
几种姜黄属植物光合特性研究
2021-09-12 00:47李莹彭玉德余丽莹陈晓英潘春柳蓝祖栽黄雪彦
江苏农业科学 2021年14期
关键词:光合特性
李莹 彭玉德 余丽莹 陈晓英 潘春柳 蓝祖栽 黄雪彦


摘要:为了解几种不同姜黄属植物的光合特性及其对环境的适应能力,以便更好地对姜黄属植物资源进行保护与利用,采用LI-6400XT便携式光合作用测量系统(LI-COR,USA),对几种处于生长旺盛期的姜黄属植物进行光合日变化、光强度-光合速率曲线和CO2-光合速率曲线等光合指标测定,并对比分析其光合特性的异同。结果表明,几种供试姜黄属植物的光合日变化参数、光响应参数和CO2响应参数均存在较大差异。随着时间的推移,几种姜黄属植物的净光合速率(Pn)日变化都呈现出先增大后减小的“单峰”曲线变化规律,均未出现“午休”现象。净光合速率大小为姜黄>广西莪术>顶花莪术>川郁金>温郁金>大莪术>郁金,且各植物净光合速率达到峰值的时间不同,气孔导度、胞间CO2浓度及蒸腾速率亦存在显著性差异;几种姜黄属植物的光补偿点(LCP)均小于45 μmol/(m2·s),差异不显著,而光饱和点(LSP)在350~1 350 μmol/(m2·s)之间,差异较大;CO2饱和净光合速率均在14~23 μmol/(m2·s)之间,差异不显著,但羧化效率、CO2饱和点和CO2补偿点均存在显著性差异。研究表明,供试7种姜黄属植物均具有较强的耐阴特性,对弱光利用能力较强,对强光也有一定的耐受性,但不同植物对光适应范围以及对强光利用能力存在一定的差异。应结合不同姜黄属植物的光合特性,适当调控其生长环境因子,根据每种植物对光照适应性采取适当遮阴或向阳措施,并适当增加环境中CO2浓度,进一步提高其光合利用效率和增加药材产量。
关键词:姜黄属;光合特性;日变化;光响应曲线;CO2响应曲线
中图分类号: S567.23+9.01 文献标志码: A 文章编号:1002-1302(2021)14-0143-06
姜黄属(Curcuma L.)植物隶属于姜科(Zingiberaceae),为多年生草本,其膨大块茎和块根均可供药用,中药材姜黄、郁金和莪术的基源植物即来自于姜黄属[1-2]。现代医学研究表明,姜黄属植物具有抗炎、抗肿瘤、降血脂和抗氧化等作用[3]。此外,姜黄属植物也具有较高的观赏价值[4-5],有些还可开发作为调味品、色素、香料、染料、化妆品和杀虫剂等[6]。近年来关于姜黄属植物的研究主要集中在化学成分[7-12]、药理活性[13-15]、分类鉴别[16-18]等方面,关于其栽培生理,尤其是光合特性研究报道较少。
植物光合作用是植物体内最重要的生命活动过程,是生物链的初级生产者,地球上几乎所有的有机物都直接或间接地来源于光合作用。光合作用在植物的生长发育过程中发挥着重要作用,是植物光合物质生产和产量形成的重要影响因素之一,在一定程度上决定了植物生长发育和生产能力的高低,植物对环境因子的适应性也可以通过测定光合特征参数的变化反映出来[19-20]。因而,了解植物的光合生理特性是植物引种栽培过程中的基础性工作,可以为植物的引种栽培和开发利用提供参考依据。
