
zeste基因同源蛋白2在肿瘤免疫中的研究进展
2021-09-15 01:12:16开姚启盛付圣赐
中国比较医学杂志 2021年8期
尚 开姚启盛付圣赐
(1.湖北医药学院,湖北 十堰 442000;2.太和医院泌尿外科,湖北 十堰 442000)
表观遗传学让我们认识到子代表型取决于亲代基因的局限性,组蛋白修饰、DNA甲基化和非编码RNA等表观遗传在肿瘤的发生过程中起重要作用。细胞的生长发育同时受到环境因素影响,特定环境中表观的遗传特征甚至表现出可逆潜能[1-2]。EZH2通过对组蛋白甲基化修饰,激活或抑制相关基因表达导致肿瘤发生,研究发现其还参与组成肿瘤微环境(tumor microenvironment,TME)干扰破坏免疫稳态而促进肿瘤进展,在不同免疫细胞中差异表达发挥不同的生理作用[3-4]。本文就EZH2在TME中如何调节免疫功能及其免疫治疗的意义作一综述。
1 EZH2基因及其生物学功能
EZH2是重要的表观遗传调控因子,是哺乳动物细胞中组蛋白H3赖氨酸27(H3K27)甲基化的两种必需的催化酶之一,哺乳动物中已发现了两个PcG复合物家族:PRC1(polycomb-complex 1)和PRC2。PRC1复合物由多个亚基组成,如PCGF、HPH、CBX和RING1同源基团组成,催化组蛋白H2A的单双基化;而PRC2复合物主要由EED、SUZ12、RbAp46/48、EZH1或EZH2组成,参与H3K27的甲基化[5]。EZH2作为PRC2的催化亚基,是PRC2体外酶活性所必需的组成部分,其基因定位于7号染色体长臂7q35,包含20个外显子,编码746个氨基酸残基。EZH2包括几个结构域:SET保守结构域(位于C端维持组蛋白甲基转移酶活性)、CXC结构域和ncRBD结构域(与其他PRC2成分和调控蛋白相互作用所必需)[6]。EZH2修饰组蛋白H3K27甲基化(H3K27me3)是一种转录抑制的表观遗传标记,参与维持干细胞特性(见表1),调控基因表达、细胞周期、细胞分化和组织器官的发育,H3K27me3可促进多种肿瘤的发生、发展和演变[7-9]。
2 EZH2在肿瘤微环境中的作用
TME影响肿瘤的发生、发展和演变,而EZH2可以调控TME不同组分的功能。肿瘤微环境不仅包括肿瘤细胞、细胞外基质、成纤维细胞、内皮细胞和脂肪细胞,还包括T细胞、B细胞、调节性T细胞(Treg)、NK细胞、树突状细胞(DCs)和肿瘤相关巨噬细胞(TAMs)等免疫细胞[4,10]。在TME中,肿瘤细胞、免疫细胞、多种细胞因子和趋化因子相互作用形成免疫稳态[4]。EZH2在肿瘤微环境中主要有两种免疫调节功能:维持慢性炎症和建立免疫耐受[5]。慢性炎症可诱导表观基因重新编程,肿瘤细胞基因和表观遗传参与形成TME,致使肿瘤发生转化,同时慢性免疫细胞与肿瘤细胞相互作用,进一步抑制抗肿瘤免疫反应,最终促进肿瘤进展[5,11]。在前列腺癌细胞中,EZH2激活PI3K-AKT信号通路介导其在S21位点发生磷酸化,致使EZH2复合体从抑制因子转变为激活因子,通过IL-6、IL-8和TNF等反作用激活NF-κB靶基因,形成慢性炎症-NFκB-EZH2通路循环并促进肿瘤进展[12]。卵巢癌中靶向EZH2发现相关microRNA表达增加,帮助肿瘤细胞逃避免疫监测[13];弥漫大B细胞淋巴瘤中EZH2基因易发生突变,导致MHC-II获得性缺陷,参与肿瘤免疫逃逸[14]。