
快大型黄羽肉鸡肉品质性状的遗传参数估计和关键基因挖掘
2021-09-27 09:03:08杨欣婷郑麦青谭晓冬赵桂苹刘冉冉
畜牧兽医学报 2021年9期
杨欣婷,郑麦青,谭晓冬,赵桂苹,黄 超,李 森,李 韦,文 杰,刘冉冉*
(1.中国农业科学院北京畜牧兽医研究所 动物营养学国家重点实验室 农业农村部动物遗传育种与繁殖(家禽)重点实验室,北京 100193;2.广西金陵农牧集团有限公司,南宁 530049)
随着人民生活质量的提高,消费者对肉品质更加关注。鸡胸肉作为饮食中优质蛋白的重要来源,深受人们喜爱。影响肉品质的主要因素包括遗传基因、营养水平、饲养管理方式和宰前应激。鸡肉品质性状的评价指标包括pH、肉色、系水力、嫩度、肌内脂肪含量、风味物质等。pH作为评价肉品质最重要的指标之一,反映了宰后肌肉糖酵解速率和程度。宰后动物自身的平衡机制被打破,ATP消耗仍在继续,为维持细胞稳态,机体通过糖酵解合成ATP。ATP水解产生H+,导致宰后肌肉pH下降。宰后pH下降的速度和程度显著影响肌肉的系水力[1],当pH降到蛋白质的等电点时,肌原纤维蛋白结合水的能力降低,同时静电斥力减小,肌原纤维粗丝与细丝的间距变小,分布在其中的水分流失,肉在烹调过程中失重[2]。肉色作为评价肉品质又一重要的指标,直接影响消费者购买意愿[3]。肉色常用感官评定或色差计测定,如CIE系统中L*值表示亮度(lightness),a*值表示红度(redness),b*值表示黄度(yellowness)。肉色主要取决于肌红蛋白(myoglobin, Mb)的氧化或还原程度[4](约占70%~80%),刚屠宰后鲜肉的肌红蛋白尚未与氧结合,处于还原状态(Mb),肉呈现暗红色;与空气中的氧接触后,成为氧合肌红蛋白(MbO2),肉呈现鲜红色;随着时间的延长,亚铁血红素中的Fe2+氧化成Fe3+,成为高铁肌红蛋白(MetMb),肉呈现暗褐色[5]。肉色还取决于血红蛋白(haemoglobin, Hb)(约占20%~30%)和微量有色代谢物的组成,如β-胡萝卜素的沉积主要影响肉色b*值[6]。此外,肉色也受到肌肉pH的影响[7],肌肉pH的下降使肌原纤维粗丝与细丝间的水分流失,大量水存在于胞外空间,形成更开放的结构,光的散射增加[8],肉色更苍白(肉色L*值升高)。肉鸡胸肌的肉色L*值介于45到67之间[9]。在实际生产中,pH终值和肉色L*值作为评判生理正常和异常肉质的重要指标,pH终值<5.7且宰后24 h肉色L*值>53的鸡肉,被判定为“类PSE肉”[10]。
目前鉴定到肉品质性状的主效基因有氟烷基因(halothane gene,Hal)、酸肉基因(rendement napole gene,RN)、磷酸化酶激酶调节亚基α1(phosphorylase kinase regulatory subunit alpha 1,PHKG1)和β-胡萝卜素加氧酶1(beta-carotene oxygenase 1,BCO1)等。氟烷基因是由兰尼定受体l基因(skeletal muscle ryanodine receptor,RYR1)的点突变(1843C-T)导致蛋白受体第615位上的Arg突变为Cys造成的[11]。