本研究对7种姜黄属植物的光合日变化、光强度-光合速率曲线和CO2-光合速率曲线等相关指标进行测定,并对比分析了其光合特性的异同及其对环境的适应能力。研究结果可为深入了解姜黄属植物的光合生理差异及其对环境的适应性提供基础数据,并为姜黄属植物资源的迁地保护与可持续开发利用提供理论基础。
1 材料与方法
1.1 试验材料
本试验所用材料为7种姜黄属植物,分别为姜黄(C. longa Linnaeus)、广西莪术(C. kwangsiensis S. G. Lee & C. F. Liang)、郁金(C. aromatica Salisb.)、温郁金(C. aromatica Salisb. ‘Wenyujin)、川郁金(C. sichuanensis X. X. Chen)、大莪术(C. elata Roxb. )和顶花莪术(C. yunnanensis N. Liu & S. J. Chen)。以上材料均来源于广西药用植物园科研基地,于2019年2—3月选择大小一致、无病虫害且饱满结实的根茎进行种植栽培,种植土为土层深厚、排灌良好的沙壤土,种植后进行统一栽植管理。
1.2 试验方法
1.2.1 光合日变化测定 于供试姜黄属植物的生产旺盛期(2019年7—8月),选择晴朗天气,采用 LI-6400XT 便携式光合作用测量系统(LI-COR,USA),測定几种供试姜黄属植物的光合日变化,每天08:00—18:00,每隔2 h测定1次,重复测定 3 d,取平均值。测定时每种植物随机选取生长健壮、长势一致、光照均一的植株10株,以植株主茎自上向下第1张成熟稳定叶片作为测试叶,记录叶片的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)等光合参数及叶片表面温度(CTleaf)、环境CO2浓度(CO2R)、大气湿度(RH_R)和外界光合有效辐射(PARi)等环境参数。为了消除时间上的误差,测定时几种供试植物随机排序。
1.2.2 光强度-光合速率曲线测定 于供试姜黄属植物的生产旺盛期(2019年7—8月),选择天气晴朗且植物光合作用较为旺盛的时间段(09:30—11:30),采用LI-6400XT便携式光合作用测量系统(LI-COR,USA),测定几种供试姜黄属植物的光强度-光合速率曲线。测定时每种植物随机选取植株3株,测试叶片选取方法同日变化测定。测定时设置CO2浓度为400 μmol/(m2·s),叶室温度为 25 ℃。使用LI-6400XT的LED红/蓝光源控制光照强度,光强辐射梯度设定为0、20、50、100、150、200、400、600、800、1 000、1 200 μmol/(m2·s),使用LED红/蓝光源叶室测定净光合速率(Pn)。每次测定前将待测叶片在1 000 μmol/(m2·s)的光照强度下诱导 20 min。重复测定3 d,取平均值。将测得的数据采用双曲线修正模型进行回归拟合,得到最大净光合速率(Pmax)、暗呼吸速率(Rd)、表观量子效率(AQY)、光补偿点(LCP)和光饱和点(LSP)。
