
布鲁氏菌BAB-RS25305蛋白对细胞炎症因子表达的影响
2021-09-15 05:37朱德馨陶婷婷王梓行邱润辉
动物医学进展 2021年9期
朱德馨, 吴 洁, 郭 嘉, 陶婷婷, 李 佳, 王梓行, 邱润辉,张 辉*
(1.石河子大学动物科技学院,新疆石河子 832000;2.新疆生产建设兵团第八师畜牧兽医工作站,新疆石河子 832000)
布鲁氏菌病(Brucellosis)是由布鲁氏菌属的细菌侵入机体引起传染-变态反应性的人兽共患传染病[1]。动物布鲁氏菌病的主要特征为关节炎、母畜流产、公畜睾丸炎等[2];布鲁氏菌在人类宿主中表现为潜伏期长,导致慢性、虚弱的感染,并伴有严重的临床表现,如发热、致炎、肝肿大和脾肿大等[3]。
布鲁氏菌主要感染吞噬细胞和树突状细胞[4]。当布鲁氏菌侵入宿主机体后,首先是持续7 d~15 d的急性感染期,导致宿主发生菌血症[5],宿主细胞的模式识别受体NLRP3炎性小体感知布鲁氏菌病原体相关的分子模式(Pathogen-associated molecular patterns,PAMP),如布鲁氏菌脂多糖(lipopolysaccharide,LPS)和鞭毛蛋白等[6],并激活caspase-1,导致白细胞介素-1β(IL-1β)和白细胞介素-18(IL-18)等促炎细胞因子的释放,进而引起炎症反应,而炎症反应的发生有利于宿主抵御布鲁氏菌的入侵[7-10]。另外有研究发现,炎症发生也与内质网应激(endoplasmic reticulum stress,ERS)有关,1995年Pahl H L等[11]发现,内质网中蛋白质积累引起的内部压力也诱导NF-κB DNA结合和NF-κB依赖的核基因表达。Maarten F等研究发现布鲁氏菌Ⅳ型分泌系统(type Ⅳ secretion system,T4SS)分泌蛋白VceC可以触发内质网应激从而导致布鲁氏菌炎症反应[12]。
本研究使用NCBI以及牛布鲁氏菌2308基因组的推导预测数据库prodoric release对BAB-RS25305基因进行生物信息学预测分析,发现BAB-RS25305基因属于通用应激蛋白家族,具有USP结构域。当细胞处于应激环境下其表达增强,并且能够增强细胞的存活率,但其在布鲁氏菌中具体功能研究甚少。本研究通过对BAB-RS25305基因进行原核表达并纯化,用其刺激小鼠巨噬细胞RAW264.7,检测其对炎症因子的影响,为初步探究BAB-RS25305蛋白的表达以及功能奠定基础,也为揭示布鲁氏菌的胞内生存以及致病机理提供理论依据。
1 材料与方法
1.1 材料
1.1.1 质粒E.coliDH5α、BL21(DE3)感受态细胞,购自北京全式金生物技术有限公司;T-Vector PMD19(simple),购自TaKaRa公司;pET-32a(+)载体,由石河子大学动物科技学院保存。
