嗜酸性硫杆菌群体感应系统研究进展
2021-09-24 00:28:42林建群陈林旭高雪彦林建强
微生物学杂志 2021年4期
林建群, 陈林旭, 高雪彦, 林建强
(山东大学 微生物技术国家重点实验室,山东 青岛 266237)
1 嗜酸性硫杆菌简介
嗜酸性硫杆菌属(Acidithiobacillus)是变形杆菌门中一类革兰阴性、化能自养的硫氧化细菌,以前被归为硫杆菌属(Thiobacillus),基于这类微生物极端的耐酸性和更近的同源进化关系,因此重新划分形成了新的嗜酸性硫杆菌属[1-2]。该属中细菌均具有高效的氧化各种还原性无机硫化合物和单质硫的能力,其中有些种群还具有氧化亚铁的能力[3]。嗜酸性硫杆菌通过硫、铁氧化获得电子以合成生物能(ATP)和还原力(NADPH),用于固定二氧化碳进行自养生长。嗜酸性硫杆菌(Acidithiobacillusspp.)广泛存在于陆地、海洋含无机硫的环境中,比如矿山、热泉、含硫的土壤及沉积物中[4]。目前嗜酸性硫杆菌属已发现报道有9个种,根据其能源底物的代谢差异,可分为既能氧化硫又能氧化亚铁的硫铁氧化种群,只能氧化硫的硫氧化种群(表1)[5-7],其中嗜酸性氧化亚铁硫杆菌(A.ferrooxidans)、嗜酸性氧化硫硫杆菌(A.thiooxidans)和嗜酸性喜温硫杆菌(A.caldus)是目前嗜酸性硫杆菌属中研究与应用最为广泛和深入的种群。
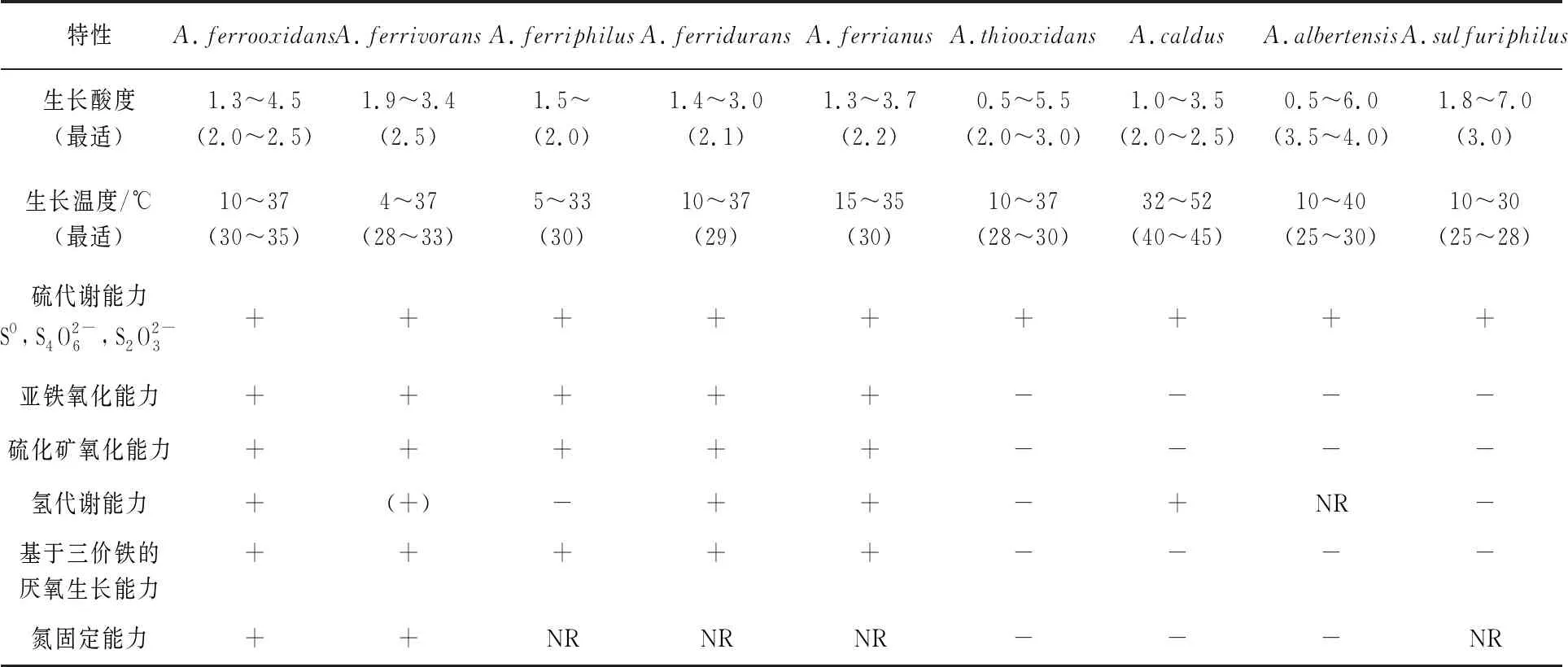
表1 嗜酸性硫杆菌属中不同种群的生理特征
嗜酸性硫杆菌作为重要的极端环境微生物和自养微生物[8-9],从20世纪20年代开始就引起了微生物学家的广泛关注[10],系统开展了微生物菌种分离[10-11]、硫铁能量代谢过程[12-14]、碳代谢能量合成过程[15-16]、重金属抗性[17]、嗜酸机制[18-19]等多方面的研究。对嗜酸性硫杆菌的研究揭示了生命在极端环境下的生存与适应策略,推动了该类微生物的跨学科研究和应用。嗜酸性硫杆菌还是地球矿山、热泉等生境中碳、氮、硫、铁等生命元素循环,以及汞、砷、铜等重金属元素转化和迁移的重要驱动者,因此该类微生物已经成为地球科学研究中重要的地质微生物,用于研究揭示矿山等生境中元素生物化学循环过程、微生物矿物相互作用等重要科学问题[20-23]。嗜酸性硫杆菌还可作为微生物合成生物学中的底盘微生物,用于生产生物材料等有价值的工业产品,比如2019年Xiao等[24]利用A.ferrooxidans作为细胞工厂生产三氧化二铁并成功制备了电池电极材料。因此,嗜酸性硫杆菌因其独特的代谢特征和极端环境适应性,已成为生命科学、地球科学、环境化学等多学科研究中的模式微生物,具有重要的研究价值和意义。
