
马缨丹在镉、铅、锌复合胁迫下的耐性和解毒机制
2021-09-28 04:16:06谢倚慧张明华蒲玉琳徐小逊张世熔贾永霞四川农业大学资源学院四川成都630四川农业大学环境学院四川成都630
生态与农村环境学报 2021年9期
谢倚慧,张明华,熊 瑞,李 婷,蒲玉琳,徐小逊,李 云,张世熔,贾永霞① (.四川农业大学资源学院,四川 成都 630;.四川农业大学环境学院,四川 成都 630)
近年来,矿产开采和冶炼导致大量重金属释放、迁移进入土壤,造成严重的土壤重金属污染[1-2]。四川铅锌矿产资源丰富,是我国5大铅锌矿生产基地之一,分布有较多共生或伴生镉的铅锌矿[3]。矿山开采带来经济效益的同时,也使矿区及周边土壤受到不同程度的重金属污染,其中以镉、铅、锌复合污染最为严重[4]。土壤中过量的重金属易被植物吸收,抑制植物根系生长,影响植物养分吸收;同时,过量的重金属能破坏植物细胞结构,抑制叶绿素合成和多种生理代谢过程,造成植物黄化、枯萎,甚至死亡[5-6]。
为提高自身对重金属的适应性,不少植物形成了独特的解毒机制,主要有外部排斥机制和内部耐受机制[7]。前者指植物可通过某些途径以减少自身对重金属的吸收,或将吸收的重金属元素大部分固定在根系中,限制其向地上部运输。如石菖蒲[8]以及木贼和蜈蚣草[9]等可通过减少对铅、镉的吸收来提高自身耐性,而覆瓦栎[10]以及香附子和土荆芥[9]则是通过根系的固持作用来缓解镉、铅、锌对植物地上部的毒害。内部耐受机制指重金属在植物体内以不具生物活性的解毒形式存在。区隔化作用是重要的内部耐受机制之一,它主要是将重金属储存在液泡或细胞壁,使之与细胞中代谢活跃的器官分隔开,从而降低重金属对植物细胞的伤害[11-12]。研究发现,蓖麻可将吸收的镉、锌固定在细胞壁上以缓解毒性[13];而萱草[14]和天蓝遏蓝菜[15]体内的镉、铅则主要存在于可溶性组分中,通过液泡区隔化以降低毒害。植物体内的蛋白质、果胶酸盐、草酸盐等物质也可与重金属结合,改变其存在形态,影响其毒性和迁移能力[16-17]。镉在华中蹄盖蕨[18]和大豆[19]中主要以氯化钠提取态和醋酸提取态等毒性较低的形式存在,而圆叶无心菜[20]和波斯菊[21]体内的铅、锌则是以毒性更低的醋酸提取态和盐酸提取态存在。以上研究表明不同植物对重金属的解毒机制不同,研究重金属在植物各器官中含量、亚细胞分布及化学形态特征对明确植物解毒机制具有重要意义。
马缨丹(Lantanacamara)又名五色梅,为马鞭草科马缨丹属常绿小灌木,是一种花、叶两用的观赏性植物,广泛分布于我国四川、云南、浙江等地。马缨丹生长速度快,生物量大,且生态适应性强,不仅能在四川省汉源县富泉铅锌矿区自然生长,而且是该矿区的一种优势植物,表明马缨丹对镉、铅、锌复合污染可能具有较高耐性。前期研究[22-23]发现,马缨丹在镉、铅单一污染条件下可通过减少镉、铅向地上部转运和区隔化作用来缓解重金属毒害。但马缨丹在镉、铅、锌复合污染条件下的耐性和解毒机制尚不明确。因此,以汉源县富泉铅锌矿区马缨丹为材料,通过盆栽试验,研究其在镉、铅、锌复合污染条件下,各部位重金属的含量、亚细胞分布模式和赋存形态,探讨马缨丹在复合污染条件下耐性和解毒机制,以期为矿区植被恢复和重金属污染土壤治理提供一定参考依据。
1 材料与方法