恶性黑色素瘤肿瘤内T细胞浸润和INF-α可使微环境中EZH2表达上调,EZH2甲基化H3K27me3可抑制MHC-I、免疫蛋白酶体、细胞引诱趋化因子等抗原提呈相关蛋白,并沉默肿瘤细胞的免疫原性[15](见表1)。一系列报道显示EZH2在肿瘤免疫微环境中的作用正在不断被发现,如免疫选择、免疫编辑、免疫监测、免疫维持、免疫逃逸等[16],研究其在不同时期不同免疫细胞类型中的调节功能,为指导临床靶向EZH2的免疫治疗具有重要意义。
3 EZH2在免疫细胞中的作用
在TME中,EZH2过表达或缺失均可干扰免疫稳态,其在多种免疫细胞中表达并发挥不同的生理作用。
3.1 T细胞
初始T细胞识别肿瘤抗原,分化为效应性T细胞,趋化至肿瘤组织,形成浸润性淋巴细胞发挥效应,但其寿命短,大部分在发挥效应后凋亡,有一少部分可快速增殖、分化成效应性T细胞,介导最主要抗肿瘤免疫反应,即细胞免疫。TME中记忆T细胞寿命长,可自我更新并在再次识别抗原时快速表达并发挥作用。EZH2通过调节T细胞分化、效应、趋化和凋亡等不同功能影响其抗肿瘤作用[17-18]。
CD8+T:EZH2对CD8+记忆性T细胞的发育和维持发挥关键作用。He等[18]人利用稳定表达卵清蛋白(OVA)抗原的黑色素瘤荷瘤发现,特异性敲除T细胞EZH2,降低OVA特异性记忆性T细胞数量。进一步研究发现EZH2可通过抑制ID2,PRDM1和EOMES等基因转录,增强H3K4me3和Pol II与ID3基因座结合,上调ID3转录而增强记忆性T细胞的分化。当TCR信号活化Akt,磷酸化EZH2,其活性被抑制,能增加效应性而减少记忆性T细胞。EZH2增强抗肿瘤免疫的能力还体现在能提高CD8+T细胞增殖活性和抵抗凋亡的能力增强。NUMB和FBXW7是Notch的主要负性调节因子,EZH2通过H3K27me3抑制2种阻遏物转录,从而激活T细胞Notch信号,增加细胞的增殖。抗凋亡基因Bcl-2也受EZH2调控。EZH2活化,上调Bcl-2信号抵抗凋亡,促进T细胞生存。Zhao等[13]发现,卵巢癌通过增加microRNA101和microRNA26a抑制浸润性CD8+T细胞中EZH2表达从而逃避免疫攻击,同时发现使用shRNA或抑制剂可干扰CD8+T细胞EZH2作用,能够抑制其抗肿瘤功能。还有学者发现CD8+T细胞长期暴露于抗原环境中易发生衰竭,表现为对抗原免疫反应性下降,进一步削弱抗肿瘤免疫[19]。
CD4+T:EZH2抑制CD4+初始T细胞分化成效应细胞。通过特异性性敲除T细胞EZH2发现,EZH2-/-CD4+初始T细胞体外分化成Th1、Th2和Th17等效应细胞增多,分泌的IFN-γ、IL13和IL-17等对应的效应分子明显增加。体内研究也有相同的结论,CD4CreEzh2Fl/FlC57在OVA接触实验中诱发的过敏性哮喘症状,病理特征更严重,进一步证实了EZH2对Th2分化的抑制作用。EZH2抑制CD4+T细胞分化可能与其沉默T-bet和Gata3等效应细胞关键转录因子相关,H3K27me3在IFNG、Il10等基因位点的调控也依赖PRC2-EZH2[20]。