在应激因子作用下,肌浆内钙离子非正常释放,造成肌肉持续收缩,屠宰后肌肉糖降解加速,进而产生PSE肉[12]。酸肉基因由AMP-激活蛋白激酶γ3亚基(protein kinase AMP-activated non-catalytic subunit gamma 3,PRKAG3)的突变密码子(R200Q)引起,突变使AMPK活性降低3倍[13],肌肉中糖原含量提高70%以上[14],导致pH终值过低,引起酸肉效应。PRKAG3的多态性位点和氨基酸替换对肉质都有不同程度的影响,显性错义突变(R225Q)会导致肌肉糖原含量显著增加[15],氨基酸替换T30N和G52S与宰后24 h肉色L*值显著相关[16],而氨基酸替换I199V则会使骨骼肌中糖原含量降低,有利于改善肉质[17]。PHKG1的Intron9剪接受体位点的点突变(8283C-A)导致开放阅读框中的32 bp缺失,终止密码子过早产生,降低PHKG1的mRNA和蛋白表达水平,降低糖原磷酸化酶激酶(phosphorylase kinase,PHK)的酶活性,造成猪肌肉组织中糖原分解速率下降,肌糖原贮积过高和pH终值过低。BCO1启动子的2个SNPs降低了启动子活性和mRNA表达量,增加鸡血浆和胸肌中的叶黄素和玉米黄质含量,鸡胸肌肉色b*值升高[18]。敲除BCO1基因的小鼠器官中也会出现β-胡萝卜素的积累[19]。肉色较浅的鱼也会比肉色较红的鱼具有更高的BCO1表达和蛋白丰度[6]。
除此之外,pH和肉色等相关肉品质性状定位到的QTL有:鸡1号染色体上396 cM的QTL影响鸡胸肌初始pH[20],区域内4个差异表达基因(KLHL15、APOO、PRDX4、ACOT9)都受到调节以保护肌肉细胞免受氧化应激来控制pH,从而影响系水力、滴水损失和肉的加工产量[21]。Bihan-Duval等[22]在6代pH终值上、下选系的肉鸡中共定位到24个QTLs与胸肌pH终值相关,10个QTLs与腿肌pH终值相关。Sun等[23]定位到COL1A2基因上游的SNP与鸡胸肌肉色L*值显著关联,并且COL1A2基因在肉色L*值高低组中差异表达。COL1A2基因在中国眉山猪的红肌和白肌之间也存在表达差异(4倍)[24]。
本研究以快大型黄羽肉鸡终端父系为研究素材,基于“京芯一号”55K SNP芯片基因分型,对胸肌肉品质性状进行遗传参数估计和GWAS分析,以期获得影响黄羽肉鸡肉色和pH性状的基因及显著位点,为肉品质性状遗传选择方案制定和主效基因/SNP鉴定奠定必要基础。
1 材料与方法
1.1 试验动物
试验群体为广西金陵农牧集团有限公司生产饲养的快大型黄羽肉鸡终端父系,来自2个世代共1 923只 鸡,其中公鸡1 013只,母鸡910只。每个世代的鸡同一天孵化,采用阶梯式笼养方式,进行常规免疫,自由采食和饮水,饲料参照NRC(1994)国际标准,所有个体健康状况良好。
1.2 性状测定
