
水分活度对轻腌大黄鱼源蜂房哈夫尼菌碳源利用能力的影响
2021-10-09 01:34王晓阳郭全友黄海潮杨絮张小敏
食品与发酵工业 2021年18期
王晓阳,郭全友,黄海潮,杨絮,张小敏
1(上海海洋大学 食品学院,上海,201306)2(中国水产科学研究院东海水产研究所,上海,200090)3(上海理工大学 医疗器械与食品学院,上海,200093)
大黄鱼(Larimichthyscrocea)属石首鱼科、黄鱼属,2019年养殖产量达22.55万t[1],居全国海水养殖鱼类之首。大黄鱼以冰鲜流通和冻品销售为主,加工品常以盐制[2-3]、冷冻调理和香糟等形式出现,其中盐制大黄鱼又分为盐渍、半干和干制等类型,目前总体趋向低盐、轻微干燥、方便食用等轻腌制品方向发展。研究表明,蜂房哈弗尼菌(Hafniaalvei)是轻腌大黄鱼低温贮藏时的特定腐败菌(specific spoilage organism,SSO)之一[4],可通过代谢分解糖类、蛋白质、脂肪等能源物质,产生小分子异味物,缩短产品货架期和流通半径。另外,微生物在温度、pH、水分活度(water activity,Aw)和抑菌剂等调控作用下,会不断消耗能量来维持内部平衡,超出其耐受范围可导致菌体死亡。因而,从能量代谢角度探究不同胁迫条件下轻腌大黄鱼SSO能源利用和生长动力学,对优化大黄鱼产品保藏及品质控制技术具有重要意义。
微生物能源利用和代谢活动,主要受水分、碳源、氮源、矿物元素和生长因子等影响。碳源是主要能源物质,包括糖类、羧酸类、氨基酸类等,受菌株自身特征和环境胁迫因子影响,能源利用种类和代谢速率存在较大差异。WILLIAMS等[5]采用Biolog技术评价菌株在特定环境中利用各种底物的能力,可确认菌株种间竞争优势,反之,针对食品腐败菌,优化能源种类可实现抑菌目的。郭全友等[6]探究了莓实假单胞菌在15、25、33 ℃下碳源生长状况,发现25 ℃时其总体碳源利用能力和活性最强,15 ℃时次之,33 ℃时最弱。单珂等[7]以轻腌大黄鱼SSO(H.alvei和普通变形杆菌)为对象,探究了NaCl质量分数(1%~5%)和pH(5.0~7.0)对其碳源代谢的影响,结果表明2株菌碳源代谢能力随NaCl质量分数增加而降低,增至3%时几乎完全被抑制。
现今水产品中腌渍、风干和蜜饯等加工手段,主要通过调节Aw达到抑菌目的以延长货架期。绳金房等[8]提出革兰氏阴性菌生长所需最小Aw一般在0.91~1.00,超出或低于最适Aw,导致渗透压变化而影响微生物生长。因此,需以Aw为主要胁迫因子,并对不同调控强度下轻腌大黄鱼SSO的碳源利用进行探究。微生物动态变化常采用修正Logistic、修正Gompertz及Baranyi方程,拟合获得生长动力学参数,最大比生长速率(μmax)、迟滞期(λ)可定量表征其生长状况和代谢速率。VERSCHUERE等[9]采用Gompertz方程,拟合溶藻弧菌(Vibrioalginolyticus)、荧光假单胞菌(Pseudomonasfluorescens)28 ℃培养时碳源利用OD590nm值,获得μmax和λ等动力学参数,对描述微生物生长速率、评价抑菌效应及预测货架期具有重要作用。
本文以源自轻腌大黄鱼的SSO(H.alvei)为研究对象,采用Biolog GEN Ⅲ微孔板测定不同Aw(0.92~0.98)下其碳源利用情况,并采用修正的Gompertz方程拟合得到动力学参数,评价H.alvei对总体和不同类别碳源利用能力的差异,为靶向抑菌、延长轻腌大黄鱼货架期提供一定理论依据,有利于促进大黄鱼加工业产值提高和快速发展。
1 材料与方法
1.1 实验材料
1.1.1 实验菌种
菌株源自5 ℃贮藏货架期终点(29 d)的轻腌大黄鱼,经Sherlock MIS微生物鉴定系统鉴定后,采用16S rRNA测序,确认H.alvei(序列号:KY684258)为优势菌,比例为35.9%,菌株冻干保藏备用。
