
杨树人工林种间混交对生长性状和食叶害虫抗性的影响*
2021-10-09 05:39陈海波孔祥波张苏芳
林业科学 2021年8期
陈海波 郭 丽 张 真 孔祥波 张苏芳 刘 福
(1.中国林业科学研究院森林生态环境与保护研究所 国家林业和草原局森林保护学重点实验室 北京 100091;2.北大荒农垦集团有限公司八五五农场有限公司 密山 158327; 3.北大荒农垦集团有限公司农业发展部林业草原处 哈尔滨 150090;4.邢台学院生物科学与工程学院 邢台 054001)
发展人工林是解决人类木材需求的重要手段,因此,全球人工林栽植面积快速增长。目前,世界人工林约占森林总面积的7%,而且这一比例还在不断增加,我国人工林面积约为世界的1/3,居世界第一(FAO, 2015)。Liang等(2016)利用来自44个国家和大多数陆地生物群系的777 126块永久地块的地面数据,揭示了生物多样性的持续丧失将导致全球森林生产力加速下降;如能维持好森林生物多样性,仅其生产木材的商业价值每年就可高达1 660亿至4 900亿美元,是实施有效的全球生物多样性保护所需费用的2倍多。这一结论突显了在世界范围内重新评估生物多样性价值、森林管理战略和保护优先事项的必要性。人工林多样性的研究是兼顾解决以上问题的一个重要突破口,特别是面对我国实施天然林保护政策以及生态文明建设的要求,解决我国人工林纯林化严重、生产力不高、生物多样性低、生态服务能力弱等问题,这一研究显得更为迫切。
杨树是我国人工林培育最重要的树种之一,具有适应性强、轮伐期短、优质速生、用途广泛等特点。目前全国杨树总面积已达1 010多万hm2,蓄积量达5.49亿m3,其中人工林约为757.23万hm2,约占全国人工林面积的20%(刘惠兰, 2010; 盛炜彤, 2018)。近年来,利用基因工程对杨树品种进行改造,取得了重要成果,如培育抗逆、抗虫和抗病新品种等,但基因工程树导致的基因稳定性、生物多样性、目标害虫产生抗性及生态系统结构等问题,引起人们对转基因树生态安全性和合理利用的广泛关注(陈盼飞等, 2017; 李丹等, 2015; 李真等, 2017; 吕威等, 2018; 苏晓华等, 2003; 孙振玫等, 2017; 姚丽等, 2016)。对于杨树抗逆性,大量研究集中在高产基因型品种的选育上,而如何运用生态学理论,用不同栽植方式提升人工林的联合抗性,提升生产量和增强抗病虫害能力,是未来人工林研究的重要方向,是丰富和提升人工林经济和生态学价值的重要课题。多树种混交人工林具有明显的抵抗树干害虫的能力,且树种丰富度与混交林抗虫性正相关(李孟楼等, 2005)。杨树混交人工林在生态系统恢复力和抗逆性方面,与单一纯林相比具有明显优势。如用3种杨树与沙棘(Hippophaerhamnoides)混交后,不论是株混还是行混方式均能显著增加杨树生产量,胸径增长6%~38%,平均树高增加8%~23%,地上部分净生产力比纯林高27%~113%(刘世荣, 2000)。新疆杨(Populusalbavar.pyramidalis)与臭椿(Ailanthusaltissima)混交,显著降低光肩星天牛(Anoplophoraglabripennis)的危害率(杨君珑等, 2009)。杨树与刺槐(Robiniapseudoacacia)、杨树与柠条(Caraganakorshinskii)、杨树与紫穗槐(Amorphafruticosa)等混交,均能提升林产品数量和质量,减少病虫害的发生(吴景现, 2009)。