56日龄(上市日龄)统一屠宰,并进行屠宰性状、胸肌肉品质性状的测定。屠宰性状包括全净膛重(g)、胸肌重(两侧)(g)、胸肌率(%),测定和计算方法参照中华人民共和国农业行业标准《肉鸡生产性能测定技术规范》(NY/T 828-2004)。肉品质性状包括宰后24 h pH、宰后15 min肉色、宰后24 h肉色,屠宰后剥离同一侧胸肌,使用HI8424便携式pH计(哈纳 HANNA,意大利)测定pH,使用CR-410色彩色差计(柯尼卡美能达公司,日本)测定肉色,测定方法参照中华人民共和国农业行业标准《肉的食用品质客观评价方法》(NY/T 2793-2015)。所有表型值,以“平均值±3倍标准差”进行质量控制。
1.3 基因分型与质控
鸡翅静脉采血后,将血液均匀涂在血液采集卡的点样圈内,然后自然阴凉至完全干燥后,常温送至实验室以磁珠法抽提血液DNA。用琼脂糖凝胶电泳进行检测,采用NanoDrop 2000进行质量评估。合格的基因组DNA利用“京芯一号”鸡55K SNP芯片[25]检测基因型。
使用PLINK(V1.9)软件[26]对SNP进行质量控制,具体过程为:个体基因型检出率≥ 90%,单个SNP检出率≥ 90%,最小等位基因频率≥ 95%,哈代-温伯格平衡检验P≥ 1×10-6,共有1 923只鸡和43 662个SNPs通过质控,用于后续研究。
1.4 遗传参数估计
使用R(V3.6.0)软件ASReml v3.0程序[27]的限制性极大似然法(REML)拟合线性混合模型,估计屠宰性状和肉品质性状的方差组分。使用Wald F检验固定效应,显示性别和世代对所有屠宰性状和肉品质性状的效应均显著(P<0.05)。运用单性状分析法,估算各性状的遗传力,基于系谱的BLUP方法的单性状动物模型:
Y=Xb+Zu+e
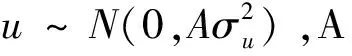
基于基因型数据的GBLUP方法的单性状动物模型:
Y=Xb+Zu+e

运用两性状分析法估算各性状间的遗传相关和表型相关,基于系谱的BLUP方法的两性状动物模型:
其中,y、b、u、e同BLUP方法的单性状动物模型。
基于基因型数据的GBLUP方法的两性状动物模型:
其中,y、b、u、e同GBLUP方法的单性状动物模型。
1.5 全基因组关联分析
使用GEMMA(V0.98.1)软件[28]构建单性状混合线性模型进行GWAS分析。SNP 作为固定因子,个体亲缘关系作为随机效应。统计模型如下:
y=Wα+xβ+u+ε,u~MVNn(0,λτ-1K),ε~MVNn(0,τ-1In)
其中,y是表型值的向量;W是固定效应矩阵,包括性别和世代;α是包含截距在内的相应系数;x是基因型向量;β是SNP的效应向量;u是随机效应;ε是残差。λ是2个方差组分的比率,τ-1是残差的方差,K是SNP估计的中心亲缘关系矩阵;ε是残差向量,In是单位矩阵。
利用Wald检验法确定SNP的显著性。利用Bonferroni校正多重检验确定显著性阈值,全基因组显著水平阈值为1.15×10-6(0.05/43 662),全基因组水平潜在显著阈值为2.29×10-5(1/43 662)。使用R(V3.6.0)软件qqman包,实现GWAS结果的可视化。
SNP 解释性状遗传变异的计算公式:
1.6 连锁不平衡分析
使用Haploview软件[29]的“Solid spine of LD”算法对显著SNPs进行连锁不平衡分析,构建单倍型区块。
1.7 基因注释
使用Ensembl、UCSC、Genecards数据库,查询显著SNPs所在位置的候选基因,进行基因功能注释。
2 结 果
2.1 性状描述性统计
本研究对1 923只黄羽肉鸡的屠宰性状和胸肌肉品质性状进行表型测定,包括全净膛重、胸肌重、胸肌率、宰后15 min肉色L*值(L*15 min)、宰后15 min肉色a*值(a*15 min)、宰后15 min肉色b*值(b*15 min)、宰后24 h肉色L*值(L*24 h)、宰后24 h肉色a*值(a*24 h)、宰后24 h肉色b*值(b*24 h)、宰后24 h pH(pH24 h)。描述性统计分析结果见表1。其中,全净膛重、胸肌重、a*15 min、b*15 min、a*24 h、b*24 h的变异系数较高,介于13.78%到20.98%,具有较大的遗传改良潜力。
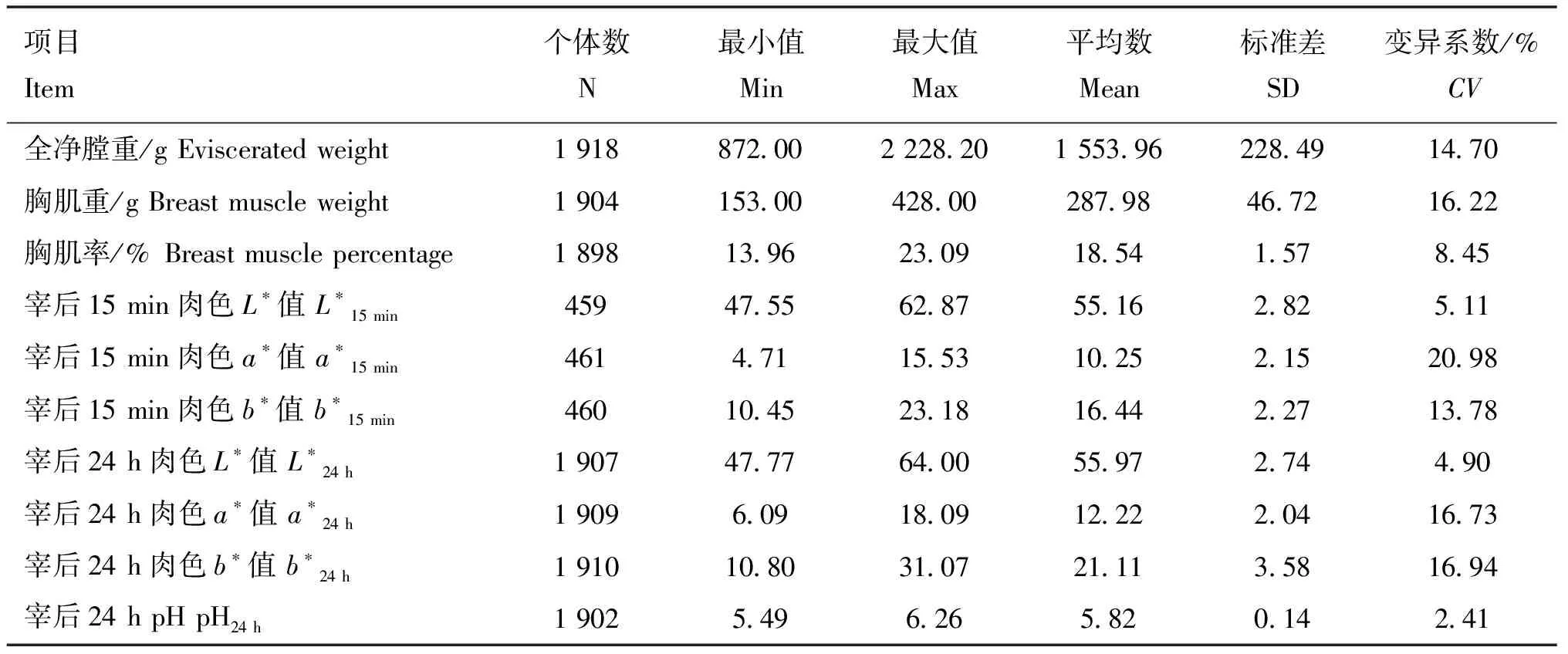
表1 屠宰性状和胸肌肉品质性状的描述性统计
2.2 遗传参数估计
基于BLUP方法估计的屠宰性状和胸肌肉品质性状的遗传参数见表2,基于GBLUP方法估计的屠宰性状和胸肌肉品质性状的遗传参数见表3。基于BLUP方法估计L*15 min、胸肌率的遗传力分别为0.47、0.50,属于高遗传力;b*15 min、L*24 h、b*24 h、pH24 h、胸肌重的遗传力分别为0.21、0.38、0.34、0.38、0.40,属于中等遗传力;a*15 min、a*24 h的遗传力分别为0.10、0.17,属于低遗传力。基于GBLUP方法估计L*15 min、L*24 h、b*24 h、pH24 h、胸肌重、胸肌率的遗传力分别为0.25、0.29、0.32、0.29、0.31、0.40,属于中等遗传力;b*15 min、a*24 h的遗传力分别为0.19、0.17,属于低遗传力。两种方法除了a*24 h的遗传力估计值一致外,BLUP方法估计的遗传力均要大于GBLUP方法估计的遗传力。表型相关方面,两种方法的估计值较一致,L*15 min与L*24 h、b*15 min与b*24 h、胸肌重与胸肌率为较高的正相关(0.52~0.70),L*24 h与a*24 h、L*24 h与pH24 h为较高的负相关(-0.39~-0.48)。遗传相关方面,两种方法的估计值较一致,L*15 min与L*24 h、a*15 min与a*24 h、b*15 min与b*24 h、胸肌重与胸肌率为较高的正相关(0.63~0.99),L*24 h与a*24 h、L*24 h与pH24 h、L*15 min与胸肌率为较高的负相关(-0.47~-0.76)。

表2 基于BLUP方法估计的屠宰和胸肌肉品质性状的遗传力(对角线)、表型相关性(对角线以下)、遗传相关性(对角线以上)及其标准误

表3 基于GBLUP方法估计的屠宰和胸肌肉品质性状的遗传力(对角线)、表型相关性(对角线以下)、遗传相关性(对角线以上)及其标准误