1.2.3 CO2-光合速率曲线测定 于供试姜黄属植物的生产旺盛期(2019年7—8月),选择天气晴朗且植物光合作用较强的时间段(09:30—11:30),采用LI-6400XT便携式光合作用测量系统(LI-COR,USA),测定几种供试姜黄属植物的CO2-光合速率曲线。测定时每种植物随机选取植株3株,测试叶片选取方法同日变化测定。测定时设置光强为该植物的饱和光强(根据光强度-光合速率曲线确定),叶室温度设定为25 ℃。利用CO2钢瓶控制CO2浓度(Ci),CO2浓度梯度设定为400、300、200、100、50、400、400、600、800、1 000、1 200、1 400、1 600、1 800 μmol/(m2·s),使用LED红/蓝光源叶室测定净光合速率(Pn)。重复测定3 d,取平均值。将测得的数据采用双曲线修正模型进行回归拟合,得出CO2饱和净光合速率(CSPn)、羧化效率(CE)、CO2补偿点(CCP)和CO2饱和点(CSP)。
1.3 数据分析
用Excel 2013进行数据整理和作图,SPSS 22.0软件进行数据差异显著性分析,光合计算软件对各响应曲线的参数进行拟合及数学模拟。
2 结果与分析
2.1 光合日变化
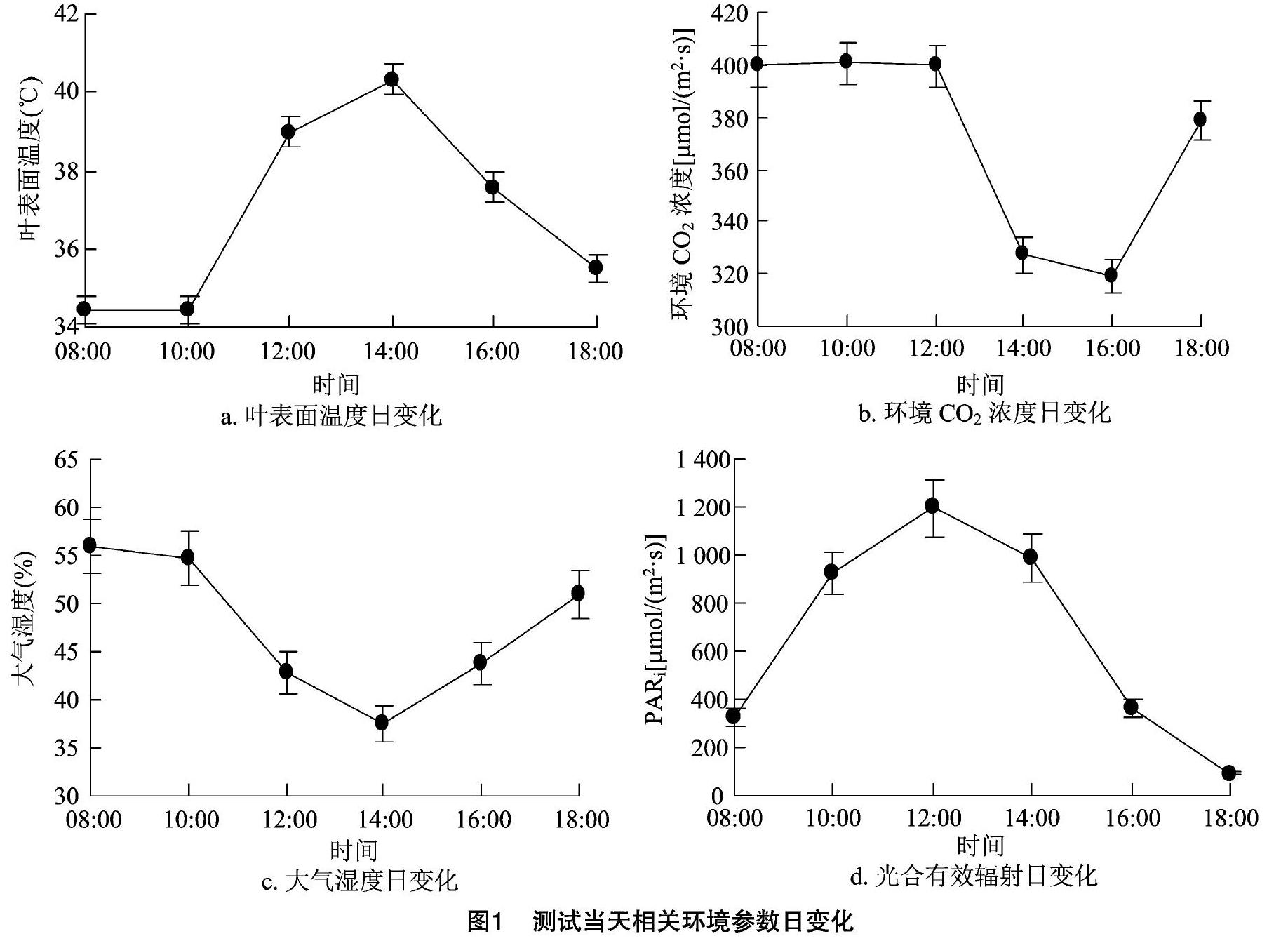
2.1.1 相关环境参数日变化 如图1所示,测试当天,随着时间的推移,植物叶片表面温度和光合有效辐射均呈现出先增大后减小的变化规律,光合有效辐射的最大值出现在12:00,叶表面温度最大值出现在14:00。而大气湿度刚好相反,随着时间的推移,大气湿度先逐渐减小,14:00达到最小值,而后又逐渐上升。上午环境CO2浓度基本保持不变,均保持在400 μmol/(m2·s)左右,午后环境CO2浓度逐渐降低,16:00达到最小值,而后又逐渐上升至380 μmol/(m2·s)左右。这与测试当天气象特征基本吻合。
2.1.2 叶片光合参数日变化 由表1可见,随着时间的推移,几种姜黄属植物的净光合速率(Pn)日变化都呈现出先增大后减小的“单峰”曲线变化规律,均未出现明显的“午休”现象。净光合速率大小为姜黄>广西莪術>顶花莪术>川郁金>温郁金>大莪术>郁金,且各植物净光合速率达到峰值的时间不同,姜黄、广西莪术、温郁金、川郁金和顶花莪
术的净光合速率最大值出现在14:00,最大值均大于17 μmol/(m2·s),姜黄、广西莪术的最大净光合速率甚至超过20 μmol/(m2·s),分别为20.79、20.18 μmol/(m2·s);而郁金和大莪术的净光合速率最大值则出现在16:00,仅分别为11.14、12.31 μmol/(m2·s)。另外,温郁金和大莪术的净光合速率在18:00仍相对较高,均在10 μmol/(m2·s)以上。
由表1还可见,随着时间的推移,姜黄、广西莪术和温郁金的气孔导度(Gs)基本维持在0~0.15 mol/(m2·s) 之间,变化较平稳。大莪术的气孔导度则随着时间的推移呈现上升—下降—上升—下降—上升的趋势,第1个峰值出现在 10:00,第2个峰值出现在14:00,峰值均在 0.3 mol/(m2·s) 左右。川郁金和顶花莪术的气孔导度则随着时间的推移,基本呈现先增大再减小的单峰曲线,不同的是,顶花莪术的峰值出现在 10:00,为0.92 mol/(m2·s),而川郁金的峰值出现在12:00,为0.59 mol/(m2·s)。郁金的气孔导度则是先减小,在10:00达到最小值,为 -0.64 mol/(m2·s),后逐渐上升并出现平稳。