1.1.2 试剂 2×Taqmaster mix(Dye)、细胞RNA提取试剂盒、cDNA反转录试剂盒、His标签蛋白纯化试剂盒,康为世纪生物科技有限公司产品;琼脂糖凝胶DNA回收试剂盒、质粒小提试剂盒,北京天根生化科技有限公司产品;Hind Ⅲ、EcoR Ⅰ,TaKaRa公司产品;EtEraserSE内毒素去除试剂盒,厦门鲎试剂生物科技股份有限公司产品。
1.1.3 主要仪器设备 恒温摇床(SY-2230),深圳市塞亚泰科仪器设备有限公司产品;电泳仪(CIea-ver),诺王(上海)生物科技有限公司产品;PCR仪(Mastercycler nexus gradient),德国EPPENDORF公司产品;超灵敏化学发光成像系统(Fluor Chem E),普诺森生物Protein Simple公司产品;多功能酶标仪(Infinite 200M/F Pro),瑞士帝肯(TECAN)公司产品;高速冷冻离心机(Thermo Heraeus Biofuge Stratos),赛默飞世尔公司产品。
1.2 方法
1.2.1 引物设计与合成 根据NCBI数据库中GenBank所提供的BAB-RS25305基因序列,使用Primer 5.0生物软件设计引物(引物序列见表1),引物由北京睿博兴科生物技术有限公司进行合成。
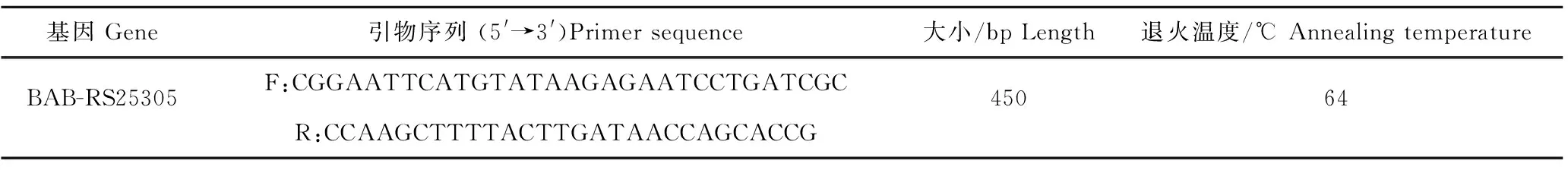
表1 BAB-RS25305基因引物序列
1.2.2 BAB-RS25305基因PCR扩增 以灭活的布鲁氏菌S2308为模板,进行PCR扩增。PCR扩增体系为20 μL:ddH2O 7.6 μL,上、下游引物各0.2 μL,模板2.0 μL,2×Taqmastermix(Dye)10 μL。反应条件:95 ℃预变性5 min;94 ℃ 40 s,64 ℃ 30 s,72 ℃ 40 s,共30个循环;72 ℃ 10 min;4 ℃终止反应。PCR产物用琼脂糖凝胶电泳检测,并使用琼脂糖凝胶DNA回收试剂盒对目的条带进行回收。将回收产物与T-Vector PMD19载体16 ℃水浴过夜连接,连接产物转化至E.coliDH5α感受态细胞后涂布于氨苄抗性的平板上,37 ℃倒置培养过夜。筛选阳性克隆菌,摇菌,菌液进行PCR鉴定并送北京睿博兴科生物技术有限公司进行测序。
1.2.3 BAB-RS25305重组质粒的构建与鉴定 将测序成功的PMD19-T-BAB-RS25305重组质粒和pET-32a(+)载体进行双酶切,酶切体系为20 μL:质粒10 μL,ddH2O6 μL,1×M buffer 2 μL,Hind Ⅲ 1 μL、EcoRⅠ 1 μL。酶切条件:37 ℃,酶切4 h。酶切产物回收后与pET-32a(+)载体16 ℃水浴锅过夜连接,然后转化至DH5α感受态细胞,筛选阳性克隆菌进行双酶切鉴定和送往公司测序。将测序和双酶切验证正确的重组质粒转化至E.coliBL21感受态细胞,涂布氨苄平板后筛选阳性菌进行菌液PCR鉴定和送往公司测序。