2.3 全基因组关联分析
L*15 min、L*24 h、b*24 h和pH24 h的显著SNPs信息见表4,全基因组关联分析的曼哈顿图和Q-Q图见图1。对于L*15 min,1个SNP(rs15291911)达到全基因组水平潜在显著阈值,位于3号染色体上,在NSL1基因内部。对于L*24 h,9个SNPs达到全基因组水平潜在显著阈值,位于5号染色体上,构成1个 575.63 kb的显著区间(Chr5: 48.49-49.06 Mb),其中3个显著SNPs(rs16506880、rs14545229、rs14545237)位于EVL基因内部,3个显著SNPs分别位于SLC25A47(rs14545421)、WARS(rs317845442)、BEGAIN(rs312473490)基因内部,1个显著SNPs(rs312331816)位于DLK1基因下游895 bp处。对于b*24 h,7个SNPs达到全基因组水平潜在显著阈值,2个显著SNPs位于5号染色体上,其中rs14528290位于ZFYVE26基因内部,rs312473490位于BEGAIN基因内部;5个显著SNPs位于11号染色体上,其中rs313386167和rs312270923位于CDYL2基因内部,rs14966310和rs15621925位于BCO1基因内部。对于pH24 h,1个 SNP(rs313754724)达到全基因组水平潜在显著阈值,位于28号染色体上,在ARHGEF18基因内部。

X轴的1~28表示1~28号染色体,29代表Z染色体;图中粗实线代表全基因组水平潜在显著阈值(2.29×10-5),细实线代表全基因组显著水平的阈值(1.15×10-6)
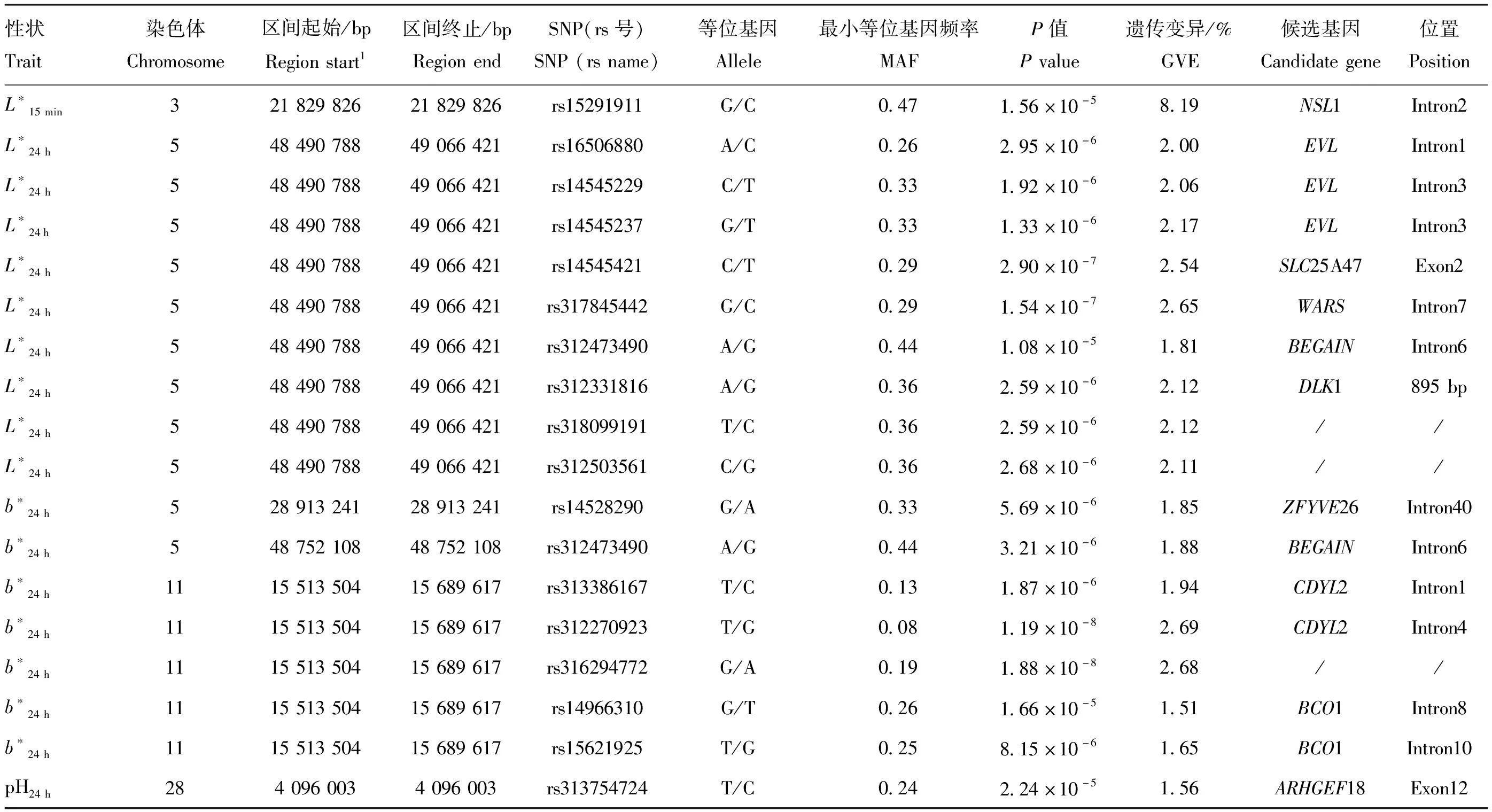
表4 胸肌肉品质性状的显著关联SNPs信息
2.4 连锁不平衡分析
L*24 h显著区间(Chr5: 48.49~49.07 Mb)和b*24 h显著区间(Chr11: 15.51~15.69 Mb)上的显著SNPs的连锁不平衡分析结果见图2。方格中的数字为D’,表示连锁不平衡的程度;方格颜色越深,表明SNPs之间的连锁不平衡程度越强。单倍型区块由软件自动生成,1个单倍型区块内的SNP完全连锁。L*24 h显著区间(Chr5: 48.49~49.07 Mb)上存在2个单倍型区块,大小分别为24 和427 kb;b*24 h显著区间(Chr11: 15.51~15.69 Mb)上存在2个单倍型区块,大小分别为57 和2 kb。

A.位于5号染色体上的2个单倍型区块;B.位于11号染色体上的2个单倍型区块
2.5 单倍型的效应分析
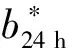

表5 单倍型对胸肌肉品质性状的影响
3 讨 论
3.1 遗传参数估计
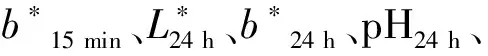