种间多树种混交对生态系统功能的影响主要表现为生态位互补、净生产力增加、天敌昆虫增多等,是加性效应和非加性效应共同作用的结果(Hughesetal., 2016; 申俊芳等, 2015)。近年来研究发现种内多基因型混交具有同样作用,在农业区域多基因型混交后既适合机械化操作,同时抗病虫害、作物产量、杂草控制等显著改善。这一理论已经在我国水稻(Sativaoryza)和德国的小麦(Triticumaestivum)种植上已实际运用,面积约3 000 hm2(Genungetal., 2012; Grettenbergeretal., 2015; Newtonetal., 2009; Schweitzeretal., 2011)。Crutsinger 等(2006)对北美黄花(Solidagoaltissima)研究发现,增加群体基因型多样性能够改变节肢动物群落结构和增加地上净生产力,这种影响效果与种间多样性相当。种植小麦时增加基因型的混种数量,可以有效减少蚜虫种群的生长速度和大小(Shoffneretal., 2013)。在林业方面,杨柳科(Salicaceae)的颤杨(P.tremuloides)、银白杨(P.alba)和黑柳(Salixnigra)3树种混交后,发现种间与种内混交均能提升树木的存活率和生产力,且天敌的数量显著增多(Grossmanetal., 2018)。在短期轮伐人工林中增加柳树基因型混栽后,对整体木材产量、热值成分没有影响,但地面生物量会有显著增加(Hoeberetal., 2018; Schweieretal., 2019)。
综上所述,基于杨树混交人工林的增产和抗虫效果,以及种内多基因型混交的相似作用为理论基础,本文选取杨树人工林4种常用的造林树种,通过杨树种间混交与单一纯林的2种栽植方式,研究2种方式对杨树生长性状和食叶害虫抗性的影响,探索提升人工林生态和经济功能的途径,为人工林科学栽植提供理论依据。
1 材料与方法
1.1 试验地概况及设置
在黑龙江农垦八五五农场(http:∥www.bdh855.com/index.html)选取试验造林样地(45°42′ 28.13″N,131°20′37.05″E),共2.31 hm2,包括有林地1.815 hm2,保护隔离行0.495 hm2。该地区属寒温带大陆性季风气候,年平均积温2 737.9 ℃,降水量541 mm。土壤以黑土为主,土层厚度15~20 cm,有机质含量>100 g·kg-1,pH值在6左右,缓坡向阳,透水性好。2016年5月上旬,选取4种(品种)3年生杨树苗,分别是迎春5号杨(P.nigra×P.simonii‘Yingchun 5’)、小黑杨(P.simonii×P.nigra)、小黑杨-14(P.simonii×P.nigra‘14’)、银中杨(P.alba×P.berolinensis),栽植前用生根粉浸根处理。分别以各树种纯林及4种(品种)随机混合造林,构建5个处理、4个重复区组,共20个小区2 880棵的试验样地。每个小区的株行距是2.5 m×3 m,栽植树苗144棵。2017年6月,更换上年度未成活树苗,确保小区内树种死亡率小于5%。
1.2 生长指标调查方法
2016—2019每年7—8月,在每个小区每个种(品种)随机选择10棵杨树,但小区边缘44棵除外,分别对样树胸径(diameter at breast height, DBH)、高度、冠幅和叶片损伤情况进行调查。每年随机调查杨树320株,并做好位置标记。冠幅从2017年开始测量。用测距测高仪(Apresys PowerLine660)测量树高,用游标卡尺(Deli DL3942)测量胸径。