几种姜黄属植物的胞间CO2浓度随着时间推移变化趋势不尽相同。广西莪术呈现先减小后增大的变化趋势,在14:00达到最小值,为 267.07 μmol/mol。郁金、大莪术和顶花莪术的胞间CO2浓度随着时间推移,均呈现先减小后增大再减小的变化趋势,但其达到最大值的时间不同,郁金在08:00达到最大值,顶花莪术在14:00达到最大值,而大莪术的最大值则出现在16:00。温郁金的胞间CO2浓度变化较大,呈现先迅速减小,后迅速上升,再逐渐减小的趋势,其最小值出现在12:00,为107.64 μmol/mol,于14:00迅速上升至最大值,为446.01 μmol/mol,而后逐渐降低。姜黄和川郁金的胞间CO2浓度随着时间推移,呈现先减小后增大再减小再增大的变化趋势,最大值均出现在14:00。
几种姜黄属植物的蒸腾速率(Tr)亦存在较大差异。姜黄和广西莪术的蒸腾速率呈现出先增大再减小再增大再减小的“M”型“双峰”曲线变化趋势,分别在10:00和14:00出现峰值;温郁金和大莪术则表现出先增大再减小的“单峰”曲线变化趋势,均在12:00达到最大值;郁金、川郁金和顶花莪术的蒸腾速率最大值均出现在早上08:00,分别为3.81、4.58、6.43 mmol/(m2·s),川郁金和顶花莪术蒸腾速率随时间推移逐渐减小,在傍晚18:00达到最小值,且最小值均小于1 mmol/(m2·s),而郁金的蒸腾速率在中午12:00降到1.99 mmol/(m2·s)后又略有回升,14:00达到2.50 mmol/(m2·s),然后又迅速回落,18:00蒸腾速率甚至下降为负值,为 -0.62 mmol/(m2·s)。
采用SPSS 22.0数据分析软件对供试的7种姜黄属植物叶片净光合速率(Pn)与气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)进行相关性分析,结果详见表2。由表2可见,在这7种植物中,除郁金Pn与Gs呈不显著正相关外,其他几种姜黄属植物Pn与Gs均呈不显著负相关;除广西莪术Pn与Tr呈不显著正相关外,其余6种Pn与Tr呈现不显著的负相关;此外还可以看出,姜黄(CL)、温郁金(CAW)、川郁金(CS)、顶花莪术(CY)这4种植物的Pn值与Ci值呈现不显著正相关,其他3种的Pn值与Ci值均呈不显著负相关。
2.2 光强度-光合速率曲线
如表3所示,几种姜黄属植物的光补偿点均小于45 μmol/(m2·s),差异不显著,而光饱和点则存在显著性差异,大莪术和顶花莪术的光饱和点均大于1 300 μmol/(m2·s),姜黄、广西莪术和川郁金的光饱和点在780~900 μmol/(m2·s)之间,而郁金和温郁金的光饱和点则小于600 μmol/(m2·s),温郁金的光饱和点最低,仅为369.81 μmol/(m2·s)。