这些证据均表明EZH2在T细胞中扮演重要角色,EZH2磷酸化维持记忆CD8+T细胞的抗肿瘤作用,与肿瘤细胞EZH2过表达提示其恶性程度较高是否矛盾?EZH2在CD4+T细胞中的作用尚未完全明确,TME中T细胞如何参与EZH1和EZH2之间的相互转换?这些问题仍有待进一步探讨。
3.2 Treg细胞
Treg细胞是一类特殊的CD4+T细胞,对维持免疫稳态和自我耐受至关重要。EZH2介导表观遗传对调节性T细胞(regulatory T cell,Treg)的募集和发挥其免疫抑制作用不可或缺[21]。EZH2在未成熟或休眠的Treg细胞中以基础速率表达,诱导激活后迅速表达。EZH2缺失的Treg细胞在突变小鼠模型中不能维持免疫稳态,表现出组织中浸润较多淋巴细胞,同时小鼠寿命缩短[22]。FOXP3是Treg的转录因子,FOXP3表达降低可导致EZH2缺失,EZH2与FOXP3相互作用激活Treg,EZH2在FOXP3结合位点的募集性降低,将导致H3K27me3沉积和基因抑制[21]。Yang等[21]评估EZH2对Treg功能的影响,在特异性缺失EZH2的转基因小鼠发现脾和淋巴结中幼稚T细胞明显减少,CD4细胞中FOXP3表达降低,INF-γ和IL-4抗体可逆转FOXP3的表达,可能机制是FOXP3在缺失EZH2的转基因小鼠体内表达减少将导致INF-γ和IL-4等细胞因子异常表达。因此,Treg中EZH2的表达在TME发挥重要调控作用[23],Treg失调如何引起效应T细胞功能紊乱仍是一大问题。
3.3 B细胞
EZH2在生发中心B细胞(GC B)中表达升高,在介导GC B细胞表型中发挥重要作用,是GC B细胞成熟和形成免疫球蛋白亲和力所必需的[24]。EZH2在B细胞各个发育和分化阶段的作用不可或缺。在B细胞前体阶段,EZH2协调IL-7响应能力、增殖和lgκ链重组;EZH2通过抑制浆细胞关键转录因子Prdm1和Irf4,使GC B细胞能够耐受AID介导的体细胞高频突变,防止末端过早分化,促进细胞增殖;在浆细胞中,EZH2与Blimp-1相互作用促进Xbp1和未折叠蛋白(UPR)反应以及特异性代谢,可抑制B细胞基因表达并提高免疫球蛋白产生率[24]。Guo等[25]也证实在EZH2缺失的情况下,抗体分泌细胞中组蛋白H3赖氨酸27三甲基化基因将发生异常调控,致使抗体分泌细胞的成熟、新陈代谢和功能发生紊乱。TME中EZH2调节B细胞功能发挥关键作用,如何促进B细胞成熟、参与形成免疫球蛋白及分泌仍有待深入研究。
3.4 NK细胞
NK细胞是先天免疫系统的组成部分,能够不经过MHC-I抗原提呈对各种外来抗原、感染和肿瘤细胞迅速作出反应[26]。研究表明EZH2可通过表观遗传机制调节NK细胞的发育参与肿瘤免疫。造血干细胞中CD122、NKG2D、TOLL样受体和NK细胞颗粒酶可促进NK细胞及其前体细胞发育增殖,抑制EZH2后均表达增加,增强抗肿瘤作用[27-28]。NK细胞中靶向EZH2免疫治疗可能是抗肿瘤治疗的一种有效方法。
3.5 DC细胞
树突状细胞(DC)是抗原呈递细胞,是连接先天免疫系统和适应性免疫之间的重要桥梁。Tian等[29]报道EZH2过表达在多数组织细胞和树突状细胞肿瘤的成瘤过程中发挥重要作用,如组织细胞肉瘤、滤泡性树突状细胞肉瘤、朗格汉斯细胞增多症和树突状细胞肉瘤。Gunawan等[30]也证实EZH2调节DCs整合素信号转导和粘附动力学,促进自身免疫性脑脊髓炎的发生。这些均表明EZH2在DC中发挥重要调控作用,但其机制尚未完全清楚。