1.3 叶片受害情况调查
以往的虫情调查多数针对某一种虫口密度,不能反映整株树的食叶害虫密度和叶片受害情况(Vehviläinenetal., 2006)。本研究采用随机抽样,并固定枝条长度,计算虫害叶片面积与叶片数的比例,衡量整株叶片受害情况。每一小区随机选取各品种10棵样树,周边行除外。从4个方向随机分2层剪下长度40 cm的树枝。检查每片叶子受害程度。叶片分为4类: 1)受害叶片面积小于25%; 2) 受害叶片面积25%~50%; 3)受害叶片面积51%~75%; 4)受害叶片面积75%以上。枝条叶片受害比P=(1/4×a+1/2×b+3/4×c+d)/N。其中,a、b、c、d依次为各类别受害叶片数量,N为剪枝的总叶片数。食叶害虫主要有舟蛾类(Notodontidae)、尺蛾类(Geometridae)、刺蛾类(Limacodidae)、毒蛾类(Lymantriidae)、叶甲(Chrysomelidae)、叶蜂类(Tenthredinidae)、潜叶蛾类(Lyonetiidae)和网蝽类(Tingidae)等。
1.4 数据分析方法
调查数据在Excel 2010中进行整理,做多重比较前进行lnX处理,使符合正态分布。采用SPSS 19.0软件进行统计分析,采用Tukey HSD进行单因素方差分析(one-way ANOVA)。
2 结果与分析
2.1 杨树生长性状比较
树高方面,4种(品种)杨树在纯林中的树高生长率存在显著差异(F(3,156)=13.466,P<0.001),银中杨的树高生长率显著大于另外3种(品种),小黑杨-14、小黑杨、迎春5号杨之间无显著差异; 混交林中4种(品种)间的生长率存在显著差异(F(3,156)=24.063,P<0.001),银中杨显著高于其他种(品种),迎春5号杨显著小于银中杨和小黑杨,小黑杨-14和小黑杨之间无差异;各种(品种)在2种栽植方式间无显著性差异(P>0.05)。表明不同栽植方式对树高生长率的影响不显著,而种(品种)间3年生的树高生长率存在显著差异(表1)。
胸径方面,4种(品种)杨树在纯林中胸径生长率存在显著差异(F(3,156)=26.082,P<0.001),银中杨和小黑杨的生长率显著大于小黑杨-14和迎春5号杨,银中杨和小黑杨之间无显著差异,小黑杨-14和迎春5号杨之间无显著差异。混交林中4种(品种)间存在显著差异(F(3,156)=98.810,P<0.001),银中杨显著高于其他种(品种),迎春5号杨显著小于其他种(品种),小黑杨-14和小黑杨之间无差异。银中杨在混交林中胸径生长率显著大于纯林(P<0.05),而迎春5号杨刚好相反(P<0.05)(表1、图1)。可见,胸径生长率在纯林中高的树种在混交林中会更高,而在纯林中表现生长率慢的树种刚好相反。

图1 2种栽植方式杨树胸径生长率变化(2016—2019)Fig. 1 Dynamics of growth rate of DHB of poplar in two planting modes(2016—2019)AB: 银中杨 P. alba× P. berolinensis; NS1: 小黑杨-14 P. simonii× P. nigra ‘14’; NS2: 小黑杨P. simonii× P. nigra; SN: 迎春5号杨P. nigra×P. simonii ‘Yingchun 5’; 下同。The same below.

