
植物单宁分离提纯及功能活性研究进展
2021-09-21 08:16黄达荣张雅甄郑百涛
食品与机械 2021年8期
黄达荣 张雅甄 郑百涛 杜 冰 黎 攀
(华南农业大学食品学院,广东 广州 510642)
植物单宁是一种天然多酚类化合物,多存在于陆地与海洋的大部分植物之中。其可以与唾液蛋白质结合,也可以联合动物胶原多肽链从而增强皮的韧性,前者是涩味的来源,后者是鞣革的原理,因此单宁也被称作“鞣质”。单宁作为植物的次生代谢产物,是丰富度仅次于纤维素、半纤维素和木质素的生物质资源[1],主要分布在植物的根[2]、茎[3]、叶[4]及皮[2-3]中,具有抗炎、抗氧化、抗肿瘤等活性[5-7],被广泛应用于食品、医药及制革[8]等领域。目前,常用的单宁提取方法主要为浸提法与萃取法,并辅以超声波、微波、加压等[9],而提取液多为甲醇、乙醇与丙酮等有机溶剂[10],但不同植物来源的单宁需要优化其提取与纯化工艺。
文章拟综述单宁的分类与应用,以及近年来植物单宁的提取与纯化方法的发展现状,并提供一个通用于大部分植物单宁提取与纯化的方法,同时归纳单宁功能活性及其机制,以期为单宁的高效获取与高值化应用提供依据。
1 植物单宁的分类及应用
1.1 植物单宁的分类
植物单宁可根据化学结构、来源品种和化学特性进行分类,目前普遍按照单宁的结构差异将其分为水解单宁和缩合单宁两大类(见图1),其相对分子量约为500~20 000 Da。此外,还有复合单宁和褐藻单宁,但两者罕见于陆生植物中,复合单宁由黄烷-3-醇与水解单宁结合而成,褐藻单宁是间苯三酚的聚合产物,可占海藻干重的30%,其已被证明具有抗菌、抗氧化等活性[11]。植物单宁的常见类型可由表1的方法[12]进行鉴别。

图1 水解单宁与缩合单宁的典型结构

表1 水解单宁与缩合单宁的鉴别反应
水解单宁即可水解的单宁,是一种以多元醇为中心,如葡萄糖,通过酯键连接数个酚酸及其衍生物的化合物[13],其中多元酚酸是决定单宁性质的关键部分。水解单宁遇酸、碱或酶时容易水解生成小分子量的化合物,根据水解生成的多元酚羧酸不同,将水解单宁分为鞣花酸单宁和没食子酸单宁。此外,没食子酸衍生物也是水解单宁的一类,其一般含有6个及以上的没食子酰基单元。
缩合单宁又称原花色素,以不含糖核的类黄酮、黄烷-3-醇(见图2)作为结构单元聚合而成,其A环可出现间苯二酚或间苯三酚形式,而B环可出现邻苯二酚或邻苯三酚形式[14],构成了不同的缩合单宁结构单元[15];分子间以碳碳键相连且不含糖基,不易被水解;与水解单宁相比,缩合单宁的分子量范围较广,可以在酸、碱或酶作用下发生缩合反应,而进一步的缩聚会产生不溶于水的红粉[12],但高相对分子质量的缩合单宁的分子体积大,在空间中移动时受到的阻力大,导致其生物活性较低。
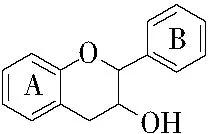
图2 黄烷-3-醇的结构
1.2 植物单宁的应用
单宁不仅可以作为调味剂在茶、咖啡等饮料中调节风味,用于生产速溶饮料,还能影响食品色泽,其机理是单宁在环境中容易被氧化为醌类物质而加深颜色。此外利用单宁与蛋白结合的性质,将牛血清蛋白与单宁酸制成多层微胶囊,可延缓胃酸的侵蚀,有效保护胶囊中的乳铁蛋白,并制备多层微胶囊的乳铁蛋白口服制剂[16],而固定化单宁可在食品加工领域中纯化活性肽与蛋白质、澄清酒与饮料[17];单宁还可以抑制食品腐败菌而延长食品保藏期[18]。此外,单宁在制革[19]、医药[20]与水处理[21]等领域也发挥着重要作用。
