植物内生放线菌多样性及其活性代谢产物的研究进展
2021-09-24 00:28:50刘文杰张新国
微生物学杂志 2021年4期
封 明, 刘文杰, 苏 誉, 王 楠, 毛 瑜, 张新国* , 张 继
(1.兰州理工大学生命科学与工程学院 甘肃省中藏药筛选评价及深加工重点实验室,甘肃 兰州 730050;2.西北师范大学 生命科学学院,甘肃 兰州 730050)
自20世纪40年代青霉素和链霉素相继被发现以来,研究人员从不同的微生物,特别是放线菌中发现了大量具有生物活性的代谢产物[1]。研究显示,已经有超过30 000多种天然产物从微生物中被分离鉴定,这其中有超过50%的活性物质(如抗生素)源自于放线菌(70%为链霉菌属)的代谢产物。放线菌无疑是天然活性物质极为丰富的重要微生物资源[2]。然而,随着放线菌分离鉴定方法的成熟及其广泛应用,从自然环境中获取新的放线菌正在变得越来越困难[3]。 植物内生放线菌是植物内生菌中重要的一员,是存在于植物各组织内而且不会引起宿主植物产生明显感染症状,并与植物长期共存的一类微生物[4]。研究显示,植物内生放线菌不仅具有促进宿主植物生长繁殖的作用,而且能产生与宿主植物相同或相似的活性代谢产物。植物内生放线菌作为一种远未完全开发的微生物资源,对其进行更进一步的探索意义重大[5]。本文介绍了近年来国内外关于植物内生放线菌的研究现状,并总结了一些重要的研究进展。
1 植物内生放线菌的多样性
植物内生放线菌广泛存在于各种植物中,在长期协同进化过程中与宿主植物形成了互利共生的关系[6]。最早被发现的内生放线菌是能与各种非豆科植物形成根瘤并具有固氮作用的弗兰克氏菌属(Frankiasp.)[7]。此后,研究人员陆续在不同植物包括藻类[8]、苔藓类[9]、蕨类[10]、地衣[11]、裸子植物[12]和被子植物[13]等中相继发现了不同种类的内生放线菌。越来越多的研究显示,在已有研究的不同种类的植物中,都有植物内生放线菌存在[14]。
内生放线菌不仅广泛分布于各类植物,而且在植物的不同组织部位如根、茎、叶等均有分布。研究证实,不同植物内生放线菌的多样性和群落结构在植物不同组织部位的分布存在着显著差异,其中以植物的根部组织分布为多[15]。Passari等[16]从7种药用植物的不同部位中分离到42株内生放线菌,其中从根中分离到的放线菌最多(22株,52.4%),其次是茎(9株,21.4%)、叶(6株,14.3%)、花(3株,7.1%)和叶柄(2株,4.8%)。Gangwar等[17]对芦荟、薄荷和圣罗勒3种药用植物的内生放线菌多样性进行了研究,发现内生放线菌最多见于根(占所有菌株的70%),其次是茎(17.5%)和叶(12.5%)。