1.2.4 BAB-RS25305重组蛋白的诱导表达 将PCR鉴定和测序正确的克隆菌接种于20 mL含20 μL氨苄抗性的 LB培养基中,37 ℃、180 r/min过夜培养。取过夜培养菌液1 mL重新接种于氨苄抗性LB中,摇菌直至OD600 nm约为0.4左右。加入IPTG诱导蛋白表达,分别在0、2、4、6、8 h时间点各收取菌液1 mL,12 000 r/min离心1 min,弃上清,于沉淀中加入80 μL ddH2O后,再加入20 μL 5×SDS-PAGE Loding buffer,吹打混匀,煮样10 min后进行SDS-PAGE。观察电泳结果,确定BAB-RS25305重组蛋白最佳表达时间,并在蛋白表达量最佳的时间点诱导表达目的蛋白。
1.2.5 BAB-RS25305重组蛋白的纯化 将活化菌液转接至120 mL LB液体培养基中,待其OD 600 nm达到0.6左右,以1 mmol/L的浓度加入IPTG诱导剂诱导蛋白表达,4 ℃离心机收集全部菌液,弃上清留下沉淀,然后用PBS缓冲液重悬菌体,并于液氮和42 ℃水浴锅反复冻融3次后超声破碎,离心后进行蛋白可溶性分析,用His标签蛋白纯化试剂盒对蛋白包涵体进行纯化,纯化后的蛋白依次通过6、4、2、1 mol/L尿素和PBS缓冲液进行透析复性,复性后的蛋白使用蔗糖进行浓缩,BCA试剂盒检测蛋白浓度,最后用EtEraserSE 内毒素去除试剂盒去除蛋白中内毒素,蛋白浓度低于1 EU/mL可用于后续试验。
1.2.6 RAW264.7小鼠巨噬细胞的培养 从液氮罐中取出冻存的RAW264.7细胞,放入37 ℃水浴锅中,不停搅拌至完全融化,细胞悬液移至15 mL离心管中,1 000 r/min、离心5 min。用细胞培养液将细胞吹打混匀后移入中号培养板中,培养24 h;吸去上清,PBS冲洗3遍,加入胰酶消化,弃胰酶加入3 mL左右细胞培养液吹打细胞至悬浮,移至大号培养板中在体积分数为5%的CO2、37 ℃条件下培养24 h。
1.2.7 BAB-RS25305重组蛋白刺激小鼠巨噬细胞RAW264.7 将培养3 d后的细胞,进行细胞计数,调整细胞密度为1×106个/mL,铺6孔板。试验组分为:BAB-RS25305重组蛋白刺激组以及PBS阴性对照组。BAB-RS25305重组蛋白组:6孔板中加入终浓度为50 μL/mL的纯化的BAB-RS25305蛋白;阴性对照组:加入等体积的PBS作为对照。铺好板后,在体积分数为5%的CO2、37 ℃条件下培养箱中继续培养24 h。