2 植物单宁提取与纯化
2.1 提取方法
单宁的含量测定[22]见表2。目前,单宁的定量分析方法有高效液相色谱法、液质联用法等,这些方法较多应用于总酚含量的测定,尽管有测定缩合单宁的方法,但几乎没有一种可以普适于各种植物单宁定量的方法,因为定量时常用的如没食子酸、儿茶素、表儿茶素等标准品,与各种植物单宁组分差异较大,其标准品选择仍是一个尚未解决的问题。因此目前定量植物单宁一般参考前人文献的方法,或者对多种测定方法进行内标法等方法学筛查,确定含量后再经高效液相色谱分离可以明确植物单宁组分。

表2 单宁的定量方法
2.1.1 溶剂浸提法 溶剂提取法是提取单宁的基础方法,可以分为无机溶剂法和有机溶剂法,如水和甲醇、乙醇、丙酮等。其中热水提取是中国最传统的方法,虽然单宁溶于水、乙醇与丙酮,但植物中也含有水溶性的树脂、蛋白质、淀粉等杂质,直接用水提取所得到的植物单宁含量较低,水与有机溶剂混合后进行提取成为主流,如水—丙酮体系能破坏单宁与蛋白质的连接键,提高单宁提取率[23]。为了更高效地提取单宁,在确定提取溶剂的基础上,辅以超声波、微波、加热等方法(见表3),并利用正交试验与响应面试验进行单宁提取工艺优化。
2.1.2 超声波、微波提取和酶解转化法 单宁是植物细胞产物,细胞膜和细胞壁是单宁提取的主要阻碍。超声波具有机械效应、空化效应及热效应,微波则使胞内水分吸收微波能而汽化,对细胞膜和细胞壁产生压力而形成小孔,酶能选择性地分解细胞膜与细胞壁,且条件更为温和,因此酶辅助提取已被广泛用于从食品和植物中提取不同种类的化合物[34]。三者一般用于辅助溶剂对植物单宁进行提取。

表3 不同植物单宁的提取方法†
2.1.3 超临界流体萃取法 超临界流体具有液体和气体的双重性质,通过控制压力和温度即能选择性地萃取出植物单宁,二氧化碳因其清洁、成本低、萃取时性质稳定等特点,目前被用作超临界流体,且利用超临界二氧化碳萃取可以保护单宁免受氧化。姜萍等[35]应用超临界二氧化碳提取五倍子中的单宁酸,经条件优化后得率高达57.83%,但目前国内外较少应用该技术提取单宁,可能是萃取所用仪器要求高,成本也较高。
2.2 纯化方法
2.2.1 溶剂萃取及沉淀吸附法 单宁在醇类、丙酮等有机溶剂中溶解度较高,而高浓度乙醇能沉淀植物的大部分蛋白质,因此有机溶剂均能在提取与纯化单宁时发挥作用,但利用溶剂萃取仍处于初步纯化阶段且溶剂残留会影响产品质量。此外,还可以利用疏松多孔的活性炭或皮粉吸附大量杂质,其中包括植物色素、树胶等。
2.2.2 色谱分离法 目前制备高纯度单宁及有关化合物的主要方法是色谱分离法。固定相一般选用大孔树脂和葡聚糖凝胶,前者的耐压性较强,装柱后能承载较高流速而不塌陷,纯化效率比后者高,而后者以Sephadex LH-20为主。范小曼[36]联合运用XAD16HP大孔树脂与葡聚糖凝胶LH-20提纯了白花败酱单宁,并对单宁组分进行了有效分离。在对不同植物单宁进行纯化时,需先筛选合适的固定相,后进行静态与动态的解吸、吸附试验,以确定层析柱的上样浓度、上样流速、洗脱剂类型及浓度等参数。
