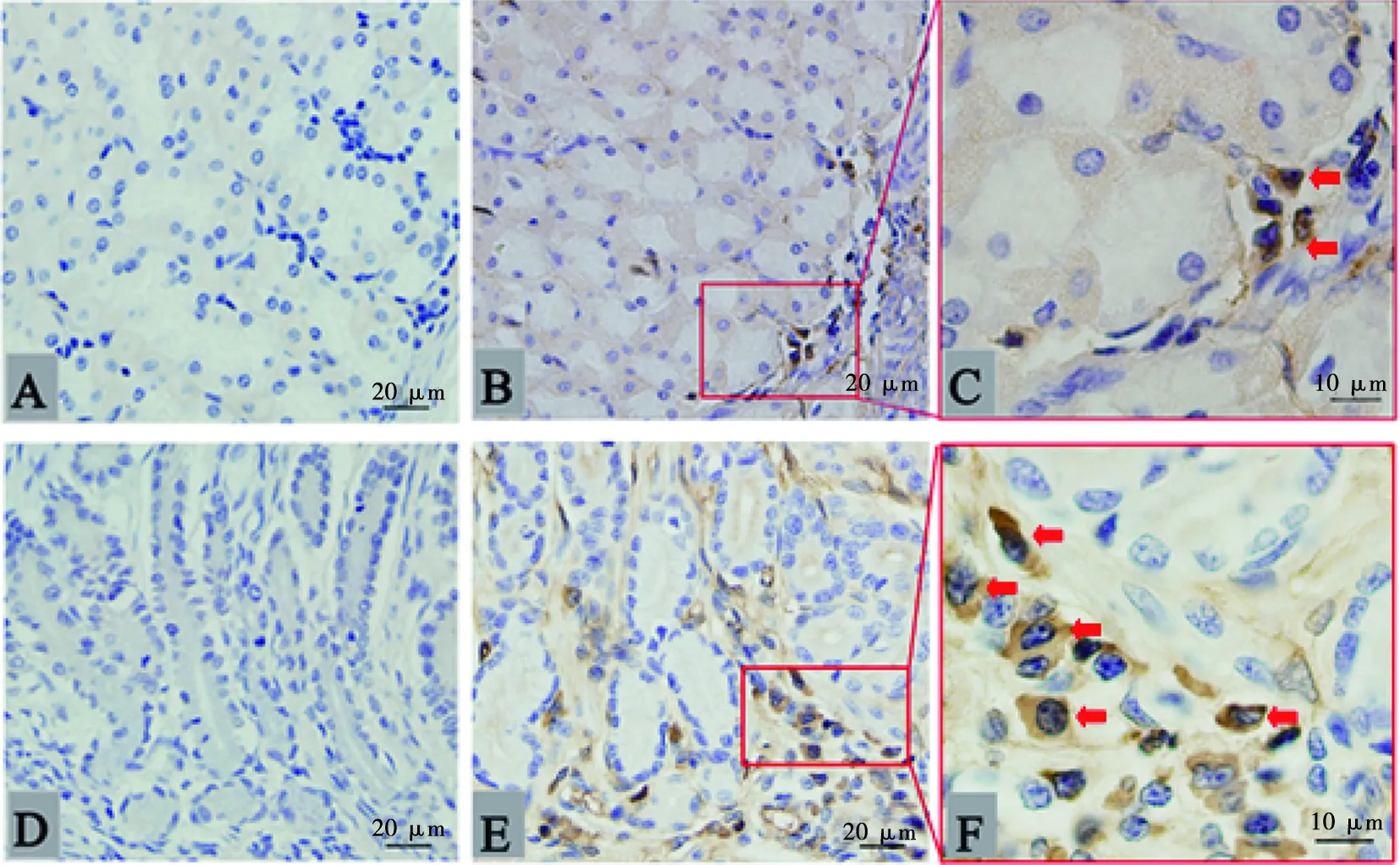
A.胃底腺区阴性对照;B.胃底腺区;C. 胃底腺区局部放大;D.幽门腺区阴性对照;E.幽门腺区;F. 幽门腺区局部放大。箭头所示为IgM分泌细胞
2.2 肠道中IgM分泌细胞的分布特点
肠道内呈阳性的IgM分泌细胞也是胞浆明显着色(图2),与胃中的结构相似。染色结果显示,IgM分泌细胞在各年龄组肠道中的分布位置相似,但存在数量上的区别,以青年组为例,IgM分泌细胞在其各肠段均有分布,但分布位置有所差异。在十二指肠中,IgM分泌细胞主要分布在绒毛固有层和绒毛基部固有层,十二指肠腺周围的黏膜下次也有较多分布,但绒毛顶部固有层分布较少(图2B);空肠和回肠中的IgM分泌细胞分布较为相似,主要分布在绒毛基部固有层,绒毛固有层存在较少(图2E、图2H);而在大肠(盲肠、结肠、直肠)中,IgM分泌细胞主要存在大肠腺周围的固有层中,其次在绒毛基部固有层也有所分布,绒毛固有层中几乎没有(图3)。
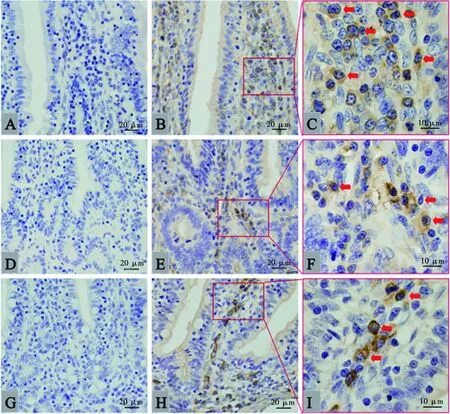
A.十二指肠阴性对照;B.十二指肠;C. 十二指肠局部放大;D.空肠阴性对照;E.空肠;F. 空肠局部放大;G.回肠阴性对照;H.回肠;I. 回肠局部放大。箭头所示为IgM分泌细胞
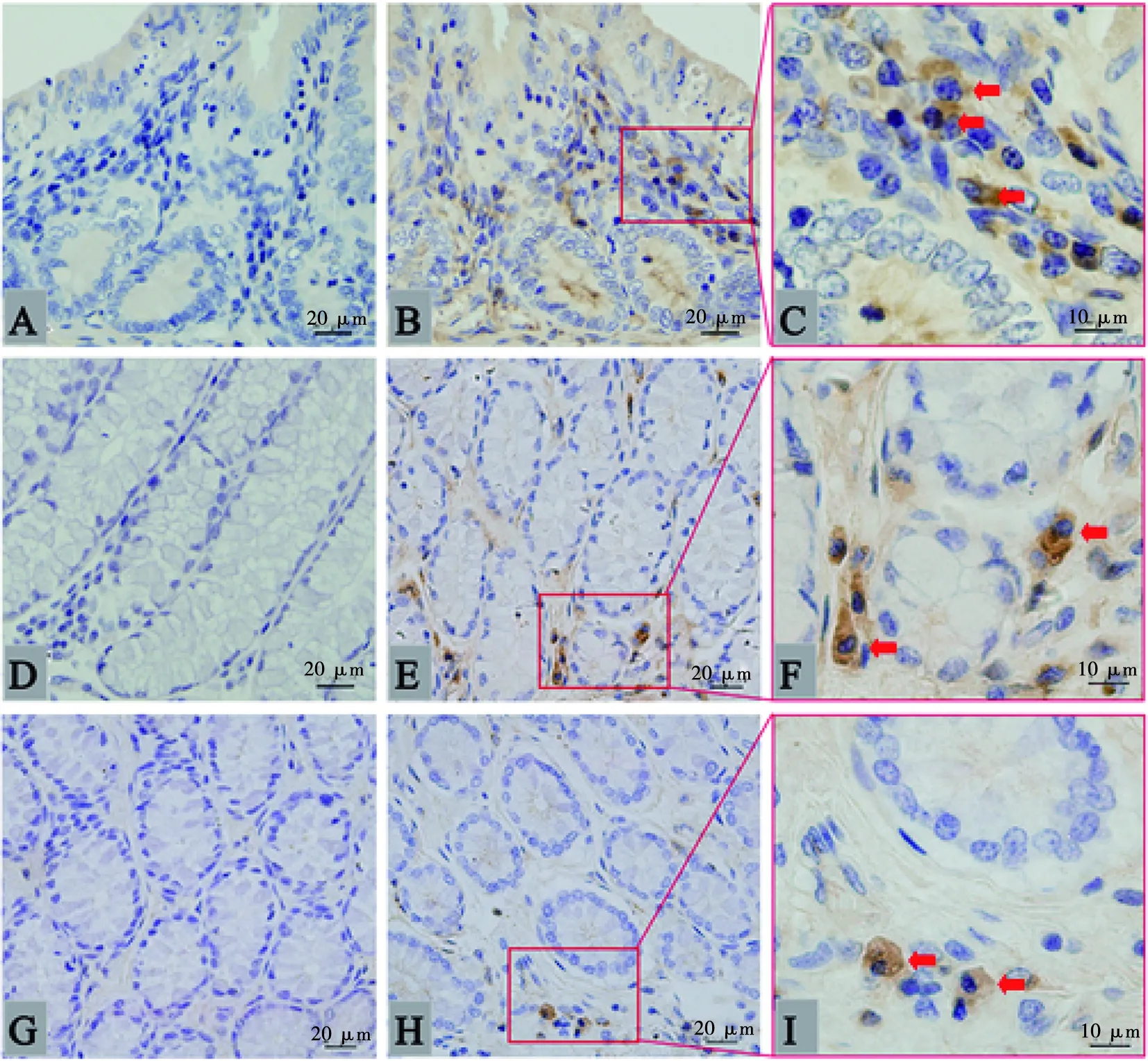
A.盲肠阴性对照;B.盲肠;C. 盲肠局部放大;D.结肠阴性对照;E.结肠;F. 结肠局部放大;G.直肠阴性对照;H.直肠;I. 直肠局部放大。箭头所示为IgM分泌细胞
2.3 各年龄段胃肠道不同部位中 IgM分泌细胞的分布密度变化