几种姜黄属植物表观量子效率(AQY)最大的是大莪术,为0.09 μmol/μmol,最小的是广西莪术,仅为0.01 μmol/μmol,其余几种植物的表观量子效率主要集中在 0.05~0.08 μmol/μmol 之间,差异不显著。姜黄和川郁金的最大净光合速率(Pmax)相对较高,分别为9.83、10.85 μmol/(m2·s),广西莪术、郁金和温郁金的最大净光合速率较小,均低于 3.6 μmol/(m2·s)。几种姜黄属植物的暗呼吸速率(Rd)在0.1~2.2 μmol/(m2·s)之间,存在显著性差异,暗呼吸速率最大的为大莪术,暗呼吸速率为2.17 μmol/(m2·s),广西莪术的暗呼吸速率最小,仅为0.10 μmol/(m2·s)。
2.3 CO2-光合速率曲线
如表4所示,7种供试姜黄属植物CO2饱和净光合速率均在14~23 μmol/(m2·s)之间,差异不显著,但羧化效率、CO2饱和点和CO2补偿点均存在显著性差异。羧化效率最大的为大莪术,为0.034 μmol/μmol;郁金的羧化效率最小,为 0.018 μmol/μmol,姜黄、广西莪术、温郁金、川郁金和顶花莪术羧化效率均集中在0.024~0.029 μmol/μmol之间,差异不显著。CO2饱和点最大的为温郁金,为2 688.43 μmol/mol,最小的为广西莪术,仅为 1 461.72 μmol/mol;温郁金的CO2补偿点最大,为225.02 μmol/mol,姜黄、郁金次之,广西莪术、大莪术和顶花莪术的CO2补偿点较小,均小于 90 μmol/mol。
3 讨论与结论
光合作用是光合物质生产和产量形成的重要因素之一,在植物的生长发育过程中发挥着重要的作用。对植物的光合生理特性进行研究可以为植物的引种、栽培和开发利用提供参考依据[21]。植物光合能力的强弱不仅可以反映不同光照条件下植物生长的差异性,还能体现植物对环境变化的适应能力。植物光合特性为物种与生长环境相适应的表现[22]。本研究结果表明,几种姜黄属植物的净光合速率(Pn)日变化都未出现明显的“午休”现象,均呈现出先增大后减小的“单峰”曲线变化规律,且大多数植物的净光合速率达到峰值的时间为14:00,而此时叶片表面温度都超过40 ℃,并达到最大值,光合有效辐射亦接近1 000 μmol/(m2·s),说明姜黄属植物的光合生产力和光能利用率较高,且对高温和强光具有一定的耐受性。光合能力强弱在一定程度上取决于物种的遗传特性,此结果与供试的几种植物在分类上为同属植物,遗传表现上会出现一定的相似性相吻合。植物的光合能力强弱不仅可以反映不同光照条件下植物生长的差异性,同时也体现了其对环境變化的适应能力。几种供试姜黄属植物净光合速率最大值和达到峰值的时间皆有不同,气孔导度、胞间CO2浓度及蒸腾速率亦存在显著性差异,说明不同姜黄属植物对光能的利用率存在一定差异。这可能是因为供试植物由于产地生境差异较大,为适宜不同生境而衍生出各自特有的光合特性。此外,7种姜黄属植物叶片净光合速率与气孔导度、蒸腾速率、胞间CO2浓度均呈现不显著的正相关或负相关,这说明姜黄属植物的净光合速率受气孔导度、蒸腾速率、胞间CO2浓度等因素影响较小。