2.2.3 膜分离 膜技术以选择透过性的膜为基础,如微滤、纳滤和超滤膜,为新兴的绿色技术,但近年来鲜有报道,可能是因为大部分植物的单宁含量较低,提取时需要除去大量杂质,易导致膜堵塞。杜运平等[37]采用聚砜中空纤维膜分离了板栗苞单宁,纯度约为64.00%。
3 植物单宁的功能活性及机制
3.1 抗氧化
单宁的抗氧化性体现在4个方面:① 作为氢供体通过还原反应降低环境中的活性氧、自由基等;② 螯合金属离子,避免催化过氧化物进一步氧化;③ 调控机体中的氧化还原系统[38],还原型谷胱甘肽可以上调Bcl-2与Bax的比值,清除机体内自由基,而丛明等[39]研究发现,用原花青素灌胃大鼠,能显著增加大鼠大脑皮质的Bcl-2/Bax值,调控集体氧化系统;④ 调控机体与氧化应激的相关信号通路,缓解应激状态下的损伤,目前主要调控的信号通路为Keap1-Nrf2/ARE,而杏仁皮中的缩合单宁能激活该通路[40],提高机体抗氧化蛋白的表达量。单宁的抗氧化活性是触发抗炎等有益机制的基础,其可能涉及的机制如图3所示,但具体的作用机制仍需更深入的研究,特别是考虑到不同植物的单宁的给药剂量及其代谢图谱的多样性。此外,单宁还能与肠道相互作用,黄烷-3-醇单体能被肠道吸收[41],但其聚合体能被肠道菌群降解吸收或与肠道蛋白结合而增强肠道屏障。
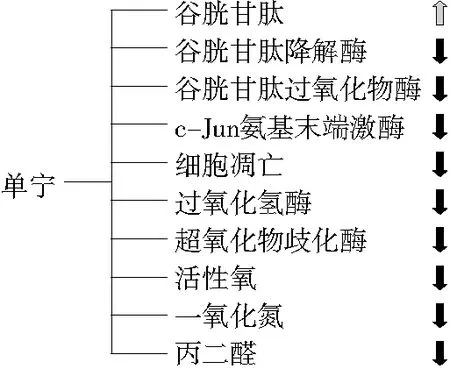
图3 单宁的抗氧化特性可能涉及的机制[11]
3.2 抑菌
单宁对致龋细菌、沙门氏菌、金黄色葡萄球菌等致病菌[42]具有显著的抑制效果,还可以促进牙体组织矿化、治疗牙周及牙体牙髓疾病,因此被广泛应用于口腔医学领域。单宁对细菌抑制的重要结构是酚羟基,而单宁抑菌的机制主要分为:① 螯合细菌所需的金属离子,如铁和铜;② 抑制并破坏细胞壁和细胞内外膜的形成,减少病菌的粘附;③ 与细菌特定靶标蛋白和药物外排蛋白结合,提高药物对细菌的杀伤力;④ 抑制细菌代谢所需的酶[43],影响细菌的蛋白质合成、能量代谢和氧化还原系统;⑤ 增强机体免疫力,如草莓单宁可以抑制NF-κB信号通路,降低人体炎症因子的水平[44]。
3.3 抗病毒
植物单宁对冠状病毒[45]和诺如病毒[46]有显著抑制能力,后者利用NoV P蛋白开发了一种以唾液为基础的结合—阻断试验,并以此来评价50种植物单宁抑制诺如病毒的能力,从而筛选出石榴单宁与五倍子单宁作为抗诺如病毒的潜在候选药物,这提示需要建立植物单宁的标准化药效评价体系。单宁抗病毒的机制与抑菌类似,会抑制病毒的酶与生物膜,阻止病毒侵入细胞和细胞核,抑制病毒反转录和复制,此外单宁还会削弱病毒粘附细胞的能力、运动能力[47],同时单宁促使病毒蛋白质变性、阻断病毒的胞间传播来实现抗病毒的功能,但不同来源和化学结构的植物单宁抗病毒性能也有所差异,因此需进一步探索其具体抗病毒机制和动物试验。
3.4 抗肿瘤
单宁可以通过抑制端粒酶活性、激活内质网应激介导的UPR信号通路诱导肿瘤细胞凋亡。其中肿瘤细胞内ROS含量是调控细胞生长的关键,一方面ROS能加快肿瘤细胞繁殖分化、激活转移相关的信号通路,而另一方面高浓度的ROS含量能诱导肿瘤细胞凋亡;单宁酸能提高肾癌细胞中ROS含量、降低黏附分子ICAM-1的表达量,促进肾癌细胞的凋亡并抑制细胞集聚与转移。同时抗炎、抗氧化的机制也对抑制肿瘤细胞具有积极影响。已有研究[48]证明植物单宁对结肠直肠癌、胰腺癌细胞的抑制作用,但多数研究仅聚焦于细胞模型的评价,这与动物模型具有较大差异,细胞模型的积极作用不能等同于动物试验的药效,因此需要建立小鼠或大鼠的肿瘤模型进行药效试验。
