
甘蓝BoSVP mRNA运输鉴定及其对砧木SVP转录表达的影响
2021-09-10 06:25赵彦婷岳智臣雷娟利李必元
核农学报 2021年10期
陶 鹏 赵彦婷 岳智臣 雷娟利 李必元
(浙江省农业科学院蔬菜研究所,浙江 杭州 310021)
植物MADS-box 蛋白家族由一类包含MADS-box结构域的转录调控因子组成[1-2],在植物花器官分化、开花时间调控以及果实成熟中扮演着重要的角色[3-4]。SVP 蛋白包含典型的MICK 结构域,属于MADS-box 蛋白家族中StMADS11 亚家族的一员。此外,SVP 是植物开花负调控因子[5-6]。SVP 可与FLC(FLOWERING LOCUS C)形成复合体抑制开花启动基因的表达,从而实现开花推迟[7]。研究显示SVP 蛋白也可以通过直接结合pri-miR172a 启动子,以抑制miR172 的转录,通过调控miR172 及其靶基因的表达推迟开花[8]。SVP 蛋白还可以通过结合到FT与SOC1 基因的启动子CArG 盒上以抑制FT、TSF和SOC1 的转录表达,从而推迟开花[9-10]。
在模式植物拟南芥中,AGL24 是SVP的旁系同源基因,两者基因序列相似,但生物学功能相反[11]。在拟南芥(Arabidopsis thaliana)中,AtAGL24 是一种剂量依赖型的开花促进基因,在顶端分生组织中调控开花[12]。前期研究发现,菜心(Brassica campestrisL. ssp.chinensisvar.utilisTsen et lee)BrAGL24 的mRNA 可在甘蓝/菜心异源嫁接体中进行长距离运输[13]。在拟南芥和青花菜中也发现AGL24 的mRNA 可长距离移动并促进接穗提前开花[14]。而序列相似的SVP基因是否也具有mRNA 长距离运输的特性尚不清楚。目前常采用构建异源嫁接体并结合转录组测序来批量研究穗砧之间的mRNA 运输。在葡萄嫁接体中,通过转录组测序检测到3 333 个基因的mRNA 可在穗砧之间运输[15]。在烟草/拟南芥异源嫁接体中检测到拟南芥中有138 个基因的mRNA 运输到烟草中[16]。在烟草/番茄异源嫁接中鉴定到1 096 个基因的mRNA 穿过嫁接连接处进行垂直方向的运输[17]。本研究构建了甘蓝/菜心异源嫁接体,并对砧木菜心花序轴进行转录组测序,利用种间差异序列在菜心转录组测序文库中筛选到来自甘蓝(Brassica oleraceavar L.capitataL.)的BoSVPmRNA read,比较砧木菜心中外源BoSVP和内源BrSVP之间的read 数量差异,分析嫁接体的砧木菜心花序轴和菜心实生苗花序轴中SVP的转录表达情况,以期为深入解析甘蓝/菜心嫁接诱导接穗提早开花的分子机制提供基础数据。
1 材料与方法
1.1 材料及取样