1.1 供试材料
供试马缨丹采自四川省汉源县富泉铅锌矿区。试验所用土壤采自四川省成都市温江区公平镇农用地。土壤基本理化性质:pH值为6.74,有机质含量为21.1 mg·kg-1,碱解氮含量为65.4 mg·kg-1,速效钾含量为212.1 mg·kg-1,速效磷含量为47.5 mg·kg-1,全镉含量为0.072 mg·kg-1,全铅含量为68.4 mg·kg-1,全锌含量为94.8 mg·kg-1,均低于GB 15618—2018《土壤环境质量 农用地土壤污染风险筛选标准(试行)》风险筛选值。
1.2 试验方法
试验于2017年6—10月在四川农业大学成都校区塑料大棚内进行。试验所用土壤经风干、压碎、过5 mm孔径筛后,装入40 cm×30 cm塑料盆中,每盆装土8.0 kg。试验共设置8个重金属处理含量(表1),以不添加重金属为对照,每个处理重复5次。将相应量的CdCl2·2.5H2O(分析纯)、ZnSO4·7H2O(分析纯)和CuSO4·5H2O(分析纯)配制成溶液,与土壤混合均匀,放置4周,使重金属达到平衡状态。
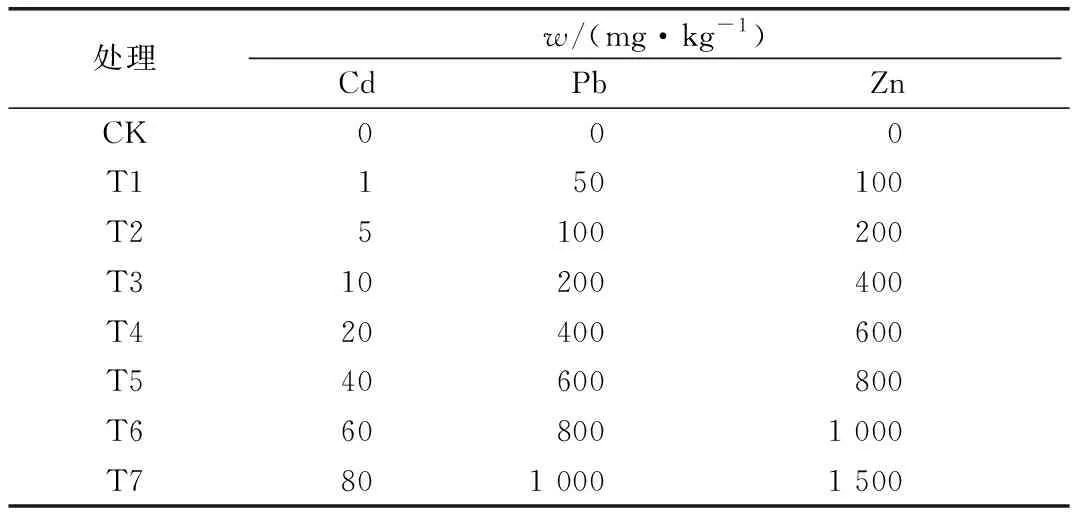
表1 不同Cd、Pb、Zn含量处理Table 1 Different treatments of Cd, Pb, Zn concentration
马缨丹采用扦插方式育苗。剪取10~15 cm左右当年生马缨丹枝条作为插穗,待扦插成活长出2~3片新叶后,选取长势基本一致的幼苗进行移栽,每盆3株。在马缨丹生长期间,定期浇水、除草,观察植物生长情况。于90 d后收获植株,进行各项指标测定。
1.3 测定方法
1.3.1生物量的测定
将采集植物样品分为地上部和根系2个部分,经洗净、烘干后测定干重。
1.3.2相对电导率的测定
电导率采用DDS-12A型电导率仪测定,相对电导率为煮沸前电导率与煮沸后电导率的比值。
1.3.3重金属含量的测定
植物样品中重金属含量测定参照ZHANG等[24]的湿灰化法:采用V(HNO3)∶V(HClO4)=4∶1的混合液将植物样品进行消化,采用原子吸收光谱仪(AAS)测定重金属含量。
1.3.4亚细胞分级
亚细胞组分的分离参照FU等[25]的亚细胞分级方法:采用差速离心法将植物样品分离为细胞壁、细胞器和可溶性组分3个部分,分离出的各组分采用V(HNO3)∶V(HClO4)=4∶1混合液消化后,采用原子吸收光谱仪(AAS)测定重金属含量。
1.3.5重金属形态测定
重金属形态测定参照白雪等[26]的逐步提取法并进行改进:分别采用w为80%乙醇、去离子水、1 mol·L-1氯化钠溶液、w为2%醋酸、0.6 mol·L-1盐酸逐级提取植物样品中重金属,提取液及残渣采用V(HNO3)∶V(HClO4)=4∶1混合液进行消化后,采用原子吸收光谱仪(AAS)测定重金属含量。各提取液对应形态分别为乙醇提取态、水提取态、氯化钠提取态、醋酸提取态和盐酸提取态,残渣经消化后为残渣态。
1.4 数据处理与分析
采用Excel 2010和SPSS 22.0进行数据统计与分析,采用最小显著差法(LSD)对差异显著性(P<0.05)进行多重比较。
2 结果与分析
2.1 镉、铅、锌复合污染对马缨丹生物量的影响
不同镉、铅、锌含量处理马缨丹生物量见表2。