Verma等[18]从印楝中分离出55株内生放线菌,大多数菌株(30株,54.54%)来自根,其次是茎(13株,23.6%)和叶(12株,21.8%)。Gangwar等[19]从印度醋栗中共分离到36株内生放线菌,有18株(50%)分布在根部,茎和叶均分离出9株放线菌,各占25%。但是也有研究显示,根以外的组织也是内生放线菌存在的主要部位。Kampapongsa等[20]从3种水稻样品中分离获得116株内生放线菌,对植物不同部位放线菌的分析表明,从叶鞘中分离出的放线菌最多,有47株(40.5%),从茎、叶和根中分别获得34(29.3%)、18(15.5%)和17(14.7%)株放线菌。Vu等[21]从越南Hoa Binh省药用植物肉桂的根、茎和叶中分离出111种放线菌,从茎中分离出的放线菌最多,有67株占60.4%,其次从根部分离出29株,叶子中分离出的内生放线菌最少,只有15株。
植物内生放线菌除了具有组织分布多样性还具有明显的地理差异性,研究表明不同地域的同一种植物其内生放线菌种类和数量均有所不同。陈萌等[22]从四川省不同地区的川楝中共分离得到403株内生放线菌,从广元地区采集的样品中分离得到的内生放线菌数量最多,有86株;而绵阳地区最少,只分离到12株。Musa等[23]从新疆伊犁和塔城两个干旱地区的中草药百里香中分离到126株内生放线菌,其中,塔城地区可培养的内生放线菌属的多样性(71株,56.3%)高于伊犁州(55株,43.7%)。姜龙芊等[24]从西双版纳、白茫雪山、波罗的海南岸地衣中共分离到1 123株内生菌,鉴定菌株417株,从西双版纳地衣样品分离出放线菌107株,从白茫雪山样品中分离到103株放线菌,从波罗海南岸地衣样品分离出的内生放线菌最少,仅有65株。
广泛分布的植物内生放线菌种类庞杂,不仅存在明显的分布差异,而且具有丰富的种属多样性,随着研究的深入,许多不同种类的放线菌群从不同的植物中被陆续分离[25]。罗红丽等[26]从植物百部块根中分离出18株内生放线菌,经16S rRNA序列分析将其分为链霉菌属(Streptomycessp.)、小单孢菌属(Micromonosporasp.)、假诺卡氏菌属(Pseudonocardiasp.)和甲基杆菌属(Methylobacterium)。Li等[27]从植物黄花蒿中共分离出228株植物内生放线菌,对其中119株菌进行16S rRNA基因测序,其主要分类结果包括糖霉菌属(Glycomycessp.)、野野村氏菌属(Nonomuraeasp.)和原小单孢菌属(Promicromonosporasp.)