3.5 调节血糖
单宁降低血糖的作用已在脂肪细胞、大小鼠、幼猪、鱼类为对象的试验中得到了证实[49-50]。其血糖调节机制可分为:① 抑制α-葡萄糖苷酶等食物分解酶的活性,影响机体吸收;② 促进胰岛β细胞再生和胰岛素的分泌;③ 促进糖酵解,抑制糖异生;④ 作用于肠道菌群而调节血糖[50]。单宁对2型糖尿病的症状有改善作用[51],但血糖调节的分子机制仍是单宁降血糖的研究点,单宁对糖酵解、糖异生途径的影响研究不足,尤其是胰岛素与糖代谢的信号通路具有较大的研究潜力。
3.6 抗炎
Aguiar等[52]报道原花青素能通过抑制新陈代谢和炎症递质而起到积极的抗炎作用;Pallarès等[53]发现葡萄籽的原花青素可以降低营养型肥胖大鼠模型的慢性低度炎症。综上,单宁的抗炎能力可能是调节细胞炎症因子水平如下调肝脏组织中的TNF-α与IL-6水平、抑制NO与前列腺素E-2等致炎物质的产生、防止致炎物质与其他分子络合,更具体的抗炎机制还有待探索,但单宁的抗炎活性已被证明,且单宁的抗炎能力与其抗氧化和抗菌等能力相关,因为致病菌和超氧化物与炎症具有密切联系。
3.7 抑制寄生虫
植物及其提取物已被用作人类和家畜的驱虫剂。从水解单宁的角度来看,其干扰虫卵孵化和幼虫运动的3种机制[54]为:① 拥有强蛋白沉淀能力的水解单宁与卵和幼虫通过非共价键结合;② 具有高氧化活性的水解单宁可能被自身氧化,并且氧化产物与卵和幼虫共价结合;③ 水解单宁进行水解,并且水解产物可能通过非共价或共价相互作用与卵和幼虫相互作用,从而破坏卵的发育及幼虫运动,但缩合单宁的驱虫机理与调控机制尚未明确,有待进一步研究。
4 小结与展望
目前单宁主要是从虫瘿五倍子中提取,但随着单宁需求量的上升,不同植物的单宁开发逐渐受到关注,而植物单宁的自然资源丰富,具有广阔的应用前景。单宁类化合物结构复杂,有关单一植物单宁的结构和营养特性研究还不够充分,学科间的交叉研究较为欠缺,导致部分结论与作用机理不明确,在不同植物单宁的生物活性研究方面难以形成完整系统。
在单宁功能活性产品开发方面,其难点在于工作人员需扩大研究范围,针对不同来源的植物单宁中起功效的活性成分、结构特点和发挥作用的机理进行探索,才能实现植物单宁在产品开发上的精准应用。但植物单宁的不同提取来源和添加浓度,以及不同动物种类和生理状态会影响其添加效果。后续需对不同植物单宁的构效关系以及单宁的添加剂量进行标准化,建立不同药效评价模型,并分析不同植物单宁的组分与结构,以明晰不同分子结构类型以及不同剂量单宁的作用机理,实现单宁在各个领域中的应用。
猜你喜欢
河北科技师范学院学报(2022年2期)2022-08-26
中外葡萄与葡萄酒(2022年4期)2022-07-26
酿酒科技(2022年5期)2022-05-24
世界科学技术-中医药现代化(2021年8期)2021-12-21
载人航天(2021年5期)2021-11-20
粉末冶金技术(2021年1期)2021-03-29
中成药(2019年12期)2020-01-04
中国果菜(2019年11期)2019-12-14
中学化学(2019年3期)2019-07-08
中学化学(2016年2期)2016-05-31