统计结果如图4所示,各年龄组IgM分泌细胞在胃肠道呈先增加后减少的趋势,青年组十二指肠中的分布密度最高,老年组直肠中的分布密度最低。在胃部(图4A),各年龄组IgM分泌细胞主要分布于幽门腺区,密度极显著高于胃底腺区(P<0.01)。小肠中(图4B),幼年组和青年组在十二指肠的密度均极显著高于空肠和回肠(P<0.01),幼年组空肠分布密度和回肠相近(P>0.05),青年组和老年组在空肠的密度均显著高于回肠(P<0.05)。大肠中(图4 C),青年组盲肠分别极显著高于结肠和直肠(P<0.01)。IgM分泌细胞在家兔胃肠中的分布密度随年龄增长呈上升趋势,青年期达到顶峰,随后逐渐下降。统计显示,幼年组在胃底腺部(P<0.05)、空肠(P<0.01)、回肠(P<0.01)的分布密度低于青年组;老年组在幽门腺区(P<0.01)、十二指肠(P<0.01)、空肠(P<0.01)、回肠(P<0.05)、盲肠(P<0.01)、结肠(P<0.05)的分布密度均低于青年组。
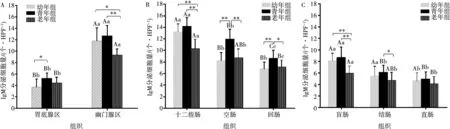
相同部位组间标注*为P<0.05,**为P<0.01;相同年龄组不同部位标注大写字母不同表示P<0.01,不同小写字母表示P<0.05,字母相同表示P>0.05
3 讨论
本试验结果表明,家兔胃肠道IgM分泌细胞主要分布在黏膜固有层、肠绒毛固有层、绒毛基部固有层以及肠腺附近。胃部结果分析可知,各年龄组胃底腺区IgM分泌细胞的密度明显低于幽门腺区,这可能因为胃底腺区的分泌细胞主要是壁细胞、主细胞、颈黏液细胞,这些细胞的分泌物形成了胃底腺区的强酸性环境,使大部分外来病原微生物难以存活,这些因素分担了免疫细胞的部分责任,因此胃底腺区所需的免疫细胞会相应较少。此外还发现, IgM分泌细胞在胃的分布密度不及肠道,这可能是由于家兔为单室胃,物质交换的较少,外来抗原侵入的机会也相对较少。