等19个属。Shan等[28]对茶树中放线菌的丰度和多样性进行系统的研究,从中国福建省15种茶树中共分离出46种放线菌,根据16S rRNA序列分析,将这些菌株分为链霉菌属、马杜拉放线菌属(Madurasp.)和韩国生工菌属(Kribbellasp.)等13个属。Kuncharoen等[29]从泰国的14种植物中分离出37种内生放线菌,根据其表型特征和16S rRNA基因序列,将其分为小单孢菌属21株、链霉菌属14株、植物生孢菌属1株(Plantactinosporasp.)和多形孢菌属1株(Polymorphosporasp.)。穆珊[30]从健康及感染黄萎病的番茄植株中分离获得了140株放线菌,其中链霉菌属109株,占77.9%;野野村氏菌属15株,占10.7%;诺卡氏菌属(Nocardiasp.)和小单孢菌属4株,均占2.9%;糖霉菌属和红球菌属(Rhodococcussp.)3株,均占2.1%,各有3株;马杜拉放线菌属仅2株,占1.4%。植物内生放线菌多样性见表1。
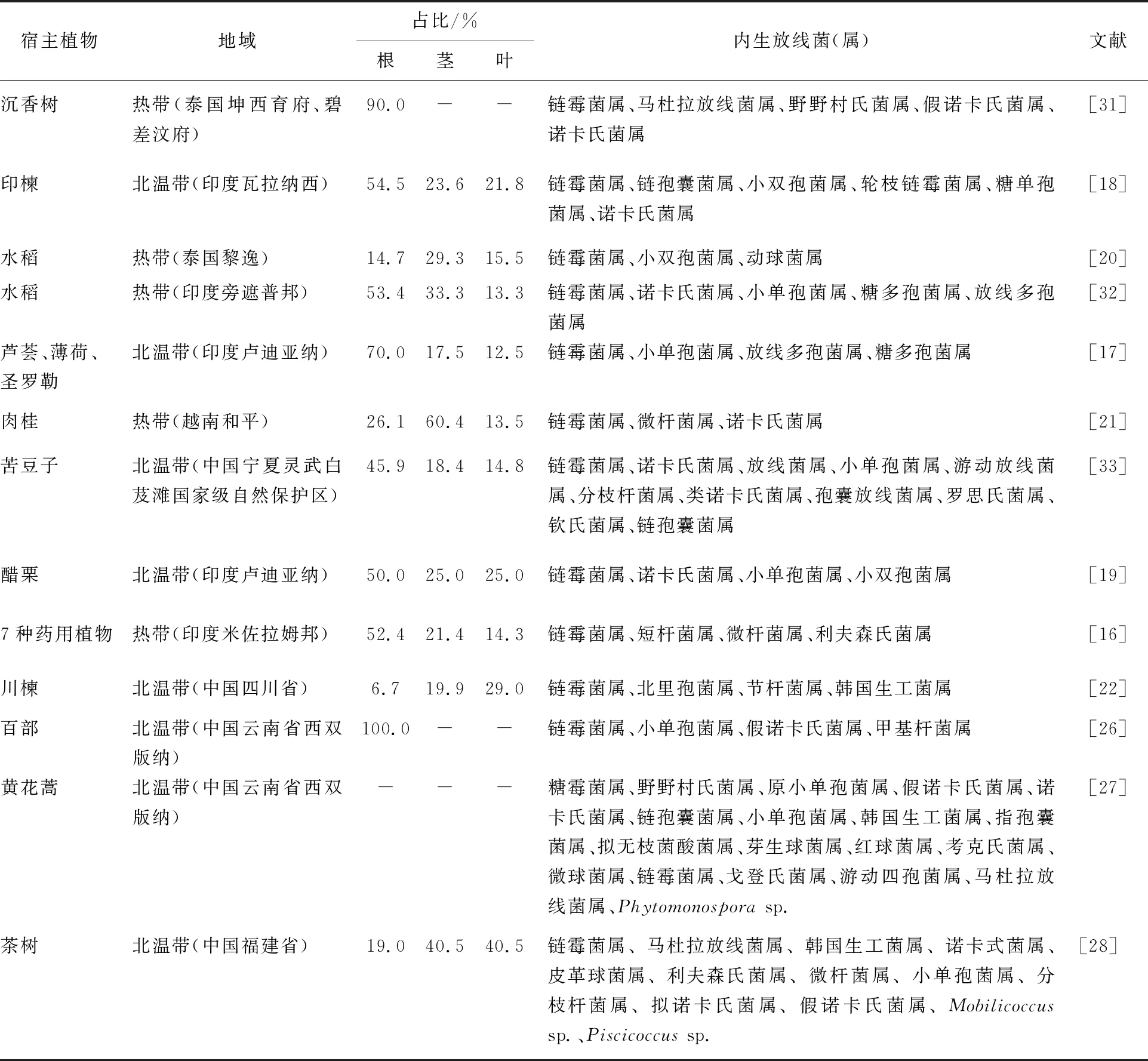
表1 植物内生放线菌的多样性
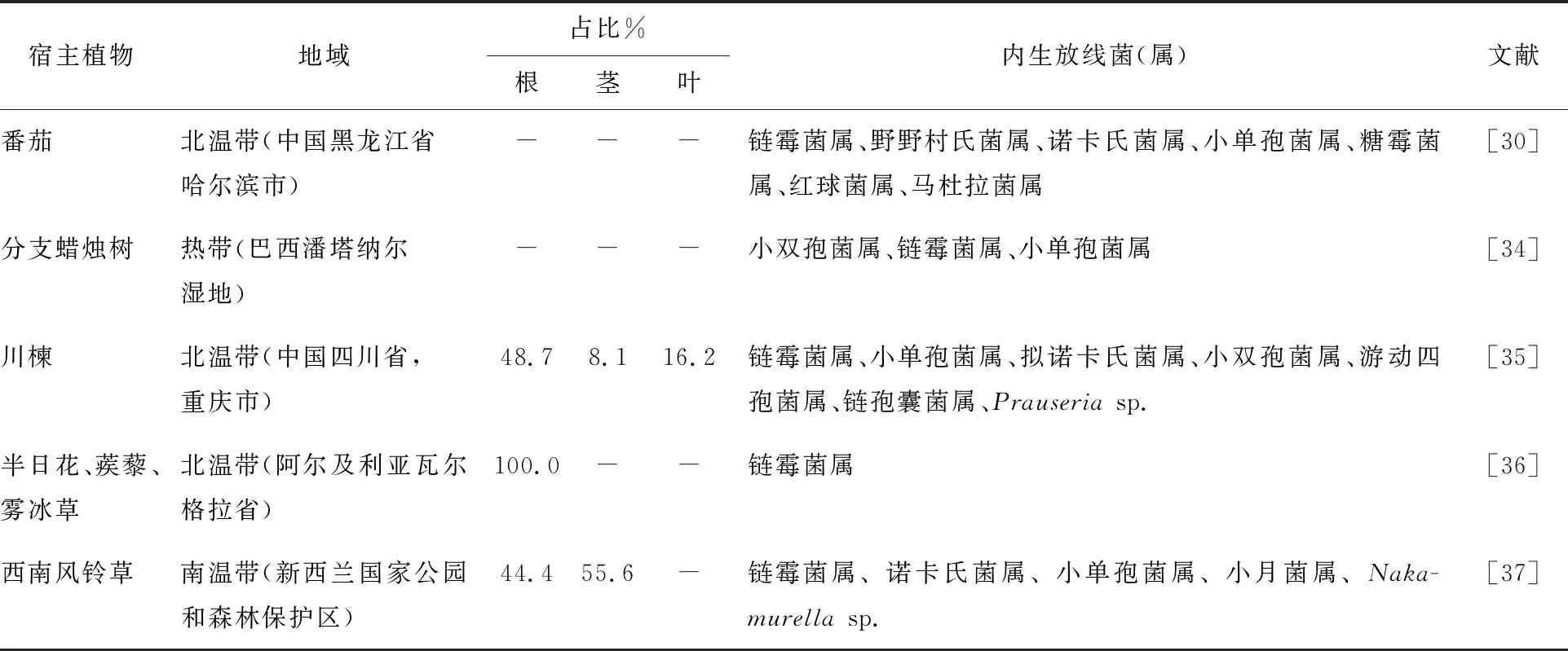
续表1
2 植物内生放线菌活性代谢产物的研究
内生放线菌作为一种亟待开发的重要微生物资源,绝大部分可产生与宿主植物相同或相似的次生代谢产物,是寻找新颖结构候选化合物的重要资源。随着越来越多活性次生代谢产物的发现,对内生放线菌挖掘性研究正引起研究人员的广泛关注[38]。本文就最近几年有关植物内生放线菌活性研究代谢产物研究进行汇总,见表2。
2.1 抗菌活性
放线菌作为重要的抗菌化合物的来源[2],研究人员从内生放线菌中分离获得了许多具有潜在应用价值的抗菌活性物质。Ameen等[39]从长叶薄荷(Menthalongifolia)、小花锦葵(Malvaparviflora)和无头公乌(Pulicariaundulata)3种植物中均分离出了具有抑菌活性的内生放线菌,其中2株小单孢菌(Micromonosporasp.)和1株放线杆菌(Actinobacteriabacterium)发酵液乙酸乙酯提取物对金黄色葡萄球菌(Staphylococcusaureus)、铜绿假单胞菌(Pseudomonasaeruginosa)、大肠埃希菌(Escherichiacoli)的最(MIC)小抑菌浓度在80~160 μg/mL之间。