嗜酸性硫杆菌的研究与应用还与人类的绿色发展密切相关。矿山环境中嗜酸性硫杆菌等化能自养菌驱动的元素生物化学循环过程,一方面导致矿山水土环境酸化与重金属污染,造成严重的生态环境安全问题[20,25];另一方面利用该生物化学循环过程发展形成了新型的绿色矿产资源开采技术——微生物冶金技术[26-27]。微生物冶金技术已实现低品位金、铜、铀等矿石浸出,有效拓展了人类可开采的有色金属资源总量,世界铜、金产量的近四分之一使用生物冶金技术开采生产。2011年《科学美国人》将微生物冶金技术列为可以改变世界的十大想法之一。人们还将矿物微生物重金属浸出的基本原理扩展到环境治理领域:利用嗜酸性硫杆菌处理电子垃圾回收重金属,处理含重金属的污泥、矿渣等工业废弃物[28-31]。我国周立祥团队利用嗜酸性硫杆菌的生物浸出作用研发的污泥生物沥浸技术2010年起成功推向工程应用[32],已在我国多个污水处理厂和生产企业得到实际应用。因此,利用嗜酸性硫杆菌等微生物形成的生物冶金、污泥沥浸等微生物浸出技术,已成为一种具有广阔应用前景和发展潜力的新型绿色资源开发与环境治理技术,为人类解决有色矿产资源短缺和重金属污染难题提供有效的解决方案。
2 嗜酸性硫杆菌中群体感应系统的类型
群体感应(quorum sensing, QS)是细菌细胞种内及种间信息交流的一种方式,在细菌代谢与行为、微生物繁殖与种群密度、微生物与环境互作等方面具有重要调节作用[33-34]。具有QS系统的细菌能产生并释放一种叫做自体诱导物(autoinducer)的信号分子,它随细胞密度增加而同步增加,当信号分子积累到一定浓度时会改变细菌特定基因的表达,也就是说当细菌的数量达到一定的密度(quorum)时才能发生感应现象(sensing)[35]。在革兰阴性细菌中,LuxI家族蛋白负责催化形成信号分子酰基高丝氨酸内酯(acylated homoserine lactone, acyl-HSL或 AHL),转录调节蛋白(LuxR家族)可以识别并结合acyl-HSL,形成LuxR-acyl-HSL复合物,从而调节基因转录(图1)。Acyl-HSLs类型信号分子由高丝氨酸内酯(Homoserine lactone)和酰基脂肪酸链(Fatty acid acyl chain)组成,信号分子间的差异与脂肪酸碳链的长度和脂肪酸碳链C3位上连接的官能团(-H、-OH、=O)密切相关[36]。
2005年,Farah等在A.ferrooxidans中首次发现 LuxI/R类型的QS系统(AfeI/R),该系统存在于一个由afeI-orf3-afeR组成的操纵子上,其中afeI编码信号分子合成酶,属于LuxI家族蛋白,afeR编码转录调节蛋白,属于LuxR家族蛋白,orf3编码未知功能的蛋白[37-40]。生物信息学分析显示,AfeI/R类型QS系统在嗜酸性硫杆菌属的硫铁氧化菌群中的分布最为广泛和保守(图2)[17]。在嗜酸性硫杆菌属的硫铁氧化种群中,除了A.ferrianus以及没有全基因组信息的A.ferriphilus外,A.ferrooxidans、A.ferridurans和A.ferrivorans中均发现了AfeI/R系统;而在只能氧化硫的硫氧化种群中,只有A.thiooxidans存在AfeI/R系统。保守的afeR-orf3-afeI类型操纵子可以在A.ferrooxidans、A.ferridurans和A.thiooxidans中发现,而在A.ferrivorans中为独立存在的afeI和afeR基因。