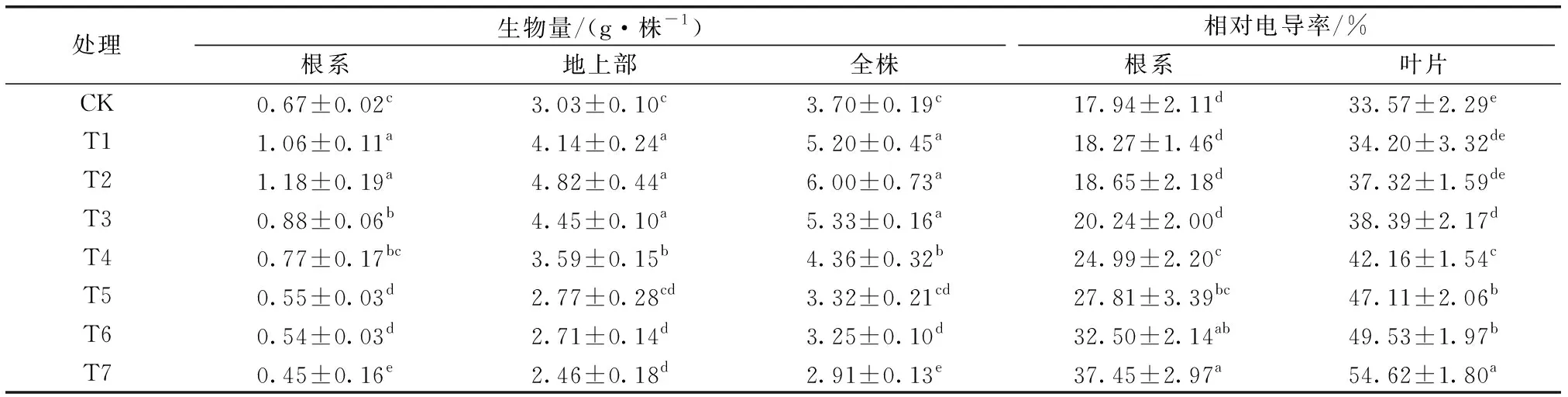
表2 不同Cd、Pb、Zn含量处理马缨丹各部分生物量和相对电导率的变化Table 2 The dry weight and relative electrical conductivity of Lantana camara under different treatments of heavy metal concentrations
如表2所示,当镉、铅、锌含量从CK增至T2处理时,马缨丹根系、地上部和全株生物量随重金属处理含量增加而增加,且以T2处理为最大,分别为CK的1.76、1.59和1.62倍;当污染含量大于T2处理时,马缨丹各部位生物量随重金属含量增加而呈不断下降趋势,T5处理马缨丹根系生物量较CK显著降低,T6处理地上部和全株生物量较CK显著降低。T7处理马缨丹根系、地上部和全株生物量最小,较对照分别降低32.8%、18.9%和21.4%。在高含量重金属复合污染条件下,马缨丹生长才会受到抑制,表明其对镉、铅、锌复合污染具有较好耐性。
2.2 镉、铅、锌复合污染对马缨丹相对电导率的影响
不同镉、铅、锌复合处理马缨丹根系和叶片相对电导率变化情况见表2。如表2所示,随着重金属处理含量增加,马缨丹根系和叶片相对电导率呈现逐渐上升趋势,分别于T3和T4处理时显著高于CK,且均在T7处理时达到最大,分别为CK的2.08和1.45倍。由此可见,随着重金属复合污染程度加重,马缨丹细胞膜受损害程度不断加深。
2.3 马缨丹各部位中镉、铅、锌含量
马缨丹根系和地上部镉、铅、锌含量见图1。如图1所示,随着镉、铅、锌复合污染程度加重,马缨丹地上部和根系镉含量先升高后降低,且在T4处理时达到最大值,分别为CK的8.25和11.07倍。马缨丹地上部和根系铅、锌含量则随着污染程度加重而逐渐上升,在T7处理时达到最大,分别为CK的10.47、6.15倍和23.35、20.80倍。各处理马缨丹根系镉、锌含量均高于地上部,而地上部铅含量则高于根系,表明马缨丹根系对镉、锌具有较好的固持能力。
2.4 马缨丹各器官中镉、铅、锌的亚细胞分布
马缨丹各器官中镉、铅、锌的亚细胞分布情况见图2。如图2所示,在不同含量镉、铅、锌复合污染条件下,马缨丹根、茎、叶中镉、铅主要存在于可溶性组分(45.2%~62.5%和44.2%~66.7%)中,细胞壁(29.7%~44.0%和25.2%~44.1%)次之,细胞器(7.1%~12.2%和6.0%~13.1%)中分布最少。随着复合处理含量升高,马缨丹各器官中镉、铅在可溶性组分中占比降低,在细胞壁中占比提高,在细胞器中占比变化不明显。马缨丹各器官中锌主要分布于细胞壁和可溶性组分中,且两者占比相差不大,分别占总量的39.8%~50.4%和39.2%~52.4%,而在细胞器中占比较少,仅占总量的4.8%~11.6%。提高重金属含量对锌在马缨丹各器官中占比的影响不明显。
2.5 马缨丹各器官中镉、铅、锌的化学形态特征