1.1.2 实验材料
甘油、营养肉汤、营养琼脂,国药集团化学试剂有限公司;Protrol IF-A接种液、GEN Ⅲ鉴定板、BUG培养基,美国Biolog公司。
1.1.3 仪器与设备
ZM-100全自动不锈钢反压高温蒸煮锅,广州标记包装设备有限公司;SW-CJ-IFB超净台,上海博讯实业有限公司医疗设备厂;BIOLOG微生物半自动鉴定仪、SENSITTITRE浊度仪,美国Biolog公司;LAB-Touch水分活度仪,瑞士Novasina公司。
1.2 实验方法
1.2.1 菌株活化
用接种环蘸取H.alvei菌液(-80 ℃甘油保藏),接种于营养肉汤中,25 ℃下摇床振荡(160 r/min)培养24 h;将活化菌液在营养琼脂平板上划线进行2次活化,25 ℃培养24 h,获得单菌落;挑取单菌落,在BUG平板上划线,25 ℃培养24 h。
1.2.2 IF-A接种液制备
IF-A接种原液Aw为0.98,采用灭菌后的80%甘油调节IF-A原接种液,使其Aw值分别为0.92、0.93、0.94、0.95、0.96、0.97、0.98。
1.2.3 接种单菌落并培养
参照Biolog GEN Ⅲ板操作说明,用无菌接菌棒挑取BUG平板上的单菌落至不同Aw(0.92~0.98)的IF-A接种液中,上下滑动使得菌体分散,采用浊度仪将菌悬液透光率调至90%~98%,用八通道电动移液器将菌悬液100 μL加入到GEN Ⅲ微孔板中,25 ℃恒温培养。
1.2.4 测定吸光度
采用BIOLOG微生物半自动鉴定仪中OmniLog读数仪,每隔3 h读取不同Aw(0.92~0.98)条件下菌株OD590nm值,共计培养150 h。
1.2.5H.alvei碳源利用能力评价
碳源利用判定:GEN Ⅲ板中的71种碳源分为5类:糖类(25种)、羧酸类(17种)、氨基酸类(11种)、胺/酰胺类和脂肪酸/酯类(5和6种)、其他类(7种)。当待测孔变为紫色或蓝色判为可利用,待测孔无色时,判为完全不能利用。
碳源利用能力:采用每孔的平均颜色变化率(average well color development,AWCD)[10-11]、碳源利用率(Sm/Sz,Sm表示每类碳源的利用面积,Sz表示全部碳源的利用面积)和动力学参数(μmax和λ)进行描述,AWCD和S计算如公式(1)和公式(2)所示:
(1)
(2)
式中:Ri为对照孔OD值;Ci为含碳源孔OD值;n为碳源孔数,71;Cti为ti时含碳源孔相对OD值。
1.2.6 模型建立
依据DALGAARD[12]的方法,代谢模型采用修正的3参数Gompertz方程[公式(3)]或4参数Gompertz方程[公式(4)]拟合[13],当细菌代谢速率较快,无迟滞期时多采用3参数方程,而迟滞期较长时,采用4参数方程拟合度更优,要求R2>0.95。Vt计算如公式(3)和公式(4)所示:
(3)
(4)
式中:A为颜色变化最大吸光度;a为最小吸光度;b为评估得到的最大吸光度与最小吸光度之差;μmax为最大比生长速率,h-1;λ为延滞期,h;t为时间,h;Vt为t时OD值,即C-R值。
1.3 数据分析
使用Origin 9.0(美国Origin Lab公司)软件对方程进行拟合、作图及分析。
2 结果与分析
2.1 总体碳源利用分析
2.1.1H.alvei碳源利用种类
H.alvei为革兰氏阴性菌,属肠杆菌科,兼性厌氧,代谢包括呼吸和发酵两种类型,能产生硫化氢和酸性风味,导致水产品腐败变质。在水产品面向市场的流通过程中,通常发生脱离冷链或变温情况,因此探究常温(25 ℃)时腐败菌生长繁殖和能源利用是必要的。