表1 2016—2019年2种栽植方式条件下杨树的生长性状(均值±标准误)①Tab.1 Growth characteristics of poplar in two planting modes from 2016 to 2019(mean ± SE) %
冠幅方面,在纯林中冠幅生长率存在显著差异(F(3,156)=39.189,P<0.001),银中杨显著高于其他种(品种),迎春5号杨显著小于其他种(品种),小黑杨-14和小黑杨之间无差异。混交林中同样存在显著差异(F(3,156)=19.04,P<0.001),情况与纯林一样。2种栽植方式比较中,各种(品种)在纯林中冠幅均高于混交林,其中迎春5号杨纯林冠幅显著大于混交林(P<0.05)(表1、图2)。
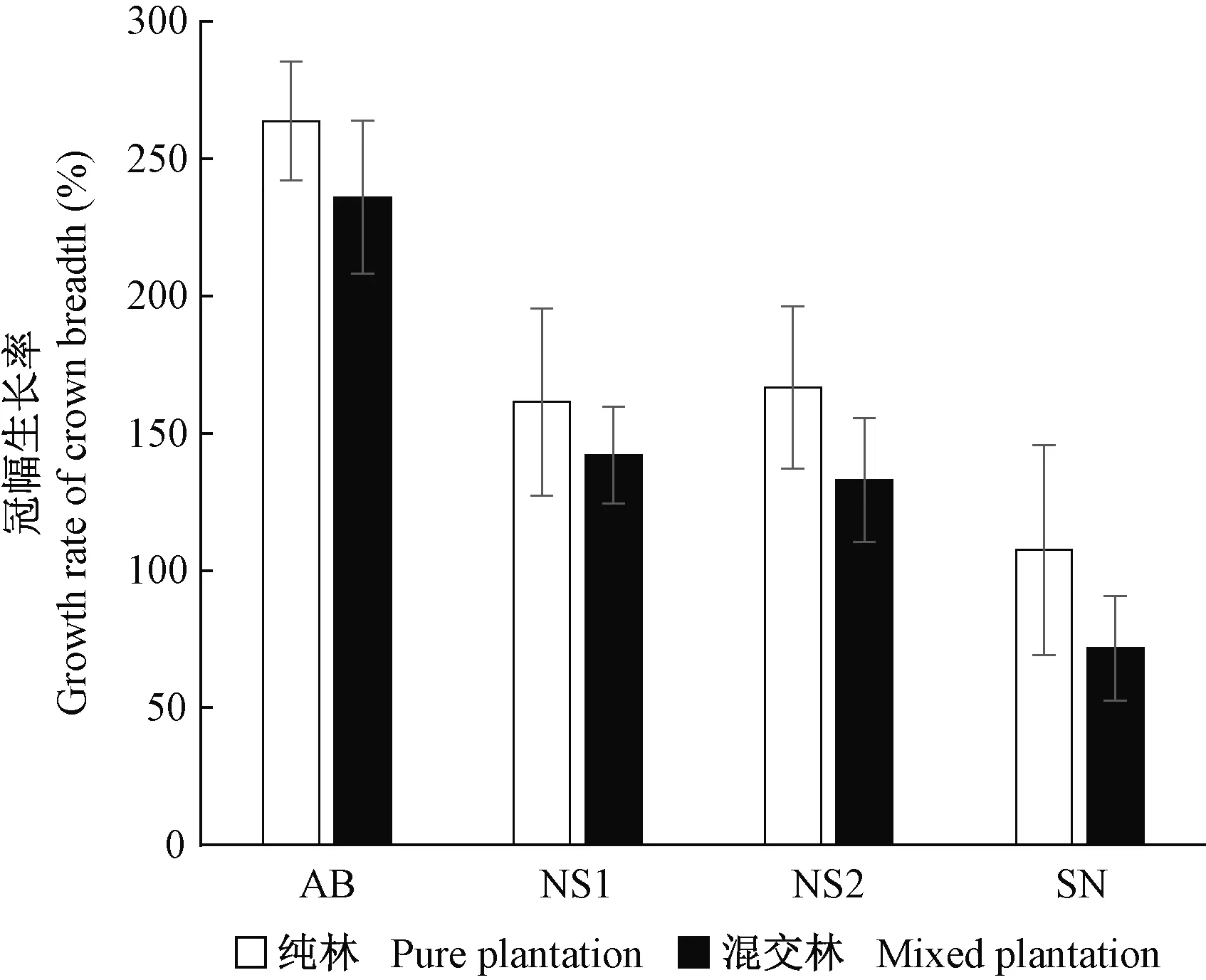
图2 2种栽植方式杨树冠幅生长率变化(2017—2019)Fig. 2 Dynamics of growth rate of crown breadth of poplar in two planting modes(2017—2019)
2.2 纯林与混交林的叶片受害情况
连续3年虫害调查表明, 2017年(F(4,195)=9.009,P<0.001)、 2018年(F(4,195)=12.302,P<0.001)、 2019年(F(4,195)=6.747,P<0.001)各处理间抗虫性差异显著。3年内随着树龄增加,各处理叶片受害率均降低。各种(品种)的抗虫性强弱依次是银中杨>小黑杨>小黑杨-14>迎春5号杨,其中小黑杨与小黑杨-14的叶片受害率差异不显著,迎春5号杨是易感品种,在2017、2018年叶片受害率显著高于其他处理(P<0.05)。2017、2018年混交林的叶片受害率明显低于小黑杨-14和迎春5号纯林,综合抗虫性表现介于4种(品种)中间。小黑杨、小黑杨-14和迎春5号杨在2017与2019年的比较中,受害率显著降低(P<0.05),而银中杨与混交林叶片受害率却没有显著差异,说明混交林与银中杨抗虫性方面表现相对稳定(表2)。

表2 纯林与混交林的叶片受害率(均值±标准误)①Tab.2 Leaf damage rate among different treatments(mean ± SE) %
2.3 混交林中杨树叶片受害情况
如表3所示,混交林中2017年(F(3,156)=7.464,P=0.002)、 2018年(F(3,156)=11.103,P<0.001)和2019年(F(3,156)=3.966,P=0.007)各种(品种)叶片受害率存在显著差异。2017年各种(品种)受害率均高于2018和2019年,表明混交林内随着树高和冠幅的增长,叶片受害率降低,其中小黑杨-14和迎春5号杨叶片受害率连续3年逐年降低。银中杨的叶片受害率显著低于其他种(品种),这一结果与在纯林中比较的结果一致。小黑杨-14叶片受害率高于小黑杨,在2017、2018年的混交林中差异显著(P<0.05)。