魏少鹏等[40]从药用植物小檗中分离出1株具有抗菌活性的内生放线菌Streptomycessp. H21,并从中分离出广谱抗生素——全霉素,其对蜡状芽胞杆菌(Bacilluscereus)、枯草芽胞杆(Bacillussubtilis)、大肠埃希菌和金黄色葡萄球菌的MIC值分别为0.078、0.313、0.156和0.313 μg/mL。Bunbamrung等[41]从绿萝根组织中内生放线菌Streptomycessp. TBRC7642中分离出的化合物Sch 68631(murayaquinone)对革兰阳性菌蜡状芽胞杆菌和革兰阴性菌大肠埃希菌、鲍曼不动杆菌(Acinetobacterbaumannii)均具有抗菌活性,MIC值分别为3.13、25.00和6.25 μg/mL,显示了进一步研究开发的潜力。
随着抗生素的不合理应用,细菌耐药性正成为日益严峻的问题,据WTO资料统计,每年因耐药菌感染而死亡的人数超过70多万[42]。耐甲氧西林金黄色葡萄球菌(Methicillin-resistantStaphylococcusaureus,MRSA)是导致大规模医疗感染并广泛存在于动物源性食品中,严重危害人类健康的耐药菌[43]。席楠等[44]从刺五加内生放线菌StreptomycestauricusCWJ-107中分离出化合物cinerubin B,对MRSA具有良好的抗菌活性,MIC值为1 μg/mL。Gos等[45]从巴西药用植物Vochysiadivergens中分离出10种内生放线菌,菌株AeromicrobiumpontiLGMB491发酵液乙酸乙酯提取物对MRSA的抑制活性最好,MIC值为0.04 mg/mL。Junaidah等[46]对巴西药用植物中的内生放线菌抑菌活性进行研究,其中生姜中分离出的内生放线菌Streptomycessp. SUK25显示出对MRSA的抑菌活性,MIC值为2.44 μg/mL。Vu等[47]从越南Yen Bai省肉桂中分离出内生放线菌StreptomycescavourensisYBQ59并从中分离出8个化合物,其中化合物1-monolinolein抗菌活性最高,对MRSA(ATCC 33591)和耐甲氧西林表皮葡萄球菌ATCC 35984(MRSE)MIC值分别为8.5和14.6 μg/mL。溶血葡萄球菌(Staphylococcushaemolyticus)和头状葡萄球菌(Staphylococcuscapitis)是临床较为常见的引起化脓性感染的多重耐药菌[48]。Sabu等[49]的研究显示,万古霉素、环丙沙星和庆大霉素对临床分离的多重耐药株S.capitis267和S.haemolyticus41抑制范围分别为17、29、21 mm和18、18、24 mm,从生姜中分离出的内生放线菌Nocardiopsissp. ZoA1发酵液乙酸乙酯提取物对上述两株耐药菌的抑制范围分别为25和28 mm,显示了较好的抑菌潜力。这些研究将为进一步从内生放线菌中发现抗耐药菌药物指明方向。