3.6 TAM细胞
巨噬细胞是在造血系统中形态表现多样,具有维持体内稳态、组织修复和免疫调节等多种功能,所谓肿瘤相关巨噬细胞(TAMs)指可促进肿瘤细胞增殖和存活或者调节免疫抑制微环境而促进肿瘤进展一类巨噬细胞[31-32]。Qiao等[33]研究发现EZH2介导H3K27me3甲基化,沉默MERTK、PPARG等基因,使IFN-γ诱导巨噬细胞活化。另有学者也提出小胶质细胞中EZH2通过抑制细胞信号抑制因子3(Socs3)可导致自身免疫性炎症,EZH2缺失直接刺激Socs3表达,从而增强肿瘤坏死因子受体相关因子6的降解,通过建立小胶质细胞EZH2缺失的小鼠模型发现沉默Socs3可重新激活小胶质细胞的功能并抑制自身免疫性炎症[32]。然而TAM中EZH2失调能够促进肿瘤进展的机制仍然不明。
3.7 其他细胞
PLZF(promyelocytic leukemia zinc finger)是EZH2甲基化和降解的靶点,EZH2通过调节PLZF浓度,使胸腺产生的iNKT(invariant natural killer T)细胞数量维持在适当水平,促进免疫稳态。EZH2的缺失使PLZF激活IL-4,促进NKT细胞扩增,破坏TME适应性免疫[34]。通过建立EZH2缺陷的小鼠模型也显示其中NKT2细胞数量增加,并导致免疫稳态紊乱和出现哮喘样病理改变;还有报道EZH2缺失将损害朗格汉斯细胞(langerhans cells,LCs)的粘附结构,导致迁移基底膜受阻,参与调节皮肤耐受[35-36]。
这些结果均表明肿瘤微环境免疫改变将破坏免疫稳态可促进肿瘤发生。EZH2在肿瘤和各种免疫细胞之间起着关键作用,对自身免疫、异源反应和抗肿瘤免疫的调控有重要意义。随着研究的深入我们可能发现新的治疗策略,提高免疫治疗的疗效。
4 EZH2的免疫治疗
大量证据表明EZH2通过表观遗传参与抗肿瘤免疫和免疫逃逸,其相关的免疫治疗如免疫检查点阻滞剂和EZH2特异性抑制剂逐渐成为活跃的研究领域,越来越多以EZH2为靶点的药物被相继报道,并在体外细胞实验、体内临床前模型实验和患者临床试验中显示出对各种恶性肿瘤的抗肿瘤作用。
免疫检查点是指免疫系统存在大量抑制通路,这些通路对于维持自我耐受和调节外周组织生理免疫反应强度及持续时间,减少组织损伤至关重要[37]。免疫检查点阻滞剂,包括抗CTLA-4抗体,GITR抗体、抗PD-1(PDCD1)抗体和抗PDL-1(CD274)抗体是抗肿瘤免疫治疗方法之一[38]。有报道在结直肠癌中,靶向抑制EZH2可以增强CXCL9和CXCL10趋化因子的表达,阻止效应T细胞向TME的转运[39]。通过小鼠肿瘤模型中发现EZH2表达升高可抑制T细胞抗肿瘤免疫,抗CTLA-4免疫治疗和靶向抑制EZH2可增强抗肿瘤免疫反应,改善抗肿瘤作用[21,40]。