表1为不同Aw条件下H.alvei总体碳源利用情况,可知Aw为0.92~0.98时,可利用碳源总数依次为17、29、38、21、20、39和32种,其中糖类、羧酸类在不同Aw下利用情况波动较大,氨基酸类(如L-丙氨酸和L-天冬氨酸)较为稳定。值得关注的是,明胶、吐温40、D-山梨醇、α-羟基-丁酸、L-半乳糖醛酸内酯等在所有Aw(0.92~0.98)下均不被利用,可基于此特性展开相关保鲜剂的研发。IVANOVA等[14]提出吐温40、肌苷、甘油、明胶和壳聚糖等常作为复合膜用于水产品保鲜,张佩华等[15]利用油酸乙酯、吐温80、戊醇制备水包油型微乳液制成复合保鲜剂,抑制腐败菌的生长。以上研究可为优化轻腌大黄鱼产品配方、开发复合保鲜剂以实现有效抑菌提供思路。

表1 碳源分类及利用情况Table 1 Utilization and classification of carbon sources
2.1.2 不同Aw下H.alvei碳源利用率及利用面积
由图1-a可知,Aw为0.92~0.98下羧酸类和氨基酸类碳源利用率(Sm/Sz)较高,均值分别为37.3%和32.6%,糖类、胺/酰胺和脂肪酸/酯类均值较接近,分别为18.7%、8.6%,其他类最低(4.8%)。郭全友等[6,16]研究得出,在15、25、33 ℃下,鱼源莓实假单胞菌和木糖葡萄球菌碳源利用率由高到低均为:糖类、羧酸类和氨基酸类。本研究中H.alvei糖类碳源利用率受Aw影响较大,如Aw为0.98时为33.7%,0.92时降至5.0%,而0.93时又增至32.2%,可能是Aw为0.92时该菌相关糖降解酶活性骤降所致。
碳源利用面积(Sm)可直观反映各类碳源利用能力。如图1-b所示,H.alvei碳源利用Sm均值由大到小依次为:羧酸类、氨基酸类、糖类、胺/酰胺和脂肪酸/酯类、其他类,与Sm/Sz规律相吻合。其中仅羧酸类在Aw为0.97时Sm最大(231.1),剩余四类碳源Sm值均在Aw为0.98时最大。随Aw降低,五类碳源Sm基本呈下降趋势,表明Aw越低,菌株碳源利用能力越弱,针对部分Aw时Sm为相反变化趋势分析原因如下:(1)可能与分解利用碳源相关酶的活性有关,导致代谢速率不同;(2)可能是不同Aw下相同生长基质中菌株生理功能、能源选择及代谢途径存在差异所致[17]。

a-碳源利用率;b-碳源利用面积图1 不同Aw下H.alvei 五类碳源的利用率及利用面积Fig.1 Utilization rates and areas of five carbon sources of H.alvei at different Aw
2.1.3 不同Aw下H.alvei碳源利用AWCD及动力学参数
AWCD是反映微生物对71种碳源总体利用活性的有效指标。由图2-a、图2-b可知,Aw为0.98时AWCD最高达0.292,拟合所得μmax(0.012 0 h-1)远大于其他Aw组,约为4倍以上,表明该Aw下H.alvei总体碳源利用能力最强,菌株生长最快;Aw为0.96时AWCD最大值为0.156,μmax(0.002 99 h-1)较Aw为0.98时次之,即菌株在该Aw下代谢较为旺盛,与轻腌大黄鱼本身Aw值为0.96±0.002联系紧密;根据μmax剩余Aw组由大到小排序为:0.95、0.92、0.93、0.94、0.97,其中Aw为0.97时μmax最小,可能与IF-A接种液与较少量调节介质(甘油)溶解后混合液的结构特性有关;Aw为0.93、0.94时μmax差异不大,几近Aw为0.98时的1/7,表明Aw对该菌总体碳源利用能力影响显著(P<0.05)。