1.2.8 检测巨噬细胞中NLRP3和caspase-1 RNA水平的表达 6孔板孵育24 h后,弃上清,用500 μL Trizol Reagent裂解细胞,收集于 RNase-Free 1.5 mL离心管中,按照RNA提取试剂盒提取RNA,Nanodrop-2000检测RNA浓度。然后按照cDNA反转录试剂盒将RNA反转录成cDNA,进行荧光定量PCR。反应体系为10 μL:SYBR Green Real-time PCR Master Mix 5 μL、ddH2O 3.6 μL、上、下游引物各0.2 μL、cDNA模板1 μL。每个样品重复3次。以NAPDH为内参,用2-△△Ct法来对NLRP3和caspase-1的相对表达量进行分析。
1.2.9 检测巨噬细胞中炎症因子蛋白水平的表达 6孔板孵育24 h后,弃上清,PBS冲洗3遍,细胞刮刮下细胞。1 000 r/min离心5 min,弃上清,加入250 μL RIPA Lysis buffer、2.5 μL PMSF和2.5 μL磷酸酶抑制剂的混合液,混匀后冰上裂解30 min,13 000 r/min离心10 min,收集上清,用BCA试剂盒测蛋白浓度。加SDS-PAGE Loding buffer,煮样备用。然后转膜、封闭、一抗孵育、二抗孵育,最后进行蛋白检测,保存图片。
2 结果
2.1 BAB-RS25305基因PCR扩增
BAB-RS25305基因以灭活的布鲁氏菌全基因组DNA为模板,经过PCR扩增后,琼脂糖凝胶电泳检测,发现一条大小为450 bp左右的特异性目的条带,与预期片段大小一致(图1)。
2.2 BAB-RS25305重组质粒的双酶切
将构建好的pET-32a-BAB-RS25305重组质粒用Hind Ⅲ、EcoRⅠ进行双酶切鉴定,经过10 g/L琼脂糖凝胶电泳检测,发现2条条带(图2),大小约为5 900 bp系pET-32a(+)载体条带,450 bp左右系目的基因条带。公司测序结果与GenBank中BAB-RS25305基因序列同源性达到100%,表明pET-32a-BAB-RS25305表达载体构建成功。

M.DNA 标准DL 2 000;1~5.BAB-RS25305基因PCR产物;6.阴性对照

M.DNA 标准DL 10 000;1、2.pET32a-BAB-RS25305Hind Ⅲ、EcoRⅠ的双酶切鉴定;3.阴性对照
2.3 BAB-RS25305重组蛋白的诱导表达以及纯化
将构建好的pET-32a-BAB-RS25305表达载体转化至大肠埃希氏菌感受态细胞DE3中进行诱导表达,在活化菌液中加入IPTG,分别在0、2、4、6、8 h收集菌液,并设置空菌以及空载,进行SDS-PAGE检测,BAB-RS25305蛋白大小约为32 ku,并且在6 h时表达量最大(图3),经过对BAB-RS25305蛋白进行可溶性分析结果显示,BAB-RS25305蛋白在主要以包涵体的形式进行表达,利用His标签蛋白纯化试剂盒(包涵体蛋白)进行纯化(图4)。