此外,以EZH2为靶点的抑制剂的研究亦是热点之一,EZH2抑制剂能够竞争性抑制S-腺苷-L-同型半胱氨酸水解酶(SAH),增加细胞SAH水平,从而抑制S-腺苷-L-蛋氨酸依赖组蛋白赖氨酸甲基转移酶活性,可显著降低H3K27me3水平,具有明显抗肿瘤增殖作用[7]。随着研究的深入,新型EZH2抑制剂如GSK126[7]、GSK343[41]、GSK503[41]、EI1[7]、CPI-1205[42]、+UNC1999[7]、Constellation Compound 3[7]、OR-S1[43]、OR-S2[43]、Tazemetostat[44](见表1)等相继被发现,特别是在EZH2高表达的肿瘤中应用前景极具吸引力,研究显示抑制EZH2有助于逆转抗肿瘤免疫,但同时在一定时间内也可降低免疫细胞的生存[39](见表1)。报道指出EZH2在抗肿瘤免疫中对肿瘤细胞和免疫细胞表现出截然相反的作用,所以此方法对抗肿瘤免疫影响尚未完全明确,EZH2对免疫细胞的作用可能取决于肿瘤类型、TME和治疗方法[15,45]。
此外,任何单一药物治疗都可使肿瘤细胞耐药,对EZH2抑制剂的耐药机制已有报道。在细胞系模型研究中,长期暴露于EZH2抑制剂的细胞中发现EZH2的两个新的次级突变(Y111L和Y661D)可产生协同作用导致细胞耐药[46]。弥漫大B细胞淋巴瘤中IGF-1R,PI3K和MAPK通路的激活亦可促进对EZH2抑制剂的耐药[14]。最新研究指出,复发性头颈部鳞状细胞癌中靶向EZH2可以增强抗原呈递,GSK126联合抗PD-1治疗可以规避抗PD-1耐药,改善药物敏感性[47](见表1);亦有报道恶性黑色素瘤中EZH2失活与CTLA-4抗体和IL-2联合治疗具有协同作用[15]。这些结果表明免疫、基因、内分泌等联合治疗可能是未来抗肿瘤的一种方法。
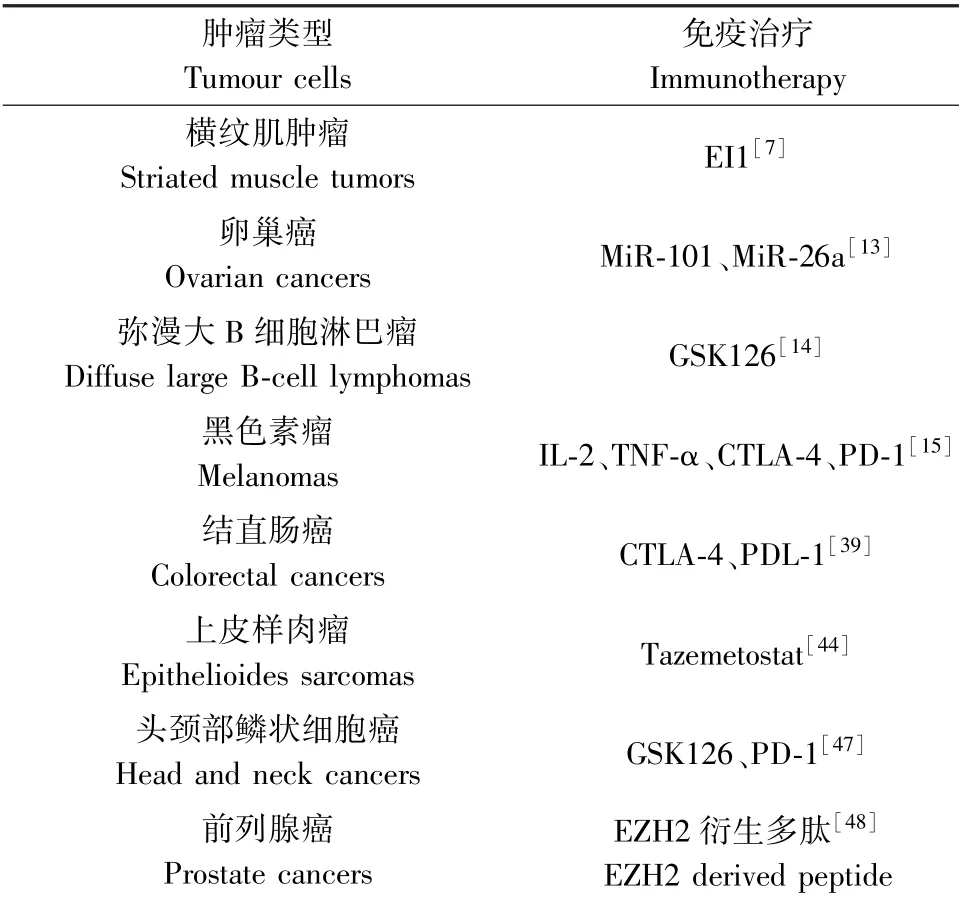
表1 临床免疫治疗结果Table 1 Clinical immunotherapy results
5 结论与展望
大量研究已表明,EZH2是表观遗传肿瘤相关的重要调节因子,其在肿瘤免疫中的作用机制尚不明确,对靶向EZH2免疫治疗提出了新的挑战。EZH2既可以作为肿瘤抑制因子,也可以作为肿瘤启动因子,取决于EZH2发挥功能的细胞环境。EZH2在肿瘤中功能也可能仅仅反映了在肿瘤的不同阶段该蛋白所扮演的作用,即在肿瘤发生起始阶段表达抑制因子,当细胞环境改变时,表观遗传学参与基因调控后成为肿瘤启动因子。了解EZH2在TME复杂的作用机制应是今后研究的重点,将EZH2抑制剂与免疫疗法相结合可能是一有效途径,但需同时考虑其对宿主和肿瘤免疫的潜在影响,这些都有待进一步研究证实。明确EZH2在TME、免疫细胞、免疫应答和抗肿瘤免疫中作用,为以后药物研发及指导临床肿瘤靶向免疫治疗具有重要意义。
猜你喜欢
大电机技术(2022年3期)2022-08-06 07:48:24
核科学与工程(2021年4期)2022-01-12 06:30:04
河北果树(2021年4期)2021-12-02 01:14:50
煤气与热力(2021年4期)2021-06-09 06:16:54
中国生殖健康(2020年7期)2020-12-10 07:48:51
中华戏曲(2020年1期)2020-02-12 02:28:18
上海公路(2019年3期)2019-11-25 07:39:28
福建基础教育研究(2019年10期)2019-05-28 08:27:04
海南医学(2016年8期)2016-06-08 05:43:00
遗传(2014年3期)2014-02-28 20:58:52