本研究中,BLUP和GBLUP方法估计的表型相关较一致,L*15 min与L*24 h、b*15 min与b*24 h、胸肌重与胸肌率为较高的正相关(0.52~0.70),L*24 h与a*24 h、L*24 h与pH24 h为较高的负相关(-0.39~-0.48)。BLUP方法和GBLUP方法估计的遗传相关较一致,L*15 min与L*24 h、a*15 min与a*24 h、b*15 min与b*24 h、胸肌重与胸肌率为较高的正相关(0.63至0.99),L*24 h与a*24 h、L*24 h与pH24 h、L*15 min与胸肌率为较高的负相关(-0.47~-0.76)。先前研究中,6495只肉鸡的L*24 h与a*24 h(-0.57)、pH24 h与L*24 h(-0.79)、pH24 h与b*24 h(-0.33)、b*24 h与a*24 h(-0.23)均为负表型相关,L*24 h与b*24 h(0.48)、pH24 h与a*24 h(0.52)均为正表型相关[35];1 022只 肉鸡的L*24 h与a*24 h(-0.28)、pH24 h与L*24 h(-0.83)、pH24 h与b*24 h(-0.53)均为负遗传相关;L*24 h与b*24 h(0.51)、pH24 h与a*24 h(0.15)均为正遗传相关[33]。以上研究与本研究估计的表型相关和遗传相关结果较一致。
3.2 全基因组关联分析
目前已有研究通过GWAS定位到影响pH和肉色等肉品质性状的候选基因。1 768头“三元杂”猪的pH终值定位到候选基因GLUL[36]。165头杜洛克母猪的初始pH和pH终值定位到候选基因PKHD1L1、VCPIP1和LOC102166532[37]。150头苏太猪的背最长肌和半膜肌pH和肉色定位到候选基因BNIP3、PRKG1和ADRB3[38]。刘大鹏等[39]结合转录组测序,定位到555只北京鸭×野鸭F2代资源群体的胸肌肉色b*值的候选基因SELENOT。
在本研究中,L*15 min定位到候选基因NSL1,该基因编码的蛋白质具有两个位于着丝粒上的卷曲螺旋域,在细胞分裂期间与微管连接,并调节染色体运动(https://www.genecards.org/)。L*24 h定位到的候选基因有BEGAIN、DLK1、EVL、SLC25A47、WARS。BEGAIN编码的蛋白质维持突触后密度的结构,与突触的蛋白质-蛋白质相互作用和化学突触间的传递相关。DLK1编码肌肉发育的促进因子[40]和脂肪发育的抑制因子[41],是体内成肌细胞正常分化和肌肉再生必需的[42],并在鸡脂肪和肌肉发育中起作用[43]。EVL编码肌动蛋白相关蛋白,增强肌动蛋白成核和聚合,参与一系列依赖于细胞骨架重塑和细胞极性的过程[44]。WARS基因上的His-257-Arg突变会降低细胞活力,抑制神经突生长,导致肢体肌肉无力和萎缩的运动神经病[45]。值得一提的是,SLC25A47属于溶质载体家族成员,该家族有众多基因均影响pH和肉色等肉品质性状。SLC24A5在欧洲和南亚人群的浅肤色中发挥作用[46];SLC2A1在6代pH终值上、下选系的肉鸡中差异表达[22];SLC3A2是猪肉滴水损失的候选基因[47];SLC37A4的突变会导致某些器官和组织的糖原贮积病[48]。