甘蓝G27 和49 菜心种子来自浙江省农业科学院蔬菜所。将甘蓝G27 种子播于穴盘中,49 菜心种子晚一周播种。待甘蓝G27 生长至45 d,将其茎尖嫁接到49菜心的花序轴上,构建甘蓝/菜心嫁接体,留6 株49 菜心实生苗继续生长作为对照组。嫁接后30 d,对甘蓝/菜心嫁接体的嫁接处下方1 cm 的菜心花序轴进行取样(标记为T1、T2 和T3),对菜心实生苗相对应部位的花序轴进行取样作为对照组(标记为T4、T5、T6)。RNA提取、转录组测序工作均由北京百迈客公司完成。
1.2 甘蓝BoSVP 和菜心BrSVP 的mRNA 全长序列的获得
参考甘蓝SVP基因(Bo4g149800)序列,从甘蓝实生苗的转录组测序文库中筛选甘蓝SVP基因的read,并拼接获得甘蓝SVP基因的mRNA 全长序列,命名BoSVP。参考白菜Brassica rapassp.pekinensis SVP基因(Bra030228)的序列,从菜心实生苗的花序轴的转录组测序文库中筛选菜心SVP的read,拼接并获得菜心SVP基因(命名为BrSVP)的mRNA 全长序列。
1.3 甘蓝BoSVP 和菜心BrSVP 的杂合SNP 分析和种间差异序列的选择
基于甘蓝转录组测序各样品的read,使用TopHat2[18]与甘蓝参考基因组序列进行比对,使用GATK The Genome Analysis Toolkit[19]软件识别测序样品与参考基因组间的单碱基错配,识别潜在的单核苷酸多态性(single nucleotide polymorphism,SNP)位点,并标注BoSVP中杂合的SNP 位点。基于菜心转录组测序各样品的read,使用TopHat2 与白菜参考基因组序列进行比对,使用GATK 识别测序样品与参考基因组中的单碱基错配,使用简并碱基标注菜心BrSVP的杂合SNP 位点。GATK 识别标准如下:(1)35 bp 范围内连续出现的单碱基错配不超过3 个;(2)经过序列深度标准化的SNP 质量值大于2.0。使用Clustal Omega 将BoSVP和BrSVP进行序列比对,在5′UTR、编码区以及3′UTR 中均匀标注种间差异序列并进行编号,以备后期筛选read。
1.4 种间差异序列的比较评估
为评估种间差异序列的准确性,以每一个种间差异序列及其反向互补序列作为检索序列,使用UltraEdit 软件在砧木菜心T1、T2 和T3 文库中筛选菜心BrSVP的read,并统计数量。由于每个文库中的read 总数不一致,需要对种间差异序列检索到read 数量进行标准化。本研究采用RPKM reads per kilobase per million mapped reads 的计算方法对所获得read 数量进行标准化,使用Excel 表格制图。计算相邻两组read 数量的标准偏差,评估种间差异序列的准确性。