此外,研究显示许多植物内生放线菌的代谢产物还具有抑制植物病原菌的生物活性。苹果树腐烂病菌(Valsamali)是一种能引起苹果树腐烂病且分布广泛的病原真菌。曹艳等[50]从冬青卫矛内生放线菌中分离得到具有抑制苹果树腐烂病菌的内生放线菌,其中StreptomycesflavofuscusG1菌株中分离出的化合物4′-deacetyl-(-)-griseusin A抑制苹果树腐烂病菌的IC50值为14.70 μg/mL,与阳性对照嘧菌酯(azoxystrobin)的IC50值2.91 μg/mL相比,表现出良好的抗菌活性潜力。Wang等[51]从黄瓜根部分离出具有成为苹果树腐烂病生防剂潜力的内生放线菌SaccharothrixyanglingensisHhs.0.15,进一步从中分离出化合物Fungichromin和1′-deoxyfungichromin,EC50值分别为5.24和4.92 μg/mL,其中化合物Fungichromin以 1 mg/mL连续处理感染苹果树腐烂病菌的试验枝条7 d后,保护率达到81.48%。Sun等[4]从丹参分离出的内生链霉菌PKU-EA00015中,分出7个新化合物strepimidazoles A~G,其中化合物strepimidazoles B和C对黄萎病菌V991(VerticilliumdahliaeV991)表现出最好的抑制作用,两个化合物抑制作用分别为77%和74%,与纽莫康定B0的抑制活性相当。Supong等[52]对分离自水稻(OryzasativaL.)的内生放线菌Streptomycessp. BCC72023进行研究,从中分离出5个化合物,所有化合物对恶性疟原虫K-1菌株(具有多重耐药性)的IC50值在1.40~5.23 μg/mL范围内。
2.2 抗肿瘤活性
目前,肿瘤仍然是我国因病死亡人数最多的疾病,每年死亡病例高达281万例。因此,寻找抗肿瘤新药仍然具有非常重要的意义。尽管基于植物内生放线菌抗肿瘤活性的研究还相对较少,但是科研人员仍然获得了很多令人欣喜的成果。Qiu等[53]从四川省13种传统药用植物中共分离出119种内生放线菌,对其中80株菌进行抗肿瘤活性筛选研究,结果发现87.5%的内生放线菌具有抑制人肝癌细胞HepG2的活性。王新位等[54]从刺五加内生放线菌Streptomycessp. CWJ-256中成功分离得到化合物efomycin G和elaiophylin,进一步研究显示两者对体外培养的人乳腺癌细胞MDA-MB-231均有增殖抑制效果,IC50值分别为4.385和2.118 μmol/L。