几乎所有的A.thiooxidans菌株都含有afeR-orf3-afeI类型操纵子,但在A.thiooxidansATCC19377的菌株中不含有该操纵子,只具有同源性更低的AfeI和缩短的AfeR。基于生物信息学的分析结果,表明AfeI/R系统在嗜酸性硫杆菌属中存在显著的种群和菌株的差异,且AfeI/R系统在硫铁氧化种群中更具有普遍性[17]。
2007年,在A.ferrooxidans中又报道了另一套可能合成acyl-HSLs分子的系统,包括glyQ、glyS、gph、act等4个基因,分别编码苷氨酸t-RNA合成酶的α亚基和β亚基、磷酸酶、酰基转移酶,在大肠埃希菌中异源表达act基因可以产生acyl-HSLs类型信号分子[41]。由于没有发现Act相应的调控蛋白,并且是否在A.ferrooxidans中也产生信号分子还未有报道,因此Act系统是否是QS系统目前还存在很大争议[41-42],需要在今后的研究中确定Act在A.ferrooxidans中是否具有信号分子合成能力及可能存在的信号感知和传递通路。
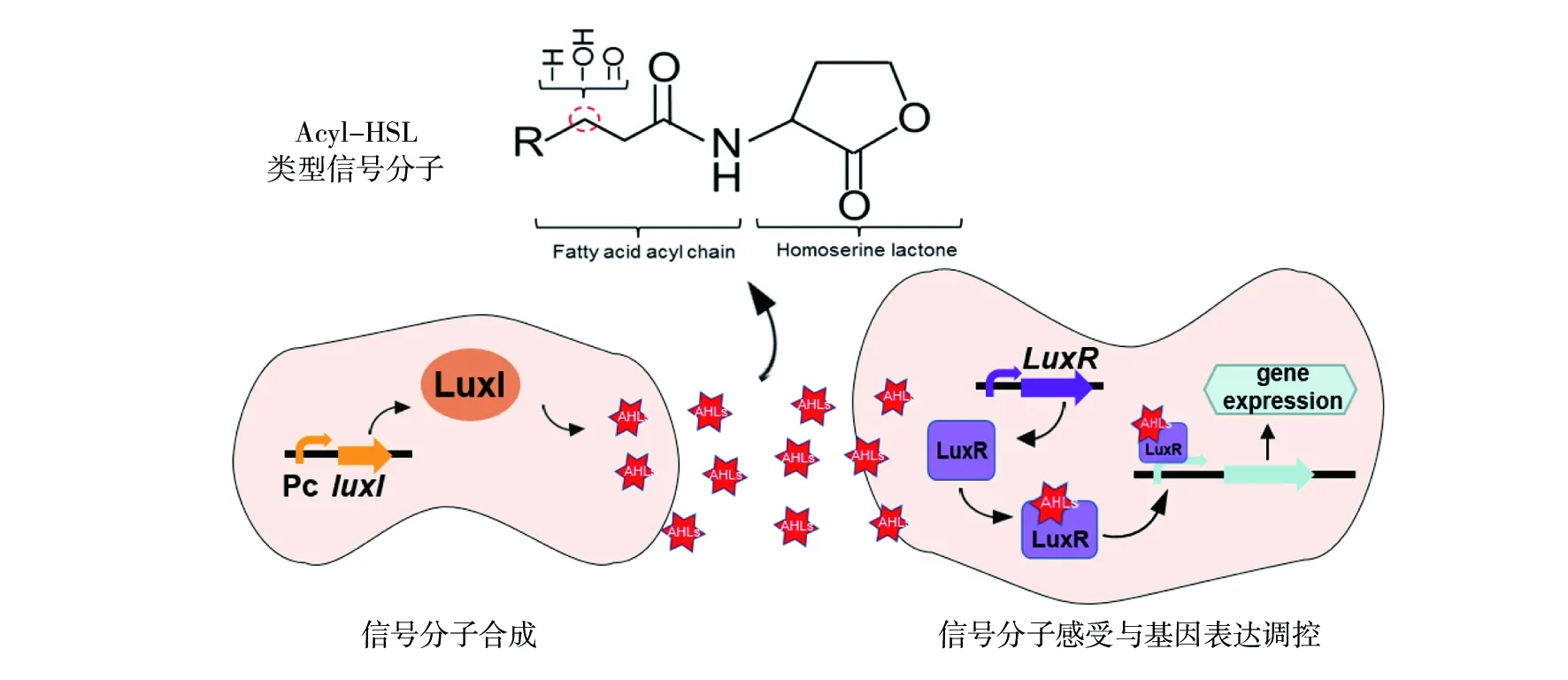
图1 LuxI/R系统调控方式与acyl-HSLs类型信号分子基本结构Fig.1 Regulation mode of LuxI/R and molecular structure of acyl-HSLs
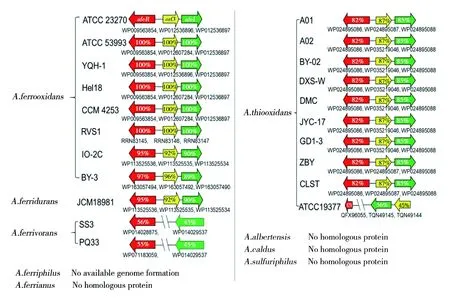
图2 AfeI/R类型群体感应系统在嗜酸性硫杆菌属中的分布情况Fig.2 Distribution of AfeI/R-like QS system in Acidithiobacillus
3 AfeI/R在A.ferrooxidans不同能源培养体系中的调控功能
AfeI/R系统在A.ferrooxidans中发现较早,但由于A.ferrooxidans等嗜酸自养菌的基因操作较为困难,直到2020年Gao等[17]通过对afeI基因的成功敲除和过表达才确定了AfeI/R在硫、铁培养体系中的调控作用。2005年,Rivas等[40]通过半定量的RT-PCR检测发现,afeI和afeR基因在以硫为能源底物培养时的表达量比以亚铁为能源底物培养时高,推测AfeI/R系统可能主要在硫培养,尤其是单质硫培养时发挥功能。2016年,Mamani等[43]通过添加acyl-HSLs类型信号分子的类似物,发现A.ferrooxidans对单质硫的吸附能力增强。2020年,Gao等[17]通过对afeI敲除和过表达菌株不同能源下的生长代谢分析研究,揭示了AfeI/R不仅在A.ferrooxidans单质硫培养时发挥重要作用,而且在亚铁培养时具有更加显著的调控作用,解答了多年来AfeI/R在A.ferrooxidans硫、铁培养体系中调控功能的疑问。