植物的光补偿点和光饱和点是评估其光适应性的重要指标。植物的光饱和点越高且光补偿点越低,则表示其对光的适应范围较大,光适应性较强[23]。表观量子效率(AQY)作为反映植物光合特性的指标,也可以反映出植物对光的利用能力大小,植物的AQY越低,其对光能的利用能力越弱[24]。通常耐阴植物的光补偿点与光饱和点均较低,它们能充分利用弱光进行光合作用;反之则属于阳性植物[25]。
本研究表明,供试的7种姜黄属植物的光补偿点均较低且差异不显著,最高者顶花莪术也仅为42.55 μmol/(m2·s),表明它们利用弱光能力均较强,表现出明显的耐阴特性,这也与彭昭良等研究结果姜黄属植物属于耐阴植物[26]相符。此外,供试的几种姜黄属植物对光适应范围有所不同,综合来看,大莪术、顶花莪术具有相对较低的光补偿点,相对较高的表观量子效率和最高的光饱和点,且两者光饱和点种间差异不显著,说明它们对光的适应范围最大,对强光适应能力强,因此大莪术和顶花莪术栽培上可适当增加光照强度,以提高其光能利用效率;姜黄、广西莪术、川郁金三者具有较低的光补偿点和相对较高的光饱和点,且三者光饱和点种间差异不显著,说明其对光的适应范围居中,对强光利用能力一般;郁金、温郁金两者的光补偿点和光饱和点相对最低,且差异不显著,说明其对光的适应范围较窄,对强光利用能力最弱,故郁金和温郁生产上可采取适当遮阴措施。
CO2是光合作用的重要原料,同时也是正常光照条件下光合作用的限制因子。本研究中,供试的7种姜黄属植物的CCP值在77.17~225.02 μmol/mol之间,其CSP值均大于1 461 μmol/mol,远高于大气中CO2浓度(320~400 μmol/mol),说明供试的7种姜黄属植物均能耐受较高的CO2浓度,对高浓度CO2利用能力较强,因而,适当增加环境中CO2浓度可在一定程度上提高姜黄属植物的光合利用能力。
综上所述,供试7种姜黄属植物均具有较强的耐阴特性,对弱光利用能力较强,对强光也有一定的耐受性,但不同植物对光适应范围以及对强光利用能力存在一定的差异,应结合不同姜黄属植物的光合特性,适当调控其生长环境因子,根据每种植物对光照适应性对其采取适当遮阴或向阳措施,并适当增加环境中CO2浓度,进一步提高其光合利用效率和增加药材产量。
参考文献:
[1]中国科学院《中国植物志》编辑委员会. 中国植物志:第十六卷第二分册[M]. 北京:科学出版社,1981:58-59.
[2]国家药典委员. 中华人民共和国药典:一部[M]. 2015版. 北京:中国医药科技出版社,2015:274-275.
[3]王 琰,王慕邹. 姜黄属常用中药的研究进展[J]. 中国药学杂志,2001,36(2):80-84.
[4]许丽珍,吴永辉,岳保超,等. 基于层次分析法对姜黄属植物的观赏应用综合评价[J]. 仲恺农业工程学院学报,2015,28(1):8-16.
[5]田雪慧. 姜黄属花卉植物研究进展[J]. 杨凌职业技术学院学报,2016,15(3):1-2.
[6]梁 恒,邓家彬,童珊珊,等. 我國姜黄属植物的系统关系研究进展[J]. 分子植物育种,2019,17(11):3695-3702.