如图2-b所示,Aw为0.96时λ最大(52.1 d),菌株环境适应性差;Aw为0.98时λ最小(4.9 d),可快速进入指数期,且μmax和Sm最大,表明此Aw可极大促进H.alvei生长,加速腐败进程。曹慧等[18]研究表明Aw对馒头中金黄色葡萄球菌μmax、λ影响分别为极显著(P<0.01)、不显著(P>0.05),Aw为0.98时菌株μmax较小,λ较长,本研究结果与之相反,但μmax与λ同样呈反比;Aw为0.92~0.95时随Aw增加λ显现逐渐缩短趋势,即该区间内菌株环境适应能力随Aw增加而增强。PARK等[19]报道Aw影响微生物的生长繁殖及孢子产生,Aw降低时微生物糖类代谢活性减弱,对本文具有一定借鉴意义。

a-AWCD;b-μmax图2 不同Aw下H.alvei总体碳源利用情况Fig.2 Overall carbon source utilization by H.alvei at different Aw
2.2 不同类型碳源利用分析
2.2.1Aw对H.alvei糖类利用的影响
图3为不同Aw(0.92~0.98)下H.alvei每种可利用糖类的动力学参数。由图3-a可知,不同Aw(0.92~0.98)下菌株糖类利用种类依次为2、11、8、3、4、12、10种,μmax均值为0.001 55、0.003 31、0.002 68、0.002 92、0.005 29、0.005 26、0.016 4 h-1。可见随Aw升高总体糖类利用能力呈逐渐上升趋势,Aw为0.98时达到最强,其中以L-鼠李糖μmax最大(0.031 6 h-1),L-岩藻糖μmax次之(0.026 4 h-1),且仅该两种单糖存在明显λ,分别为10.2、7.8 h,表明该Aw下菌株生长初期先利用α-D-葡糖、D-甘露糖、D-麦芽糖等糖类,而指数期L-鼠李糖和L-岩藻糖最易被利用,成为主要的糖类供给。RADKOV等[20]研究发现生物体绝大多数可利用L-氨基酸,D-氨基酸利用较少,可类比解释本实验中L-型糖较D-型糖利用更好的结果。由图3-b可知,Aw<0.98时随Aw增大λ有显著增长趋势,且Aw为0.97时λ最长,β-甲酰-D-葡糖苷λ达77.0 h,说明Aw增加可延缓菌株对碳源的利用,对延长轻微加工大黄鱼货架期有一定指导意义。

a-μmax;b-λ图3 H.alvei利用糖类碳源的动力学参数Fig.3 Kinetic parameters of carbohydrates utilization of H.alvei
2.2.2Aw对H.alvei羧酸类利用的影响
由图4-a可知,不同Aw(0.92~0.98)下H.alvei羧酸类利用种类依次为4、6、13、11、7、12、7种,μmax均值为0.002 77、0.003 20、0.003 02、0.003 55、0.006 22、0.014 9、0.032 2 h-1,可见菌株对羧酸类的利用主要在Aw为0.97、0.98下,μmax数量级均为10-2。Aw为0.98时D-葡糖醛酸被利用能力最强,D-半乳糖醛酸次之且仅在该Aw下被利用,可能由于醛酸类物质具有一定亲水性;Aw为0.97时甲酸μmax最大,D-苹果酸次之,可能是受到碳源分子质量的影响。另外,乙酸在不同Aw下均可被利用,当Aw为0.97时μmax最大(0.017 0 h-1),Aw为0.98时次之(0.015 5 h-1),说明该菌利用乙酸时对Aw的耐受范围较广。乐毅全等[21]和修艳辉等[22]均提出乙酸是希瓦氏菌较为理想的碳源,且对温度耐受性广泛,与本文结果类似。由图4-b可知,不同Aw下羧酸类利用λ均值依次为41.3、39.1、49.6、38.3、30.8、28.2、3.87 h,其中Aw为0.98时λ最小,如D-半乳糖醛酸仅2.2 h,而Aw≤0.97时λ大幅延长,最长达75.0 h。可见羧酸类利用曲线基本呈现Aw越高,μmax越大,λ越小的规律,即Aw越大,羧酸类利用程度越高。修艳辉等[23]探究了15 ℃时腐败希瓦氏菌在Aw调控下的生长/非生长状况,结果表明Aw为0.93时菌株μmax为0.006 h-1,Aw为0.96时μmax增至0.079 h-1,Aw≤0.95时λ>50 h,Aw为0.96时无λ,与本研究结果相呼应,验证了腐败菌生长与底物能源代谢间的对应关系。