表3 混交林中杨树叶片受害率情况(均值±标准误)Tab.3 Leaf damage rate of poplar in mixed plantation (mean ± SE) %
2.4 2种栽植方式中杨树叶片受害情况
银中杨、小黑杨-14和小黑杨在混交林中的叶片受害率普遍低于纯林,并且受害率总趋势逐年降低。迎春5号杨在纯林处理中是叶片受害率最高的树种,在混交林中受害表现更为严重,并且2018、2019年混交林中的叶片受害率比纯林严重。银中杨在2种栽植方式中是受害率最低的树种,在混交林中受害率更低, 2018年2种栽植方式中差异明显(P=0.02),受害率显著低于其他3树种(P<0.05)。对于2017、2018年小黑杨的叶片受害率,纯林与混交林存在显著差异(P=0.002、P=0.033)。在2019年迎春5号杨的叶片受害率混交林显著比纯林严重(P=0.037)。

图3 2017—2019年2种栽植方式中杨树叶片受害率变化Fig. 3 Dynamics of leaf damage rate of four poplar species (cultivar) in two planting modes from 2017 to 2019
3 讨论
本研究表明在生长性状方面,树种(品种)间的差异显著大于栽培方式的影响,这一结果与郭丽等(2020)2013—2016年对河南孟州杨树无性系林的生长性状调查结果不同,表明杨树无性系间混交林生长差异小于种间混交林的生长差异。银中杨是4树种(品种)中的优势树种,在2种栽植方式中胸径增长率存在显著差异,而迎春5号杨胸径和树高的增长率明显低于其他树种,作为4树种(品种)中的劣势树种,在混交林中的胸径增长率与纯林相比存在显著差异。同样,在叶片受害率方面,银中杨在混栽中的受害情况低于其在纯林中的表现,而迎春5号杨却刚好相反。以上研究结果表明,在混交林中杨树树种间的竞争,存在强者更强、弱者更弱的生物竞争法则。这一结果符合生物学角度的加性效应(Additive effects),不同基因型组成的生态系统功能效应不同、相互独立,即存在优势效应(Fox, 2005)或选择效应(Huston, 1997),使优势基因型在多样化系统中获得更多优于单一基因型的公共资源,从而多样性高的系统保留有利基因型的概率要高于单一基因型系统。因此,结合本文杨树在2种栽培方式中生长量结果及生态学加性效应理论,为满足人工林生态和经济需要,提高生物稳定性水平,需要在加强高效新种培育的基础上,强化栽培方式研究,建立良种与良法配套的人工林经营体系。
在叶片受害率方面,混交林2017—2019年在6.1%~8.96%之间,与纯林银中杨3.05%~4.30%和纯林迎春5号杨8.34%~17.32%分别存在显著性差异。在3年叶片受害率调查中,混交林叶片受害率逐年降低,变化范围很小,仅次于抗性最强的银中杨,明显小于其他树种(品种)纯林。以上结果表明杨属树种(品种)间混交能够提高人工林抗虫性,且抗虫稳定性与优势树种相当。这一结果符合生物学角度的非加性效应(Non-additive effects),不同基因型对系统功能效应是非独立的、不可加的,而是交互效应的结果。