Zhang等[55]从健康大豆根部内生放线菌Streptomycessp. neau-D50分离出的化合物3-acetonylidene-7-prenylindolin-2-one和7-isoprenylindole-3-carboxylic acid对人肺腺癌细胞系A549具有细胞毒活性,IC50值分别为3.3和5.1 μg/mL,与阳性对照阿霉素(4.2 μg/mL)相当。Igarashi等[56]从豆科植物根瘤中的内生放线菌Micromonosporalupini分离到一种新的蒽醌衍生物lupinacidin C,对小鼠结肠癌细胞有强烈的抑制作用,IC50值为0.019 μg/mL。Conti等[57]从巴西药用植物LychnophoraericoidesMart.的内生放线菌Streptomycessp. RLe8中分离出化合物2,3-dihydro-2,2-dimethyl-4(1H)-quinazolinone,对结肠癌细胞系HCT-8、胶质母细胞瘤细胞SF-295、黑素瘤细胞MDA-MB435和早幼粒细胞白血病细胞系HL-60均有强烈的细胞毒活性,IC50值分别为1.10、2.20、1.86和1.76 μg/mL。
2.3 抗虫活性
研究发现很多植物内生放线菌的次级代谢产物中含有抗虫活性物质。陶玲等[58]从红树林植物木榄内生放线菌 (Streptomycesalbidoflavus)发酵液的乙酸乙酯提取物中分离出不饱和脂肪酸-5,8-二烯十四酸,其对秀丽隐杆线虫的LC50值为162.8 μg/mL,远高于杀虫剂甲胺磷的LC50值(3 282.46 μg/mL),显示出较强的抗虫活性。Zhao等[59]从传统中药直立百部内生菌寻找新的杀虫次生代谢产物的过程中,从内生放线菌Streptomycessp. BS-1中分离鉴定出10种具有显著杀虫活性的新型吡咯羧酸酯类化合物AJ(1~10),所有化合物均对棉蚜虫表现出了很强的致死性(72 h的LC50值为3.55~32.00 μg/mL)。Chen等[60]对印楝树中分离出的85株放线菌进行杀虫活性筛选,结果显示有8株放线菌粗提物对桃蚜的杀虫活性最高(60%以上),其中放线菌Streptomycessp. G30发酵液乙酸乙酯提取物对桃蚜的毒力最强,48 h的LC50和LC95值分别为1.68 mg/mL和4.37 mg/mL。Ezra[61]从附生藤本植物Monsterasp.的内生放线菌Streptomycessp. (MSU-2110)中分离出一种新型多肽类化合物Coronamycins,该化合物对疟原虫具有较强抑制活性,IC50值为9.0 ng/mL。从药用植物内生放线菌中发现具有显著活性的农药天然产物,将为害虫防治的发展开辟新的途径。