[7]野 津,张文森,王知斌,等. 姜黄属多糖的提取分离和含量测定研究进展[J]. 化学工程师,2019,33(12):45-47.
[8]廖 婉,张 倩,孙良堃,等. 萜烯缀合姜黄素类微量成分与姜黄素类成分在不同姜黄属药材中的定性与定量分析[J]. 中草药,2020,51(4):1076-1081.
[9]杨小芳,陈育才,刘建福,等. 不同产地姜黄属植物中姜黄素类成分及矿质元素分析[J]. 植物学报,2019,54(3):335-342.
[10]陈金凤,熊 亮,刘 菲,等. 蓬莪术姜黄素类化学成分研究[J]. 中草药,2020,51(1):16-20.
[11]王 颖,郭兰萍,黄璐琦,等. 姜黄、莪术、郁金的化学成分与药理作用研究进展[J]. 中国药房,2013,24(35):3338-3342.
[12]Liu F,Chen J F,Qiao M M,et al. Seven pairs of new enantiomeric sesquiterpenoids from Curcuma phaeocaulis[J]. Bioorganic Chemistry,2020,99:103820.
[13]宋莉平,王 宇. 姜黄素药理作用及机制研究进展[J]. 中国医药导报,2020,17(20):29-33.
[14]窦晓鑫,杨玉莹,卜志超,等. 姜黄素逆转结直肠癌化疗耐药的药理作用机制研究进展[J]. 药物评价研究,2020,43(9):1893-1897.
[15]秦宇雯,赵 祺,赵宇烁,等. 温郁金抗新型冠状病毒肺炎(COVID-19)的机制探索[J]. 中草药,2020,51(8):1977-1983.
[16]吴东雪,候 宁,李 晶,等. 基于药性组合的姜黄、郁金、莪术的性效关系研究[J]. 中国中药杂志,2019,44(2):229-234.
[17]拱健婷,王佳宇,李 莉,等. 基于电子鼻气味指纹图谱与XGBoost算法鉴别姜黄属中药[J]. 中国中药杂志,2019,44(24):5375-5381.
[18]张付轩,卢红委. 郁金、姜黄、莪术、片姜黄鉴别[J]. 中国中医药现代远程教育,2017,15(21):147-148.
[19]王三根. 植物生理学[M]. 北京:科学出版社,2013:95.
[20]Wang K,Zhu J J,Yu L Z,et al. Effects of shading on the photosynthetic characteristics and light use efficiency of Phellodendron amurense seedlines[J]. Chinese Journal of Plant Ecology,2009,33(5):1003-1012.
[21]张 婷. 6种常春藤品种的光合生理特性比较[J]. 西北林学院学报,2019,34(6):62-67.
[22]周玉霞,巨天珍,王引弟,等. 3种旱生植物的叶绿素荧光参数日变化研究[J]. 干旱区资源与环境,2019,33(5):164-170.
[23]李永华,张开明,于红芳. 10个秋菊品种的光合特性及净光合速率与部分生理生态因子的相关性分析[J]. 植物资源与环境学报,2012,21(1):70-76.
[24]郭运雪. 3种金鸡菊的光合生理特性及抗旱性研究[D]. 杭州:浙江农林大学,2012.
[25]何文兴,易 津,李洪梅. 根茎禾草乳熟期净光合速率日变化的比较研究[J]. 应用生态学报,2004,15(2):205-209.
[26]彭昭良,宋凤鸣,黄威龙,等. 不同光照强度对9种姜科植物生长的影响[J]. 广东农业科学,2018,45(2):29-35.
猜你喜欢
江苏农业科学(2016年8期)2017-02-15
热带农业科学(2016年11期)2017-01-21
山东农业科学(2016年12期)2017-01-21
山东农业科学(2016年12期)2017-01-21
山东农业科学(2016年11期)2016-12-17
长江蔬菜·学术版(2015年1期)2015-07-10
长江蔬菜·学术版(2015年6期)2015-06-24