Castagneyrol等(2017)发现混交林通过改变叶片性状和周边寄主,间接影响昆虫的食草性,从而提高抗虫性; 树种不同功能性状的基因型,由于其形态、物候或生态生理属性的差异,会影响生态系统功能,进而影响生物量生产力(Violleetal., 2007),本文的结果与以上研究结果一致。混交林抗虫性的提升,主要是树种间联合抗性的作用。首先,联合抗性导致害虫在混交林中定位和寻找寄主植物的能力降低(Anderssonetal., 2013; Root, 1973)。不同植物间物理和化学性状的差异,使得与混交林相比害虫更喜欢寻找一致性的纯林林分(Damienetal., 2016; Jacteletal., 2011; Zhangetal., 2004),但也有研究发现随着混交林树种增加,杂食类害虫数量也增多,但种类没有变化。而专食类却刚好相反,害虫数量没有变化,但种类增多(Zhangetal., 2016)。 其次,不同的寄主来源,食物的切换可能影响食叶害虫的消化,进而影响生长发育(Kosetal, 2015),这是因为昆虫取食量取决于植物的叶片韧性、含水量、碳氮比和次生代谢产物等性状(Lorangeretal., 2013; Moreiraetal., 2014; Pearse, 2011)。以上2种是自下而上(Bottom-up)效应调控机制引起联合抗性的结果。最后,“天敌”假说(Enemies hypothesis)认为在纯林中天敌的数量或效果远不如在多样化林分中突出,因为多样化可以为天敌提供可选择的猎物、庇护所或有利的小气候的植物种类更多(Root, 1973)。同时,多样化环境中害虫不断地寻找寄主,增加了遇到天敌的风险(Straubetal., 2013)。这一假说解释了自上而下(Top-down)效应调控引起的联合抗性结果。结合本文研究结果,表明杨树人工林树种多样化栽植方式能够提升林分的联合抗性,一定程度降低虫害发生率,提升林分质量。
4 结论
本研究证实4种(品种)杨树混交林种(品种)间差异显著大于栽培方式影响,银中杨是4种(品种)中的优势树种,迎春5号杨是相对弱势树种。4种(品种)在混交林中生长性状表现,只有银中杨胸径生长率显著增加,各种(品种)生长性状表现出加性效应,种(品种)间的竞争存在强者更强、弱者更弱的竞争法则。小黑杨抗虫性优于小黑杨-14。混交林叶片受害率连续3年相对稳定,与银中杨纯林相似,表现出优势树种的抗虫性,是加性效应的结果。迎春5号杨在混交林中叶片受害率最大。除银中杨,混交林中各树种叶片受害率低于单一纯林,表现出联合抗性的非加性效应。因此,杨树人工林造林要加强树种的多样性化配置,最好是在优势树种间混栽,可以提升联合抗性、降低病虫害发生率,提升林分质量和产量。
猜你喜欢
电影文学(2022年4期)2022-05-20
保健与生活(2022年8期)2022-04-08
时代邮刊·下半月(2022年1期)2022-03-22
共产党员·下(2018年6期)2018-12-05
方圆(2018年23期)2018-01-07
中国火炬(2013年2期)2013-07-24