2.2.3Aw对H.alvei氨基酸类利用的影响
如图5-a所示,不同Aw(0.92~0.98)下H.alvei氨基酸类利用种类依次为10、10、10、6、7、7、6种,μmax均值为0.002 45、0.003 20、0.004 33、0.007 95、0.008 73、0.006 80、0.026 8 h-1。该菌氨基酸类利用Aw为0.95~0.98,除Aw为0.97外,μmax随Aw降低而降低,与羧酸类、糖类结果相似。各Aw下菌株优先利用氨基酸种类不同,其中γ-氨基-丁酸仅在Aw为0.92、 0.93下被利用但利用程度较低,可能和其低Aw耐受性有关。由图5-b可知,Aw为0.98时菌株氨基酸类利用λ较短,其中L-丝氨酸λ最短(0.3 h),氨基乙酰-L-脯氨酸λ最长(8.0 h);Aw≤0.96 时λ明显延长,如L-谷氨酸、L-丙氨酸、L-天冬氨酸λ分别骤增至60.0、65.4、68.5 h,说明在Aw降至0.96时,该菌生长初期对以上三类氨基酸的适应能力急剧降低。据研究,养殖大黄鱼肌肉中氨基酸含量丰富,主要为谷氨酸、天冬氨酸、赖氨酸等[24],可为H.alvei的生长繁殖提供营养。菌株通过代谢分解鱼体蛋白质、氨基酸及其他含氮物质,可生成氨、胺类、硫化物等,产生腐臭气味,导致鱼体腐败变质。因此,通过调整Aw范围,可一定程度降低氨基酸底物利用率,使腐败菌生长速率减缓,有效延长大黄鱼加工品货架期。
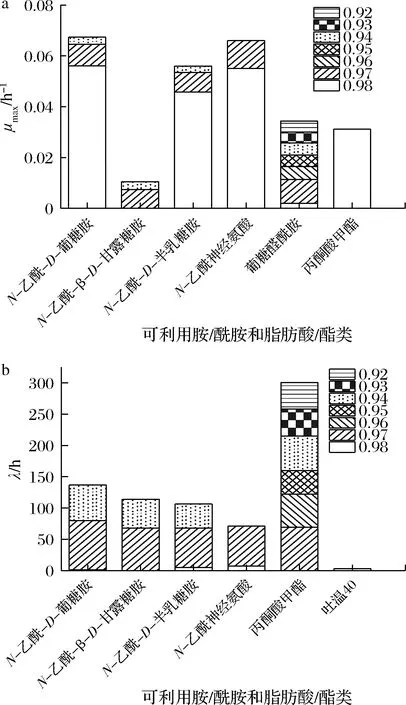
2.2.4Aw对H.alvei胺/酰胺类和脂肪酸/酯类利用的影响
由图6可知,各Aw组H.alvei对胺/酰胺和脂肪酸/酯类碳源均有利用。曲同宝等[25]研究表明糖类和氨基酸类在不同植物群落土壤细菌中均得到较好利用,但胺类均较差,本研究结论与之不同,说明H.alvei对底物利用范围更广。如图6-a所示,Aw为0.92~0.98时菌株胺/酰胺和脂肪酸/酯类利用种类依次为1、1、4、1、1、5、5种,μmax均值依次为0.004 34、0.004 28、0.003 29、0.004 62、0.005 07、0.008 78、0.038 0 h-1,可见Aw为0.98时该类碳源利用速率最高。N-乙酰-D-葡糖胺、N-乙酰-D-半乳糖胺、N-乙酰神经氨酸在Aw为0.98时μmax最大,于Aw为0.97时次之;葡糖醛酰胺在各Aw下均可被利用,Aw≤0.93时菌株仅利用该种胺/酰胺类物质,表明低Aw抑制该类碳源利用,与氨基酸类结果一致。由图6-b可知,Aw为0.98时菌株碳源利用λ最短,其中N-乙酰-D-葡糖胺λ仅1.2 h,N-乙酰神经氨酸λ最长(7.0 h),表明菌株于Aw为0.98时优先利用此类碳源;Aw为0.97时,λ均较长,最长为78.6 h,最短为11.8 h;Aw<0.97时λ均>37.8 h,说明该Aw下此类碳源不是H.alvei前期生长的主要能源。综上,随着Aw增高,菌株利用胺/酰胺和脂肪酸/酯类的μmax相对增大,λ减小,与前述几类碳源变化规律相似。
2.2.5Aw对H.alvei其他类碳源利用的影响