1.5 BoSVP mRNA 运输分析
分别以种间差异序列G7 和G8 及其对应的反向互补序列作为检索序列,使用UltraEdit 软件在甘蓝/菜心嫁接的砧木菜心花序轴转录组测序文库(T1~T3)和对照组菜心实生苗的花序轴转录组测序文库(T4~T6)中筛选来自甘蓝的BoSVP的mRNA 运输的read,并以R 开头分别命名为R1、R2、……。为进一步验证从甘蓝中筛选出来的read 来自于甘蓝的BoSVP序列,将筛选出的reads 与BrSVP和BoSVP序列进行比对。如果read 的序列与BoSVP序列一致,说明甘蓝BoSVP的mRNA 长距离运输到了砧木菜心中。
1.6 SVP 基因表达分析
采用RPKM 作为衡量转录表达水平的指标,计算甘蓝/菜心嫁接体的砧木菜心花序轴和菜心实生苗的花序轴中SVP的转录表达情况。
2 结果与分析
2.1 甘蓝和菜心SVP 基因序列的比较及杂合SNP的鉴定
以甘蓝全基因组测序数据为参考,分析甘蓝BoSVP基因的SNP 变异情况,结果显示BoSVP基因在5′UTR、编码区和3′UTR 均未发现SNP 变异位点,无杂合SNP 位点。参考白菜全基因组数据,基于菜心叶片转录组测序数据,分析49 菜心BrSVP基因的SNP变异情况,结果显示BrSVP在编码区中存在4 个杂合位点(图1)。
将甘蓝BoSVP和菜心BrSVP的mRNA 全长序列进行比对,显示两者在序列上相似性极高。当除去菜心的4 个杂合SNP,两者在编码区中只有4 个核苷酸的差异。而在5′UTR 和3′UTR 之间分别存在6 和16个核苷酸的差异。避开杂合SNP 的干扰,在SVP基因全长mRNA 上选择了9 条种间差异序列,依次命名为C1~C9,分布在5′UTR、编码区以及3′UTR 上,每条种间差异序列总长为30 nt,每条种间差异序列在BoSVP和BrSVP之间至少存在1 个nt 的差异(图1)。

图1 甘蓝BoSVP 和菜心BrSVP 的mRNA 序列比对Fig.1 Sequence alignment of mRNA of BoSVP and BrSVP
2.2 种间差异序列的评估分析
本研究使用菜心的9 条种间差异序列(C1~C9)在菜心转录组测序文库T1~T3 中分析菜心BrSVP基因的转录本read 数量,将其标准化并作图。结果显示使用相同种间差异序列在不同文库中(T1、T2 和T3)检索到read 数量重复性较好。而使用不同的种间差异序列(C1~C9)在相同文库中的检索所获得的read数量之间有较大误差,其中使用位于5′UTR 的C1 和3′UTR 的C9 在T1/T2/T3 文库中检索到的read 数量均明显低于C2~C8 检索所获得的read 数量(图2)。比较相邻两个种间差异序列检索所获得的read 数量标准差发现,使用C7 和C8 进行检索,两者read 数量之间的差异最小。位于3′UTR 的C7 和C8 在BoSVP和BrSVP中分别存在4 个和5 个nt 的差异,序列本身的特异性较高,检索获得的read 数量之间的差异小,重复性最好。