不同能源条件下,AfeI/R对A.ferrooxidans的调控效应存在显著差异。单质硫为唯一能源时,afeI过表达可以提高菌株在延迟期和指数期的细胞浓度和产酸能力,但不影响培养体系最终的群体密度,因此AfeI/R可以作为A.ferrooxidans单质硫培养时细胞生长的“加速器”,在延迟期和指数期调控细胞生长速度;亚铁为唯一能源时,afeI过表达会显著抑制菌株整个培养过程的细胞浓度和亚铁氧化能力,因此AfeI/R可以作为A.ferrooxidans亚铁培养时细胞生长的“抑制器”,调控细胞代谢生长和培养体系中最终群体密度[17]。AfeI/R在A.ferrooxidans培养体系中根据能源底物种类产生的不同调节效果,可能是A.ferrooxidans平衡自身硫、铁两套代谢系统适应硫、铁环境的一种重要方式,这也可能是嗜酸性硫杆菌属中硫铁氧化种群普遍具有AfeI/R系统的一个重要原因,因此相关研究成果为今后从环境能源底物与微生物代谢协同调控的角度,研究和认知AfeI/R的环境与生态效应提供了思路和启示。
4 AfeI/R在A.ferrooxidans不同能源底物下的调控方式
单质硫培养时,AfeI/R对A.ferrooxidans胞外多聚物(Extracellular polymeric substances, EPS)的调控,是实现AfeI/R调控A.ferrooxidans代谢生长和群体密度的关键[17,42]。单质硫是嗜酸性硫杆菌等化能自养硫氧化菌生长代谢中重要的固体能源底物,A.ferrooxidans等硫氧化菌分泌EPS及其在固体能源底物表面形成生物被膜(Biofilm)是微生物代谢利用单质硫的关键步骤[44-45]。群体感应系统是调控微生物EPS分泌和生物被膜合成的一种重要方式[46]。Gao等[17]的研究发现过表达afeI不仅可以促进A.ferrooxidans的EPS分泌与生物被膜形成,还可以增强细胞对单质硫的侵蚀作用、细胞硫氧化能力等特性,因此A.ferrooxidans利用AfeI/R对 EPS合成的调控作用,实现对其单质硫培养时细胞生长的调控。单质硫培养时,AfeI/R对A.ferrooxidans的EPS合成和细胞生长的调控具有显著的生长周期依赖性,即AfeI/R在延迟期和指数生长期可以通过调控EPS影响细胞生长,但在稳定期AfeI/R对EPS和细胞生长的调节作用消失。因此,AfeI/R基于A.ferrooxidans生长周期调控其EPS合成,可能是A.ferrooxidans适应环境单质硫固体能源的一种重要方式,其内在的分子机制及生态学意义有待进一步研究揭示。
当亚铁为唯一能源底物时,AfeI/R的调控作用如何实现呢?Gao等的实验研究发现afeI过表达不影响亚铁为唯一能源底物时A.ferrooxidans的EPS合成,说明亚铁培养时AfeI/R不是通过调控EPS合成影响细胞生长代谢和群体密度。通过转录组学的研究发现,亚铁培养时afeI过表达显著抑制了A.ferrooxidans中一个氢化酶合成基因簇的表达(AFE_0700-0719)。