2.4 抗氧化活性的次级代谢物
预防衰老的重要途径之一是抗氧化,若能消除过多的氧化自由基对于预防因自身自由基引起的老化相关疾病意义重大[62],因此,以植物内生放线菌为资源进行抗氧化活性研究正备受关注。Singh等[63]从植物曼陀罗中分离出一种新的内生放线菌StreptomycescalifornicusADR1,其次级代谢产物具有良好的抗氧化性能,对1,1-二苯基-2-三硝基苯肼(DPPH)的抑制浓度(IC90)值为(217.24±6.77) μg/mL。凯迪亚·阿亚提等[64]从地锦草中分离出的13株内生放线菌中有3株菌的发酵液乙酸乙酯提取物显示出抗氧化活性,其中内生放线菌MicrobacteriumoleivoransL57发酵液提取物在0.2 mg/mL时,对DPPH自由基清除率超过50%,内生放线菌G4和G26的发酵液提取物对羟基自由基清除率超过50%。Wang等[65]对虎杖及其内生链霉菌A0916提取物的抗氧化活性进行比较,当阳性药抗坏血酸在128 μg/mL、DPPH抑制率为93.8%时,链霉菌A0916提取物和虎杖提取物抑制率分别为93.2%和92.7%。结果表明,链霉菌A0916提取物和虎杖提取物均具有较强的抗氧化活性。
2.5 其他生物活性
除抗菌、抗肿瘤、抗虫和抗氧化活性外,植物内生放线菌次级代谢产物的其他生物活性研究也在逐渐增加[66]。Kuncharoen等[67]从芭蕉根部分离出1株内生放线菌Micromonosporamusaesp. nov.,该放线菌能生产维生素——甲基萘醌。刘敏等[68]从海漆叶部分离出1株具有抗H1N1病毒的内生放线菌T.tyrosinosolvem,其稀释20倍的发酵液对H1N1抑制率为76.5%,与阳性对照利巴韦林(50 μg/mL)80%的抑制率相比,显示出较强的抗病毒活性。Merzaeva等[69]从冬小麦中分离出能产吲哚乙酸(IAA)的内生放线菌Streptomycessp.,可促进冬小麦的生长发育。Goudjal等[70]从5种植物中分离到27株内生放线菌并进行体外产IAA能力的筛选,有18株菌株能产生IAA,其中链霉菌PT2的IAA产量最高,对促进番茄种子萌发和根系伸长方面表现出显著效果。Hassan等[71]从植物酢浆草(Oxaliscorniculata)的叶子中分离出的两种内生放线菌Streptomycessp. Oc-5和Acv-11具有介导纳米氧化铜颗粒合成的作用。内生放线菌作为寻找天然活性代谢产物的重要资源正在变得越来越引人注目。