b*24 h定位到的候选基因有BCO1、BEGAIN、CDYL2和ZFYVE26。其中,ZFYVE26突变会导致遗传痉挛性截瘫,表现为肌萎缩、轴突神经病、阵挛[49]。BCO1是鸡胸肉色b*值的主效基因,在动物体内类胡萝卜素的代谢过程中发挥重要作用,对鸡胸肉色[18]、家禽皮肤颜色[50]、鱼肉色[6]、牛肉脂肪颜色[51]、人类黄皮病[52]均有影响。pH24 h定位到候选基因ARHGEF18,该基因编码调节多种细胞功能的GTP结合蛋白,诱导肌动蛋白应力纤维的形成。20日龄鸡血液pH同样定位到ARHGEF18,该基因可能是影响鸡胸肉色pH24 h的重要候选基因[53]。
3.3 单倍型的效应分析
单倍型效应分析显示,5号染色体上Block1为CCCCGG型的个体具有更低的L*15 min、L*24 h、b*24 h和更高的pH24 h,5号染色体上Block2为CCGGAAAATTCC型的个体具有更低的L*24 h、b*24 h和更高的pH24 h,说明这2个单倍型对鸡的pH和肉色性状具有一因多效作用,并且是鸡肉品质性状的有利单倍型,可以尝试在实际育种中加以选择利用。
4 结 论
本试验对快大型黄羽肉鸡的肉品质性状进行了遗传参数估计和全基因组关联分析,胸肌pH、肉色L*值和b*值为中等遗传力性状(0.21~0.38),适合进行遗传改良;pH和肉色性状的全基因组关联分析,共筛选到18个达到全基因组水平潜在显著阈值的SNPs和5个达到全基因组显著水平的SNPs,分别位于3、5(Chr5: 48.49~49.06 Mb)、11(Chr11: 15.51~15.69 Mb)和28号染色体上,候选基因包括ARHGEF18、BCO1、BEGAIN、CDYL2、DLK1、EVL、NSL1、SLC25A47、WARS、ZFYVE26。其中,BCO1是已鉴定的鸡胸肌肉色b*值的主效基因;SLC25A47属于溶质载体家族成员,影响L*24 h,该家族有众多基因均影响pH和肉色等肉品质性状。此外,位于5号染色体上的2个单倍型对胸肌pH、肉色性状均有极显著影响。这些结果为黄羽肉鸡肉品质性状的遗传改良和分子机制的深入研究奠定了重要基础。
猜你喜欢
国外畜牧学·猪与禽(2024年4期)2024-01-01 00:00:00
花卉(2023年21期)2023-11-16 07:54:00
食品科学(2022年17期)2022-09-30 07:49:12
养猪(2022年4期)2022-08-17 07:07:02
食品工业科技(2018年20期)2018-10-24 01:09:28
幼儿教育·父母孩子版(2018年12期)2018-04-20 11:35:52
恋爱婚姻家庭(2017年18期)2017-12-26 07:20:42
黑龙江八一农垦大学学报(2017年1期)2017-03-10 01:46:42
广东饲料(2016年7期)2016-12-01 03:43:36
湖北畜牧兽医(2015年11期)2016-01-11 10:08:24