小肠中IgM分泌细胞的分布密度变化则是十二指肠、空肠、回肠依次减少,车传燕等[11]在断奶仔猪小肠中也发现IgG分泌细胞在十二指肠的数量极高,显著高于空肠,极显著高于回肠。Allen等[12]研究仔猪小肠中IgM分泌细胞的分布规律后,也得出了相似结论。而出现这种趋势的原因很有可能是因为顽强的外来微生物在逃过胃的防御机制后,首先经过十二指肠,该部位肠黏膜内外的物质交换非常频繁,受到各种抗原的不断刺激,承担的免疫压力最大,且大量进入肠道的外源性抗原以及消化液等物质会刺激免疫细胞的发育和成熟[13-14]。因而,此处最有可能发生体液免疫应答,诱导效应B细胞前体细胞发育成IgM分泌细胞,以产生更多的sIgM抵抗外源性抗原的入侵。随着肠内容物后移,大部分抗原通过各种途径被清除掉,因此,到达空肠后段乃至回肠时,IgM分泌细胞数量显著减少,这说明小肠前段可能是发挥免疫功能的主要部位。
大肠作为消化道的后段,能对前段消化道中未吸收的物质进行补偿吸收。家兔是草食性单胃动物,主要通过大肠微生物发酵来吸收纤维素,大量细菌会在此处繁殖。试验结果表明,IgM分泌细胞在大肠中主要分布于盲肠,这可能与家兔盲肠特殊的生理结构有关,盲肠固有层中丰富的孤立淋巴小结和弥散淋巴细胞为IgM分泌细胞的发育提供了充足来源,家兔发达的盲肠,功能与反刍动物的瘤胃相似,主要进行微生物消化,食物残渣在此处停留的时间较长,盲肠复杂的微生物环境使它拥有比结肠和直肠更加强有力的免疫能力,以便应对随时发生的免疫应答。田波等[15]对牦牛黏膜免疫相关细胞在大肠各段的分布研究结果显示,浆细胞在盲肠中分布最多,直肠最少,主要存在固有层肠腺之间,与本试验结果高度相似。