其他类碳源包括肌苷、肌醇等,是不属于糖类、羧酸类、氨基酸类等具有重要作用的碳源物质。由图7-a可知,不同Aw(0.92~0.98)下H.alvei利用其他类碳源的种类依次为0、1、3、0、1、3、4种,μmax均值依次为0、0.002 27、0.003 39、0、0.005 87、0.006 85、0.018 7 h-1。Aw为0.92、0.95时H.alvei不利用此类碳源,可能由于其生长环境的渗透压改变,高渗条件引起细胞质质壁分离,使得细胞失活逐渐消亡[26]。由图7-b可知,D-葡糖-6-磷酸在Aw为0.94、0.97,λ为Aw为0.98时的8、10倍,但μmax分别为其1/9、1/2,表明Aw为0.98时,D-葡糖-6-磷酸μmax最大而λ最短;D-果糖-6-磷酸λ值从大到小依次是在Aw为0.94、0.96、0.93、0.97、0.98时。因此,随Aw降低,菌株利用其他类碳源的λ有增长趋势,进一步验证了胺/酰胺和脂肪酸/酯类碳源利用λ和Aw之间的关系。综上可得,不同Aw下H.alvei利用单一碳源的种类、μmax、λ均有差异,马艳等[27]从碳代谢机制角度发现不同环境因子可能影响H.alvei群体感应信号分子酰基高丝氨酸内酯(acyl-homoserine lactones,AHLs)基因的表达,进而影响AHLs的分泌和生物被膜形成,最终改变该菌的生长繁殖和能源利用速率。因此,通过确定Aw调控范围,探究λ和μmax的关系,可为深入探究环境因子与H.alvei代谢机制的关系提供理论参考。
a-μmax;b-λ图6 H.alvei利用胺/酰胺类和脂肪酸/酯类碳源的动力学参数Fig.6 Kinetic parameters of amines/amides and fatty acids/esters utilization of H.alvei

a-μmax;b-λ图7 H.alvei利用其他类碳源的动力学参数Fig.7 Kinetic parameters of other carbon sources utilization of H.alvei
3 结论
研究发现25 ℃、不同Aw(0.92~0.98)下,H.alvei可利用碳源总数依次为17、29、38、21、20、39、32种,羧酸类和氨基酸类利用率较高,均值分别为37.3%和32.6%,糖类、胺/酰胺和脂肪酸/酯类均值分别为18.7%、8.6%,其他类碳源利用率(4.8%)均值最低。糖类和羧酸类在不同Aw条件下利用情况波动较大,氨基酸类较为稳定。Aw为0.98时对71种碳源的总体利用能力和活性最强,Aw为0.96时次之,剩余组总体碳源代谢能力由强到弱依次为0.95、0.92、0.93、0.94、0.97。
在Aw(0.92~0.98)下,H.alvei生长利用的优势单一碳源主要有:L-岩藻糖、D-岩藻糖、L-鼠李糖、D-葡 糖醛酸、乙酰乙酸、乙酸、D-丝氨酸、L-丙氨酸、L-天 冬氨酸、N-乙酰-D-葡糖胺、L-组氨酸、D-葡糖-6-磷酸和D-果糖-6-磷酸。菌株在Aw为0.96~0.98时对各碳源利用能力较强,Aw<0.96条件下利用程度明显降低。综上表明,Aw对H.alvei不同类型碳源的利用情况有较大影响,且不同Aw下各类碳源λ和μmax变化明显,总体呈现Aw越高,μmax越大和λ越小的趋势,即Aw越高,菌株碳源利用环境适应能力越强且代谢速率越高。通过调节底物碳源因子及环境因子,对H.alvei碳源利用和动力学进行分析,旨在为深入探究腐败菌代谢调控及腐败机制、延长轻腌加工大黄鱼货架期提供理论支撑。
猜你喜欢
海洋开发与管理(2022年10期)2022-11-11
农产品市场周刊(2021年17期)2021-09-28
建材发展导向(2021年7期)2021-07-16
建材发展导向(2021年24期)2021-02-12
建材发展导向(2019年5期)2019-09-09
当代水产(2019年6期)2019-07-25
世界农药(2019年2期)2019-07-13
红领巾·探索(2018年12期)2018-01-26
家庭用药(2018年11期)2018-01-23
渔业致富指南(2018年16期)2018-01-17