2.4 BAB-RS25305蛋白对巨噬细胞NLRP3和caspase-1 mRNA水平表达的影响
根据荧光定量PCR检测细胞中炎症因子结果显示,在BAB-RS25305重组蛋白刺激小鼠巨噬细胞24 h后,BAB-RS25305蛋白试验组NLRP3的表达量极显著高于PBS对照组(P<0.01),caspase-1 mRNA表达量显著高于PBS对照组(P<0.05)(图5)。
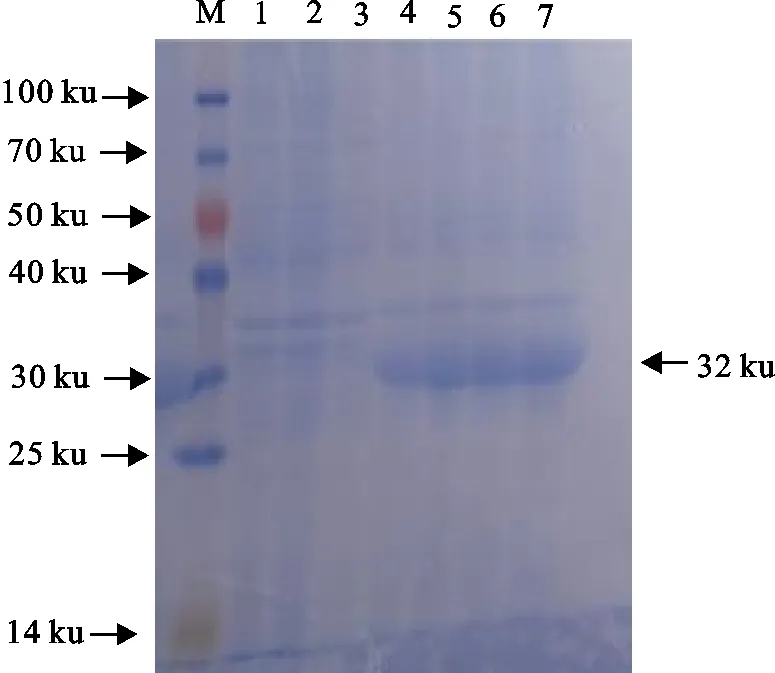
M.蛋白分子质量标准;1.E.coli BL21(DE3)阴性对照;2.PET-32a空载体诱导产物;3~7.表达菌诱导0、2、4、6、8 h

M.蛋白分子质量标准;1~3.纯化后rBAB-RS25305蛋白
2.5 BAB-RS25305蛋白对巨噬细胞NLRP3和caspase-1蛋白水平表达的影响
根据Western blot检测结果,与PBS对照组相比,BAB-RS25305蛋白刺激小鼠巨噬细胞24 h后NLRP3和caspase-1的蛋白表达量都极显著的增加(P<0.01)(图6)。
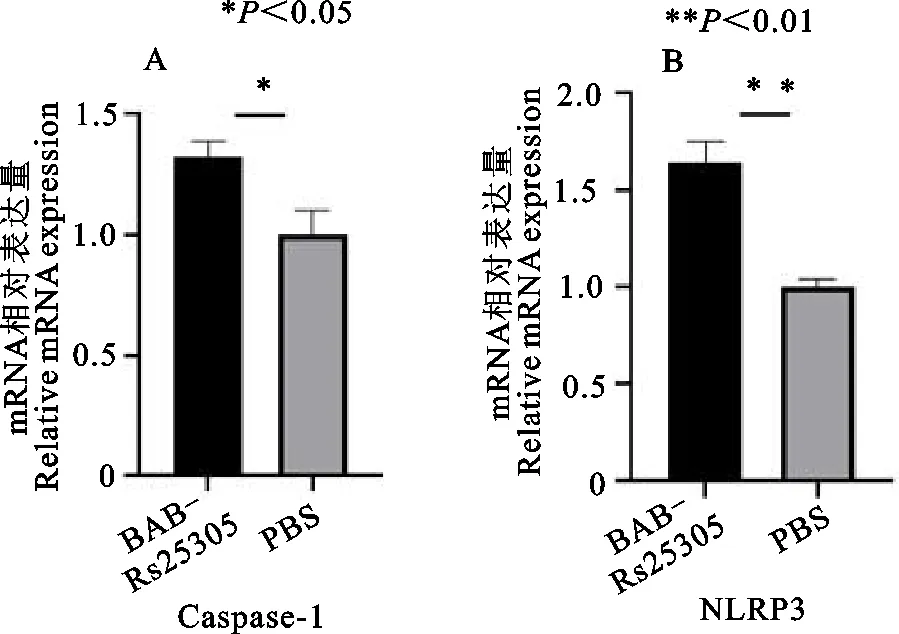
A.NLRP3;B.caspase-1
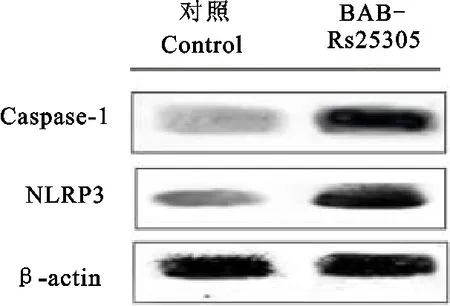
图6 Western blot检测炎症因子NLRP3和caspase-1在24 h时蛋白的表达量
3 讨论
布鲁氏菌是一种兼性的胞内病原体,可抵抗中性粒细胞的杀伤,也可在巨噬细胞和“非专业”吞噬细胞中复制,并与宿主细胞保持长期相互作用[13]。布鲁氏菌的一个关键致病特征是它们在免疫细胞中生存并在网状内皮系统的组织中持续存在的能力。与其他革兰氏阴性菌相比,一旦感染布鲁氏菌并没有立即导致受侵袭部位产生炎症,而是在出现症状之前有一个2周~4周的潜伏期[5],所以布鲁氏菌在黏膜表面和全身扩散的过程中,必须逃避先天免疫来躲避宿主检测,进而在细胞中大量的增殖[14-15],因此,更好地了解布鲁氏菌与宿主免疫系统之间的相互作用,对研究布鲁氏菌的致病机制以及治疗方法尤为重要。
宿主先天性免疫系统的第一个防御机制是通过模式识别受体(pattern recognition receptor,PRR)感应病原体,并通过产生细胞因子和抗微生物中间体迅速做出反应[16]。由PRR介导的微生物可识别病原体相关的分子模式,保守的微生物成分[17]和源于危险的信号(危险相关的分子模式)[18]。最常见的PAMP是细菌脂蛋白、肽聚糖、LPS、鞭毛蛋白,以及源自细菌、病毒、真菌和原生动物的核酸[19]。而模式识别受体参与诱导促炎性细胞因子和Ⅰ型干扰素产生的信号传导途径,这对于宿主抵抗各种病原体是非常重要的。