不同形态镉、铅、锌在马缨丹各器官中的分配存在差异(图3)。镉在马缨丹根、茎、叶中主要以氯化钠提取态和醋酸提取态(50.7%~73.2%)形式存在,乙醇提取态和水提取态(14.2%~20.8%)占比较少。当镉、铅、锌复合含量增加时,马缨丹根中氯化钠提取态镉占比增加,醋酸提取态镉占比降低,而在茎、叶中变化规律却相反,其他形态镉在根、茎、叶中占比变化不大。铅在马缨丹各器官中占比规律与镉相似,主要以氯化钠提取态和醋酸提取态为主,占总量的47.9%~58.9%,乙醇提取态和水提取态较少,仅占总量的14.6%~28.4%;随着污染程度加重,马缨丹叶片中乙醇提取态铅占比降低,水提取态铅占比上升,而在根、茎中两者变化不明显;在各器官中,氯化钠提取态铅占比随重金属含量增加而降低,醋酸提取态、盐酸提取态和残渣态铅占比却随重金属含量增加而增加。锌在马缨丹各器官中主要以氯化钠提取态和醋酸提取态(46.7%~63.2%)形式存在,乙醇提取态和水提取态锌(19.6~32.2%)次之,残渣态和盐酸提取态锌(12.8%~21.1%)占比最小;随着重金属含量增加,马缨丹根部乙醇提取态锌占比增加,醋酸提取态锌占比降低,而在茎、叶中,前者变化不明显,后者则有所上升;在各器官中,氯化钠提取态锌占比随重金属含量增加而降低,盐酸提取态、残渣态锌则相反,而水提取态锌占比变化不明显。
3 讨论
镉、铅、锌等重金属易被植物吸收并积累,积累超过一定程度时,会对植物产生毒害作用,使植物表现出叶片失绿、生长迟缓、植株矮小等症状;然而,少量重金属却对植物生长有一定促进作用[16,27]。研究[28]表明,低水平重金属复合污染可刺激玉米幼苗生长,高水平污染才会对玉米幼苗生长表现出抑制作用,且随着胁迫水平增加,抑制作用逐渐增强。笔者试验发现,低含量镉、铅、锌复合污染条件下马缨丹生物量显著高于CK,在高含量污染条件下,马缨丹生物量才随污染程度加重表现出逐渐降低趋势,直至T6〔w(Cd)、w(Pb)和w(Zn)分别为60、800和1 000 mg·kg-1〕处理,土壤重金属含量远高于GB 15618—2018中风险筛选值〔w(Cd)、w(Pb)和w(Zn)分别为0.3、120和250 mg·kg-1〕和风险管制值〔w(Cd)和w(Pb)分别为3.0和700 mg·kg-1〕时才较CK显著降低。这表明马缨丹能在高含量镉、铅、锌复合污染条件下正常生长,对镉、铅、锌具有较高耐性。T7处理马缨丹根系和地上部生物量均达到最小,且根系生物量下降幅度(32.8%)大于地上部(18.9%),表明在同样高含量复合污染水平下,马缨丹地上部受到的毒害作用小于根系。
重金属离子可与巯基和磷脂类物质反应,破坏细胞膜蛋白和磷脂类结构,使得细胞膜透性增加,大量电解质外渗,引起离子平衡失调和细胞代谢紊乱[29]。相对电导率是表征膜透性的重要指标之一。研究[30]发现,在镉、铅、锌单一污染条件下,天竺葵叶片相对电导率随重金属含量上升而显著上升。笔者试验结果表明,随着镉、铅、锌复合污染程度加重,马缨丹根系和叶片相对电导率不断上升,但直到T3处理才与CK表现出显著差异,表明马缨丹对低含量重金属胁迫有一定耐受能力,但随着污染程度加重,马缨丹细胞膜损害程度加深。T7处理马缨丹根系和叶片相对电导率均达到最大,且根系相对电导率上升幅度(2.08倍)大于叶片(1.45倍),表明在高含量镉、铅、锌复合胁迫下,马缨丹叶片损害程度小于根系。这可能是马缨丹地上部受重金属毒害作用低于根系的原因。