氢化酶是微生物氢代谢中的一个双向催化酶,推测在A.ferrooxidans中AfeI/R调控的氢化酶倾向于催化氢气生成质子和电子,因此相关研究发现和预测分析结果显示,A.ferrooxidans中AfeI/R可能通过调控微生物氢代谢,实现对亚铁培养时细胞生长代谢和菌群浓度的控制。综上,AfeI/R在A.ferrooxidans亚铁培养时可能具有新型调控方式,群体感应系统在化能自养菌铁代谢中具有基础性调控作用,这些发现揭开了微生物氢、铁代谢之间的紧密联系,在此基础上进一步开展相关代谢过程及相应调控分子机制研究,将有助于解答和认知极端环境下生命的生存及适应策略。
5 AfeI/R系统的信号分子种类及其功能
信号分子是群体感应系统执行功能的关键。2005年,Farah等[37]首次报道了A.ferrooxidansATCC 23270中存在acyl-HSL类型的信号分子,并且检测到该菌在亚铁、单质硫和硫代硫酸盐的三种培养条件下一共可以产生9种acyl-HSLs信号分子,分别是3-OH-C8-HSL、3-OH-C10-HSL、C12-HSL、3-OH-C12-HSL、3-O-C12-HSL、C14-HSL、3-OH-C14-HSL、3-O-C14-HSL、3-OH-C16-HSL。由于A.ferrooxidans存在AfeI/R和潜在的Act类型QS系统,一直无法确定AfeI/R系统真实产生的信号分子种类及其功能。2020年Gao等[17]通过敲除A.ferrooxidansATCC 23270中act基因和过表达afeI基因,构建了只含afeI的突变株,通过研究确定了AfeI在单质硫和亚铁培养时产生的信号分子种类(表2)。Acyl-HSLs类型信号分子合成时,其高丝氨酸内脂环衍生自 S-腺苷甲硫氨酸(S-adenosylmethionine,SAM),而其酰基侧链来自脂肪酸代谢,由酰基载体蛋白 (acyl-ACP) 提供,acyl-HSL分子结构的差异取决于酰基载体蛋白提供的酰基侧链的碳链长度以及第三个碳原子上的取代基[47-49]。A.ferrooxidans在硫、铁不同能源下代谢差异可能导致细胞产生不同种类的酰基侧链,进而影响AfeI催化合成的信号分子类型。因此,A.ferrooxidans等嗜酸性硫铁氧化菌在不同能源下信号分子的合成过程及形成差异,也将是今后AfeI/R类型群体感应系统研究的关键工作,通过对信号分子合成过程的解析,将有利于从分子水平阐明AfeI/R系统与能源底物的密切关系。
AfeI合成的多种信号分子中,一些关键的信号分子只有在特定能源底物条件下才能发挥调控作用。Gao等[17]通过信号分子标准品的反添加实验,证实3-OH-C14-HSL和C12-HSL是调控A.ferrooxidans生长代谢和群体密度的两个关键信号分子。3-OH-C14-HSL可以在A.ferrooxidans单质硫培养的前期,调控EPS合成和细胞生长代谢,但不影响培养体系最终细胞浓度;C12-HSL可以在A.ferrooxidans亚铁培养时,抑制其代谢生长和最终群体密度。由于C12-HSL只在A.ferrooxidans单质硫培养体系中检测到,而未在亚铁培养体系中检测到(表2),并且亚铁培养时含量最低的3-OH-C16-HSL由于缺乏标准品,目前还没有反添加实验验证,因此是否3-OH-C16-HSL或者还存在其它未鉴定的新型信号分子,在A.ferrooxidans亚铁培养时发挥调控功能,该问题的研究和解决将是今后A.ferrooxidans亚铁培养过程AfeI/R调控机制研究的关键问题之一。