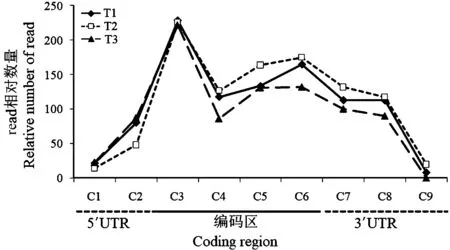
图2 检索所获read 数量的标准化值Fig.2 The standardized value of the number of the retrieved reads
2.3 甘蓝BoSVP 基因mRNA 运输研究
本研究使用特异性和重复性较好的G7 和G8 序列在砧木菜心的转录组测序文库(T1~T3)和菜心实生苗的转录组测序文库(T4~T6)中筛选来自甘蓝的BoSVPmRNA read。在菜心实生苗的花序轴转录组测序文库(对照组)中均未找到甘蓝BoSVPread,而在嫁接苗菜心的转录组测序文库中找到了潜在的异源read(表1)。

表1 甘蓝BoSVP mRNA 运输的read 数量Table 1 The number of transported reads of BoSVP mRNA
为验证砧木菜心中识别的异源read 是否来自接穗甘蓝的BoSVP,从T1~T3 文库中提取这些异源read序列,实际获得的异源read 共有11 个,并命名为R1~R11(图3)。来自T1 文库中的R3 中同时含有完整的G7 和G8 种间差异序列,使用G7 和G8 均能检索到,R3 在G7 和G8 中各计入1 次。对上述提取的11 条运输read 进行序列比对,结果显示R1~R11 均属于甘蓝的BoSVP基因。R1 3′端有一段34 nt 的序列不属于BoSVP基因(图3),将其分别在甘蓝基因组和白菜基因组进行比对,显示R1 的这段34 nt 的序列仍属于甘蓝基因组。

图3 砧木菜心中的异源read(R1~R11)与甘蓝BoSVP和菜心BrSVP 的序列比对Fig.3 The heterologous reads (R1-R11) from rootstocks of the grafted seedlings were aligned with BoSVP and BrSVP
2.4 甘蓝BoSVP 基因mRNA 运输对砧木菜心中SVP 转录表达量的影响
砧木菜心花序轴中SVP基因的转录表达包括内源的BrSVPmRNA 和外源的BoSVPmRNA。为比较内源BrSVPmRNA 和外源BoSVPmRNA 之间数量,本研究使用C7/G7 和C8/G8 两组种间差异序列在砧木菜心转录组测序文库T1/T2/T3 中进行检索。结果显示在T1、T2 和T3 中外源运输的BoSVPmRNA read 数量明显少于内源表达的BrSVPmRNA read 数量(图4)。对菜心实生苗的花序轴和甘蓝/菜心嫁接体的菜心花序轴中的SVP基因的转录表达情况进行分析,结果显示SVP基因在菜心实生苗花序轴(对照组)和嫁接苗的菜心花序轴中的转录表达量差异小于2 倍,P值等于0.81,两组数据差异不显著(图5)。