NLRP3炎症小体作为布鲁氏菌侵袭细胞的主要模式识别受体,它属于NOD样受体家族(nucleotide binding oligomerization domain like receptors,NLR),其激活一部分与布鲁氏菌感染诱导的线粒体活性氧(reactive oxygen species,ROS)产生有关,ROS的产生诱导硫氧还蛋白(thioredoxin,TRX)相互作用蛋白(thioredoxin interacting protein,TXNIP)与NLRP3相互作用,导致其激活释放caspase-1,最终产生IL-1β[20-21]。布鲁氏菌激活NLRP3炎症小体的另一条途径是布鲁氏菌在感染期间诱导的内质网应激激活了未折叠蛋白反应(unfolded protein response,UPR)的肌醇需求酶1ɑ(inositol-requiring transmembrane kinase /endonuclease-1 ɑ,IRE1ɑ)途径,导致TXNIP和NLRP3募集到线粒体,在那里它们以一种独立于caspase-1的机制参与线粒体ROS的产生[22]。但是有研究表明,炎性小体过多的激活可能会导致一些与神经布鲁氏菌病症状相关的疾病[23]。本试验首先对布鲁氏菌BAB-RS25305基因进行原核表达,将纯化好的布鲁氏菌BAB-RS25305蛋白刺激小鼠巨噬细胞导致NLRP3和caspase-1的表达水平都显著提高。说明BAB-RS25305蛋白刺激巨噬细胞后,引起了NLRP3炎性小体的激活来识别布鲁氏菌病原体相关分子模式,进而活化caspase-1,从进下游细胞因子的分泌与成熟产生。但是BAB-RS25305蛋白刺激后NLRP3的激活是与线粒体ROS的产生有关还是通过未折叠蛋白反应的IRE1ɑ途径,目前还不清楚。
总之,本研究证明布鲁氏菌BAB-RS25305蛋白侵入宿主细胞后,炎症因子NLRP3和caspase-1的表达显著提高,进一步说明此蛋白可能参与炎症反应,为研究布鲁氏菌的胞内生存以及致病机制提供了理论依据。但是BAB-RS25305蛋白刺激小鼠巨噬细胞RAW264.7炎症因子的释放的机制,以及炎症反应的发生是否与未折叠蛋白反应有关还需要进一步研究。
猜你喜欢
食品安全导刊(2021年21期)2021-08-30
现代畜牧科技(2021年4期)2021-07-21
科学(2020年3期)2020-11-26
当代水产(2020年3期)2020-06-15
天津医科大学学报(2019年6期)2019-08-13
现代检验医学杂志(2016年2期)2016-11-14
中国民族医药杂志(2016年9期)2016-05-09
小星星·阅读100分(高年级)(2015年11期)2015-11-28
医学研究杂志(2015年12期)2015-06-10
当代畜禽养殖业(2014年5期)2014-08-31