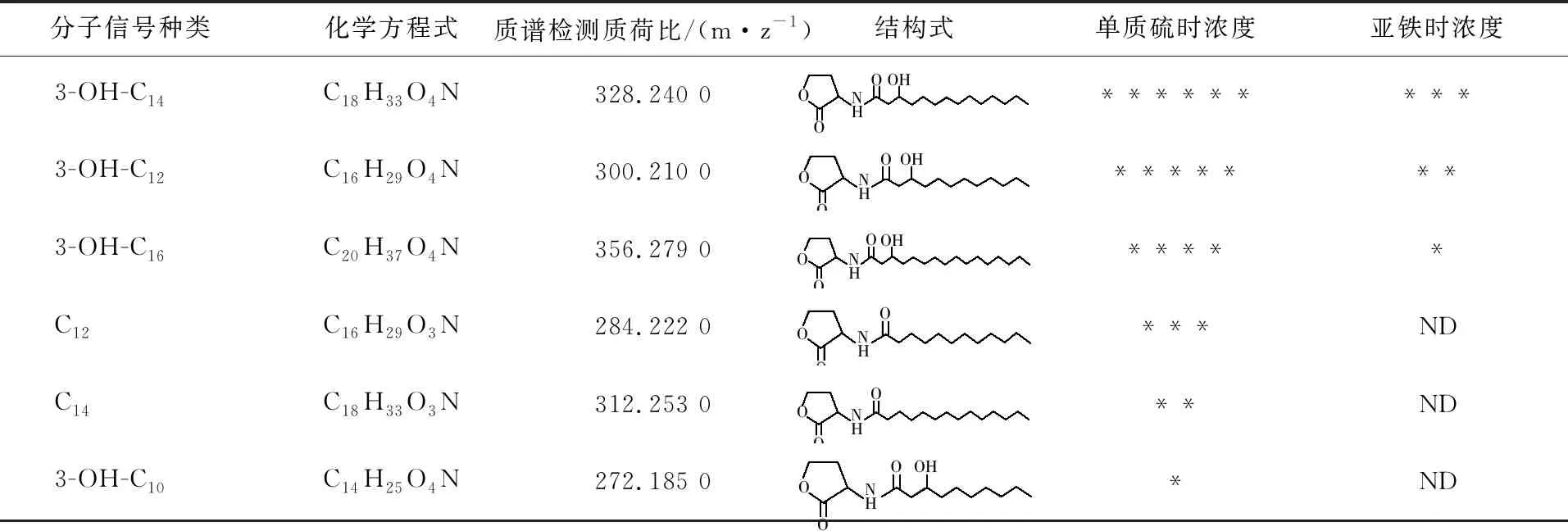
表2 A.ferrooxidans单质硫或亚铁培养液中信号分子的种类
6 AfeI/R在A.ferrooxidans中基于能源底物的调控策略
A.ferrooxidans培养体系中,能源底物可以影响AfeI合成的信号分子种类、信号分子的调节效应、AfeI/R的调控途径及其群体效应等[17]。基于这些研究发现,Gao等[17]提出AfeI/R系统在A.ferrooxidans中基于能源底物的调控策略理论,即AfeI根据能源底物种类产生不同类型的信号分子,特定功能的信号分子在特定能源底物条件下执行调控功能。如图3所示,单质硫培养时,AfeI合成的3-OH-C14-HSL通过调控EPS合成影响细胞硫代谢与生长,但该调节效应只发生在生长延迟期和指数期,不影响培养体系的最终群体密度。单质硫培养时产生的另一种信号分子C12-HSL可能具有调控A.ferrooxidans亚铁氧化系统的功能,因为AfeI过表达导致A.ferrooxidans亚铁氧化系统的一些基因表达下调,并且C12-HSL对亚铁培养的A.ferrooxidans具有抑制作用[17]。因此A.ferrooxidans在单质硫培养时,AfeI/R可能通过3-OH-C14-HSL调控其硫代谢系统,通过C12-HSL调控其亚铁代谢系统,实现对A.ferrooxidans硫、铁两种能量代谢系统的协同调控。亚铁培养体系中,AfeI/R可能通过调控细胞氢代谢和亚铁代谢过程,控制细胞生长和群体密度,但亚铁培养时发挥调控作用的关键信号分子仍需进一步解析确定。A.ferrooxidans中的AfeI/R基于能源底物调控策略理论的提出,表明群体感应系统可能是化能自养硫铁氧化菌响应和适应极端环境的重要机制,为研究和揭示A.ferrooxidans等硫铁氧化菌群与矿山酸性环境协同演化过程提供了新理论与新思路。
嗜酸性硫杆菌是矿山酸性微生态系统形成、演化的关键功能菌群,该类微生物的硫铁能量代谢过程、EPS分泌及生物被膜形成过程是矿山环境硫铁元素循环和矿物氧化过程中的重要驱动因素。群体感应系统形成的从细胞分子水平到群体行为的调控方式,是微生物群落与环境协同演化研究中的重要研究内容。A.ferrooxidans中AfeI/R基于能源底物调控策略的发现,表明一些化能自养硫铁氧化菌可能利用群体感应系统,构建起环境能源底物-群体感应信号分子-基因表达调控-细胞代谢与行为-菌群更替与演化-环境条件与物质组分变化的微生物群落与硫铁环境协同演化过程,这可能是硫铁氧化菌群适应矿山硫、铁能源环境,获得生态竞争优势的一个重要方式。因此A.ferrooxidans中AfeI/R调控策略的发现及相应理论的提出,将有助于今后从环境生态系统角度解析群体感应系统的调控效应与环境生态作用。