此外,对不同年龄组同一肠段IgM分泌细胞的密度进行对比后发现,青年组数量明显多于幼年组和老年组,其随年龄呈下降趋势,这可能与生理特性以及发育水平有关。Zhang等[16]研究不同年龄双峰驼小肠中IgA和IgG分泌细胞的分布时发现,这两种细胞密度从幼龄到青年呈上升趋势,在青年达到峰值,然后随着年龄的增长逐渐下降。据研究,老年动物淋巴滤泡中B淋巴细胞的分化、增殖和成熟都会受到抑制,从而导致外周血中的浆细胞数量减少[17],而IgA分泌细胞可以通过肠道归巢分子的作用,从外周血中归巢到肠道固有层[18],外周血中的浆细胞数量的减少,可能会相应引起肠道中抗体分泌细胞的减少。但也有一些免疫细胞没有这种变化趋势,随着年龄的增长其分布密度反而升高,如调节性T细胞(regulatory T cell,Treg)和辅助性T细胞(helper T lymphocyte,Th)数量随着年龄的增长而增加[19]。Stervbo等[20]对24名青年人(19~30岁)和26名老年人(53~67岁)的外周血中的免疫细胞进行计数后发现,中间型单核细胞(CD14+ +CD16+)和非经典型单核细胞(CD14+CD16+ +)在老年人中较年轻人多。由此可发现衰老会显著降低IgM分泌细胞在健康家兔胃肠道的分布密度,但不会改变其分布位置,在其他种属动物的胃肠道中IgM分泌细胞以及其他免疫细胞也具有类似的分布模式。
猜你喜欢
现代畜牧科技(2021年5期)2021-07-20
中华胃食管反流病电子杂志(2017年2期)2017-10-27
中国内镜杂志(2017年2期)2017-03-20
中华胃食管反流病电子杂志(2016年2期)2016-10-10
湖南畜牧兽医(2016年1期)2016-06-05
冰雪运动(2016年4期)2016-04-16
动物营养学报(2015年9期)2016-01-07
中国民族民间医药·下半月(2014年1期)2015-02-02
重庆医学(2014年14期)2014-08-14
河北医科大学学报(2011年6期)2011-03-25