重金属在植物体内各器官中的积累和分布与其耐性密切相关。研究[31-32]发现,在复合污染条件下,铅、锌可抑制川芎和天蓝遏蓝菜对镉的吸收,提高植物耐性。对比前期研究,镉、铅、锌单一污染条件下,马缨丹各部位中重金属含量随污染含量升高而升高[22-23,33];而笔者研究中,随着镉、铅、锌复合含量增加,马缨丹各部位中铅、锌含量逐渐增加,镉含量则表现出先增加后降低的趋势。这可能是由于镉、铅、锌均为二价阳离子,较多的铅、锌离子会占据根系表面的吸附位点[34],使得在复合污染条件下,马缨丹根系对镉的吸收减少,减轻了镉对马缨丹的毒害,增强了马缨丹对复合污染的耐性。在不同含量镉、铅、锌单一污染条件下,马缨丹根系中镉含量始终高于地上部;在低铅含量条件下较地上部更高,在高铅含量条件下较地上部更低;锌含量则相反[22-23,33]。笔者试验中,在复合污染条件下,马缨丹根系铅含量低于地上部,而镉、锌含量均高于地上部,表明在重金属胁迫下,马缨丹可将大部分镉、锌固定在根系中,减轻其对地上部的伤害,增强自身耐性。对比马缨丹在单一和复合污染条件下各部位重金属分布情况,发现复合污染时镉含量与单一污染时相同,复合污染时铅、锌含量则与单一污染时不同,复合污染条件下,马缨丹地上部铅含量均高于根系,锌含量均低于根系,表明复合污染对铅在马缨丹体内的转运有一定促进作用;对锌则有一定抑制作用。简敏菲等[35]发现,镉、铅复合处理时,镉可促进丁香蓼体内铅的转运;而在镉、锌复合污染条件下,镉会抑制柔毛委陵菜(Potentillagriffithii)体内锌向地上部转移[36]。这是因为一方面,植物根系中核酸、蛋白质和多糖可与重金属结合形成大分子物质或不易溶解的有机分子沉积,将其固定在根部,提高自身耐性。但马缨丹根系对重金属的固持能力有限,在复合污染条件下,Cd2+、Zn2+与Pb2+间相互竞争,抢占结合位点,从而促进铅向地上部转运;另一方面,重金属离子向地上部的运输通过某些载体完成,不同金属离子可能是由相同的转运蛋白介导,因此,在镉、铅、锌复合污染条件下,Cd2+、Zn2+可能会抢夺同一运输载体,从而抑制锌向地上部的转运[37]。由此可见,重金属离子由根系转运至地上部时,相互之间的作用是复杂的,具体作用机制还有待进一步探究。
区隔化作用是植物在重金属胁迫下重要的内部耐受机制之一。植物细胞壁上含有大量果胶酸、多糖和蛋白质等物质,可与重金属离子结合,将其固定在细胞壁中,限制重金属在细胞内的跨膜运输,维持细胞的正常生理代谢过程。然而,细胞壁并不能完全阻隔重金属,部分重金属离子仍可进入原生质体。已进入原生质体的金属离子大多会被运送到液泡中,与富硫肽和有机酸等物质结合,贮存于液泡中,减少对细胞器造成的伤害[11-12]。朱光旭等[38]发现,野艾蒿、胜红蓟和野茼蒿3种菊科植物根、茎、叶中镉、铅、锌主要分布于细胞壁和可溶性组分中,在细胞器中分布较少。笔者研究中,镉、铅主要储存于可溶性组分(45.2%~62.5%和44.2%~66.7%)中,细胞壁(29.7%~44.0%和25.2%~44.1%)次之,这表明细胞壁对镉、铅的固持能力有限,进入植物体内的镉、铅主要被转运至液泡中以缓解毒害;锌在马缨丹细胞壁(39.8%~50.4%)和可溶性组分(39.2%~52.4%)中占比较高,且相差不大,表明相较于镉、铅,马缨丹细胞壁对锌的固持作用更强,细胞壁固持和液泡区隔化作用均是马缨丹降低锌毒害的重要机制。