7 AfeI/R系统在微生物矿物氧化过程的调控作用
目前的硫化矿生物浸出理论认为嗜酸性硫杆菌等化能自养菌通过分泌EPS吸附到矿石表面并聚集形成生物被膜,细胞与矿物之间的EPS层提供了矿石生物、化学氧化的活性基团及反应发生的微环境,因此浸矿菌群EPS分泌及生物被膜形成是调控微生物矿物氧化过程的关键[45],在生物冶金产业应用和自然环境矿山酸性污染治理中具有重要的应用价值。
群体感应系统是微生物调控其EPS分泌和生物被膜形成的重要方式。2013年,González等[42]研究发现添加C12/C14-HSLs混合信号分子,可以促进A.ferrooxidans在矿石表面形成生物被膜。2020年,Gao等发现afeI/R基因簇过表达菌对矿物的吸附作用、侵蚀能力以及细胞重金属浸出能力显著增强,并提出了群体感应系统调控微生物矿物浸出的理论模型[50]。如图4所示,在矿物培养系统中A.ferrooxidans细胞分泌的信号分子通过积累达到一定阈值后,可以促进细胞分泌EPS,游离状态的细胞吸附能力增强,在矿石表面加速聚集形成生物被膜,进而增强了A.ferrooxidans的硫代谢能力、矿物侵蚀和氧化能力,加速矿物的重金属浸出过程[50]。AfeI/R调控微生物矿物浸出过程的发现和调控理论的提出,表明群体感应系统在调控嗜酸性硫杆菌等浸矿微生物矿物氧化过程中具有重要作用,为研究和开发微生物矿物浸出过程调控手段和技术提供了理论基础和方法指导。
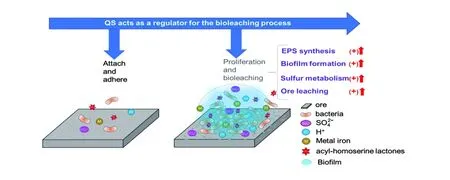
图4 AfeI/R调控A.ferrooxidans矿物浸出过程的理论模型Fig.4 The regulation of AfeI/R on the bioleaching process of A.ferrooxidans
8 展 望
嗜酸性硫杆菌等自养微生物中群体感应系统的研究不仅有助于认知群体感应系统在极端微生物中的分子调控功能与调节机制,也有利于揭示极端环境下微生物的生存与适应策略,实现新认知,构建新理论,开发新技术。目前的研究表明,群体感应系统参与微生物硫铁代谢过程、微生物矿物相互作用以及嗜酸菌与极端环境协同演化过程,揭示了群体感应系统在化能自养微生物中独特的功能及其基础性调控作用。基于前期研究成果和理论模型,今后可以从群体感应系统的胞内调节分子机制、种间调控作用、环境生态效应以及应用技术开发等几个方面开展系统研究:①嗜酸性硫杆菌群体感应系统调控的详细分子机制:以A.ferrooxidans为例,系统解析各种信号分子可能的调控作用,重点研究确定亚铁培养时起调控作用的关键信号分子;揭示信号分子-调控蛋白-调控序列-调节基因与代谢途径的信号感受与传递通路;通过解析信号分子结构、调控蛋白结构、调控序列特征,在分子层面揭示群体感应系统在不同能源体系下信号传递和调控通路,建立完善、精确与系统的AfeI/R调控分子网络,形成化能自养菌群体感应系统研究的理论创新与突破。