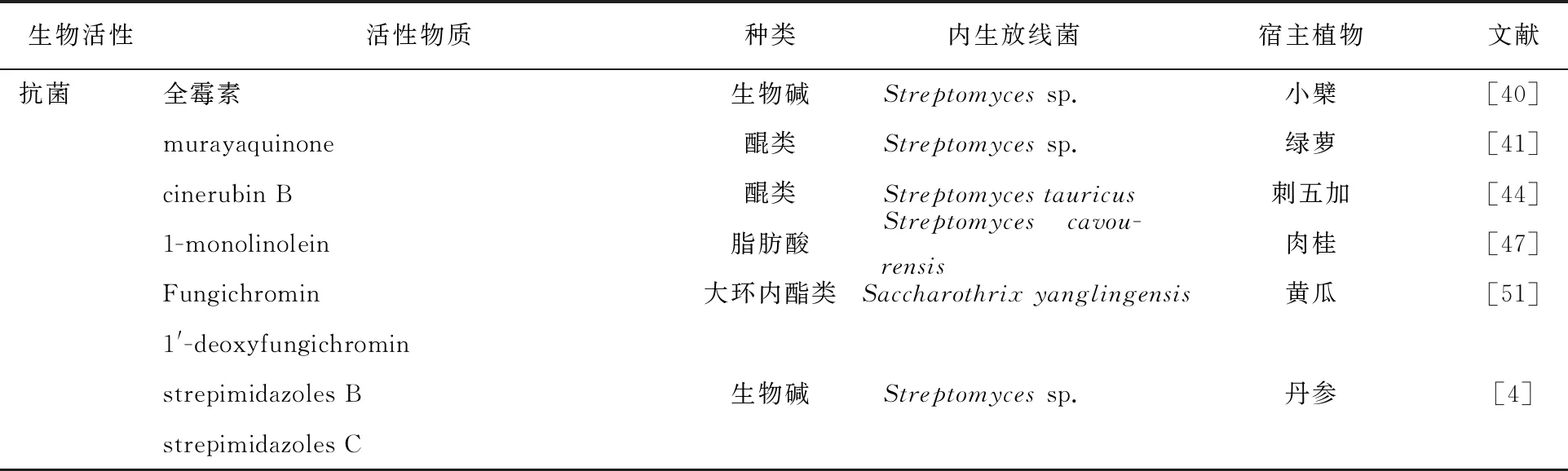
表2 植物内生放线菌活性代谢产物研究

续表2
3 植物内生放线菌的应用与展望
植物内生放线菌具有丰富的生物多样性和代谢产物生物活性,在抗菌、抗肿瘤、抗虫和抗氧化等方面显示出极具潜力的开发前景,正在成为发现新颖先导化合物的重要资源,将为医药、食品、农业等领域开发新型活性物质的发现提供重要保障。植物内生放线菌资源丰富,从植物内生放线菌次生代谢产物中筛选有效的活性物质或新型化合物,将在很大程度上丰富人类医药宝库,解决自然资源不足等问题,尤其对某些珍贵或濒危植物内生放线菌活性物质的开发利用来代替宿主植物本身,能有效解决相关植物资源紧缺和培养条件苛刻等难题,实现可持续发展。随着人们对内生放线菌及其次级代谢产物活性物质研究的不断深入,相信会有更多、活性更强的化学成分被发现,内生放线菌次级代谢产物活性成分的研究无疑将具有广阔的应用前景。
现阶段对植物内生放线菌的研究程度远不如内生真菌与内生细菌。目前,植物内生放线菌的分离培养是其研究最大的瓶颈,由于宿主植物与其内生放线菌之间的协同生长关系复杂,研究人员很难在内生放线菌培养过程中模拟。而且人工培养过程中难免存在丢失内生放线菌所需要的生长因子,因此脱离宿主植物,很难在体外培养出内生放线菌,或者培养出的是退化或变异后的内生放线菌[25]。尽管有科研人员通过模拟微生物的生长环境设计出培养装置IChips[74],有望用于解决植物内生放线菌的培养困难问题,但是植物内生放线菌的研究仍存在诸多问题亟需解决。植物内生放线菌作为一种重要的微生物资源,对其开发利用仍然任重而道远。
猜你喜欢
湖北农机化(2020年4期)2020-07-24 09:07:16
世界农药(2019年4期)2019-12-30 06:25:10
国际呼吸杂志(2019年22期)2019-12-09 09:20:36
今日农业(2019年11期)2019-08-15 00:56:32
乡村地理(2018年2期)2018-09-19 06:44:02
天然产物研究与开发(2018年4期)2018-05-07 06:47:53
中成药(2018年1期)2018-02-02 07:20:03
应用海洋学学报(2014年4期)2014-11-22 07:43:54
天然产物研究与开发(2014年6期)2014-04-27 14:15:56
中国医学科学院学报(2013年2期)2013-03-11 20:25:49