细胞器作为植物细胞内代谢最活跃的部位,其重金属含量越多,植物受到的损害越大[15]。笔者试验发现,在复合污染条件下,只有极少数镉、铅、锌分布于细胞器中,分别仅占总量的7.1%~12.2%、6.0%~13.1%和4.8%~11.6%,这与马缨丹在镉、铅、锌单一污染条件下的研究结果[22-23,33]一致,表明在单一或复合污染条件下,区隔化作用都是增强马缨丹耐性、维持其正常生长的重要途径。马缨丹根系及其细胞器中重金属总含量大于地上部,这可能是马缨丹根系在镉、铅、锌复合胁迫下受抑制程度高于地上部的重要原因。
重金属在植物体内的毒性和迁移能力与其存在形态密切相关。乙醇提取态(主要提取无机盐和氨基酸盐)和水提取态(主要提取有机酸盐和水溶性磷酸盐)重金属具有较强迁移能力和较高毒性,氯化钠提取态(主要提取蛋白质和果胶酸盐)和醋酸提取态(主要提取难溶性磷酸盐)次之,盐酸提取态(主要提取草酸盐)和残渣态的迁移能力和毒性最弱[16-17]。笔者研究发现,镉、铅、锌在马缨丹各器官中主要以氯化钠提取态和醋酸提取态形式存在,这与XIN等[39]对荻的研究结果相似,表明重金属在马缨丹体内主要与蛋白质、果胶酸盐和难溶性磷酸盐结合,形成毒性较低的复合物以提高自身耐性。随着重金属处理含量增加,马缨丹茎、叶中醋酸提取态镉,醋酸提取态、盐酸提取态和残渣态铅、锌占比增加,表明马缨丹地上部可通过将重金属转变为毒性较低的形态以减轻镉、铅、锌的毒害。同时,马缨丹茎、叶中乙醇提取态和水提取态锌较同形态镉、铅占比更高,这可能是由于锌是植物生长的必需营养元素之一,在植物体内迁移能力较强,能被转移至生理代谢所需部位,满足植物生长发育需要。
4 结论
(1)马缨丹对镉、铅、锌复合污染具有较好耐性,能够忍耐w(Cd)、w(Pb)和w(Zn)分别为40、600和800 mg·kg-1的复合污染,可用于锌矿区植被恢复和重金属污染土壤治理。
(2)镉、铅、锌复合胁迫下,马缨丹可将大部分重金属,尤其是镉、锌滞留在根系中,限制其向地上部转运以提高自身耐性;同时,马缨丹根系中重金属总量高于地上部,这是地上部受毒害程度低于根系的重要原因。
(3)细胞壁固持和液泡区隔化作用是马缨丹增强自身耐性的重要途径,且相较于锌而言,马缨丹对镉、铅的耐性主要依赖于液泡区隔化作用。同时,通过螯合作用将镉、铅、锌转变为毒性较低的氯化钠提取态和醋酸提取态也是马缨丹重要的耐性和解毒机制之一。
猜你喜欢
快乐语文(2020年26期)2020-10-16 03:37:00
浙江工业大学学报(2017年5期)2018-01-22 02:03:42
浙江工业大学学报(2017年5期)2018-01-22 02:03:40
现代园艺(2017年23期)2018-01-18 06:57:46
广东农业科学(2017年5期)2017-08-29 10:37:54
技术与教育(2014年2期)2014-04-18 09:21:33
食品工业科技(2014年9期)2014-03-11 18:15:40
华东师范大学学报(自然科学版)(2014年3期)2014-03-11 16:18:22
食品科学(2013年24期)2013-03-11 18:30:38
植物营养与肥料学报(2010年4期)2010-11-06 07:28:54