②嗜酸性硫杆菌等化能自养微生物中群体感应系统发掘及其种间调控作用研究:以矿山环境中硫铁氧化菌为对象,发掘不同种群的群体感应系统及其同源关系;解析不同种群中群体感应系统的调控功能及调控网络;研究不同种群间群体感应系统的信息传递和调节效应;通过对不同种群群体感应系统的解析,揭示矿山环境不同硫铁氧化种群基于群体感应系统的调控策略,建立基于群体感应系统的不同硫铁氧化种群间的协同调控网络。③群体感应系统在嗜酸菌群与酸性环境协同演化中的作用:A.ferrooxidans中群体感应系统存在典型的依赖环境能源底物的调控方式,并且环境能源底物能够影响A.ferrooxidans群体感应系统信号分子合成及其调节效应,因此群体感应系统可能是嗜酸性硫杆菌等化能自养嗜酸菌群适应矿山硫铁环境而形成的独特环境适应调控机制,通过群体感应系统的调控作用,嗜酸自养菌群可能更利于在矿山环境微生物竞争中占据生态优势,成为优势功能菌群,驱动环境酸化和嗜酸微生物群落形成。因此利用环境微生物组学研究理念和技术并结合实验模拟,系统揭示群体感应系统的生态与环境调节效应,将是未来从环境生态学角度研究群体感应系统的重要内容,将有助于解答微生物与环境协同演化等重要的科学问题。④基于嗜酸自养微生物群体感应系统的功能开发新型应用技术:嗜酸性硫杆菌等化能自养菌群在生物冶金等生产实践和矿山酸性环境治理中都是关键的功能菌群。在微生物冶金等产业应用中,群体感应系统可以作为浸矿菌合成生物学研究和改造的重要遗传元件,选育高效的浸矿菌株和复合浸矿菌群,强化浸矿菌群的矿物氧化与重金属浸出过程;另外还可以针对浸矿菌群群体感应系统的调控特点优化浸出体系组成,针对性开发矿物浸出过程调控技术,提高生物冶金效率。在酸性矿山环境治理方面需要抑制嗜酸菌群体感应系统生态环境调控作用的发挥,针对嗜酸菌群体感应系统可以开发阻遏物或者寻找抑制微生物,打断或削弱群体感应系统的信号传递以调控过程,阻止矿山环境嗜酸菌的形成种群优势,实现从源头上控制矿石生物氧化与环境酸化过程,构建矿山酸性环境生态修复的新理论和新技术。
猜你喜欢
能源工程(2022年1期)2022-03-29 01:06:40
云南化工(2021年10期)2021-12-21 07:33:28
云南化工(2021年6期)2021-12-21 07:30:56
科学(2020年2期)2020-08-24 07:57:00
福建基础教育研究(2019年8期)2019-05-28 08:39:51
中学生数理化·八年级物理人教版(2017年6期)2017-11-09 06:00:43
中国中西医结合皮肤性病学杂志(2016年4期)2016-07-18 10:59:52
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15 12:47:46
生物技术通报(2015年1期)2015-04-10 16:15:19
卫生职业教育(2014年9期)2014-02-16 07:22:54