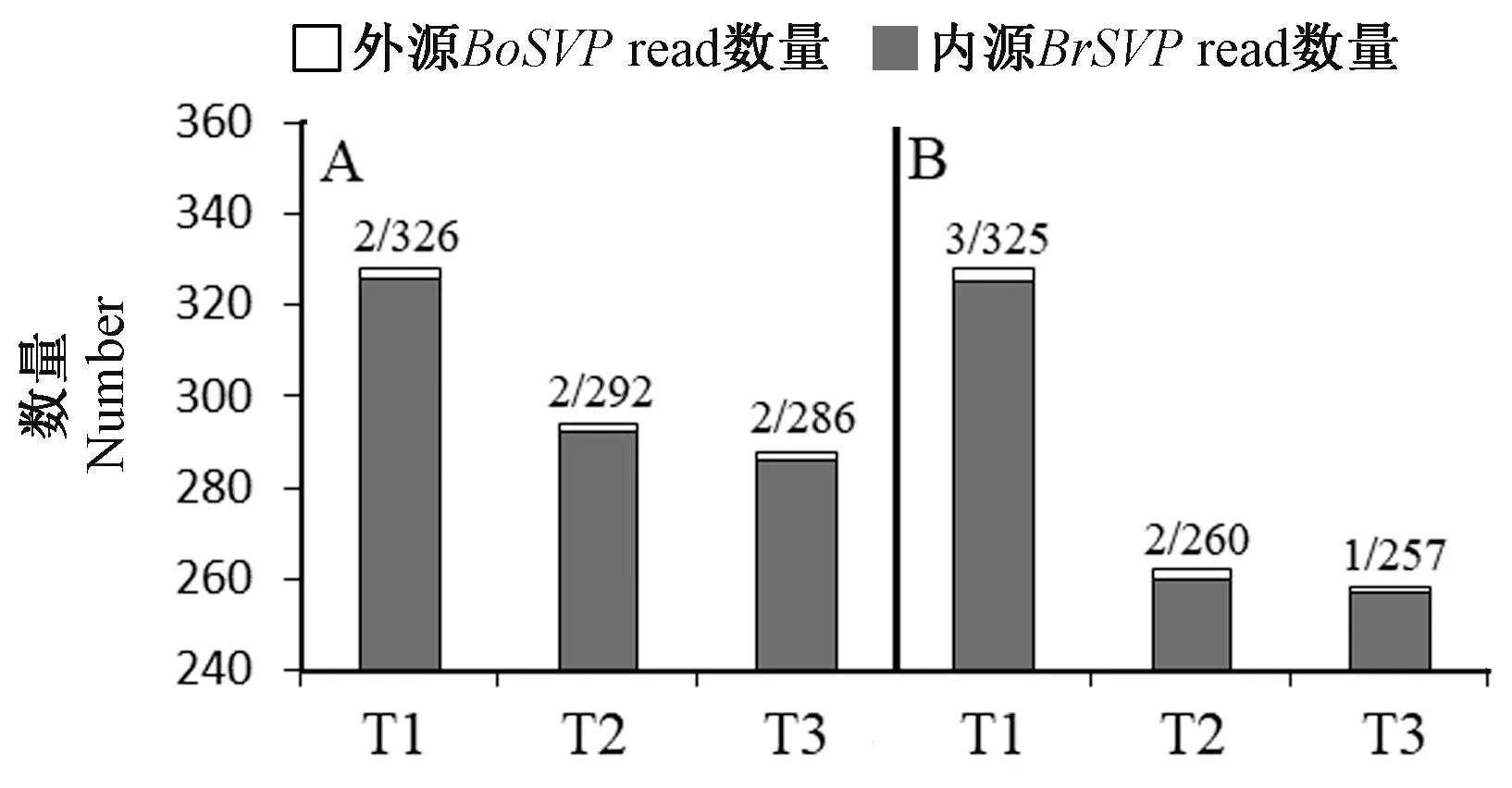
图4 不同文库中内源BrSVP 和外源BoSVP read 数量Fig.4 The number of endogenous BrSVP reads and exogenous BoSVP reads in different library
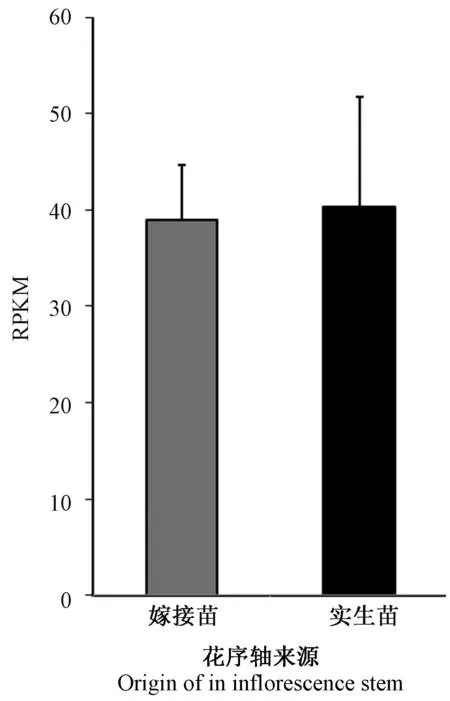
图5 SVP 基因在菜心花序轴中的转录表达量(P 值=0.81)Fig.5 The transcriptional expression of SVP gene in inflorescence stems of Caixin (P value=0.81)
3 讨论
利用甘蓝和菜心SVP基因之间的种间差异序列,在甘蓝/菜心异源嫁接体的砧木菜心花序轴的转录组测序文库中可以准确筛选到甘蓝BoSVPmRNA 长距离运输的read。选择的种间差异序列中不能出现杂合的SNP。由于自身的杂合SNP 可能会导致异源BoSVP的错误识别。所以选择种间差异序列进行菜心BrSVP内源表达评估时候,应尽量避免使用位于mRNA 两端的种间差异序列。在分析菜心砧木转录组测序文库中鉴定异源甘蓝的BoSVPmRNA 的reads时,种间差异序列中的差异核苷酸越多,其特异性越高,所获得的异源甘蓝BoSVPmRNA 可信度越高。使用不同位置的种间差异序列评估菜心内源BrSVPread值时,显示位于BrSVP5′UTR 和3′UTR 两端的种间差异序列获得菜心内源BrSVPread 值与其他差异序列所获得值的偏差较大,可能与mRNA 两端所获得测序机会较少有关。
本研究利用G7 和G8 在砧木菜心的转录组测序文库中鉴定到来自甘蓝BoSVP的11 条mRNA read,在每个嫁接体的砧木菜心的转录组测序文库中均找到了1~3 个甘蓝BoSVPread。mRNA 的长距离运输是一个极其复杂的过程,当前尚未形成统一的观点[20-21]。前期研究认为mRNA 运输可能与mRNA 的序列和基序有关[22]。SVP与AGL24 在序列上具有相似性,同时两者均具有mRNA 运输的特性[13],暗示某些保守的序列决定了SVP基因的mRNA 的运输。比如,拟南芥GAI基因mRNA 在3′UTR 区和编码区的特定基序和序列介导了其mRNA 在嫁接体中的运输[23],马铃薯(Solanum tuberosum)StBEL5 基因的mRNA 可在韧皮部移动,其3′UTR 决定了mRNA 的稳定性及其运输到根部的能力[24-25]。但一些研究推测伴胞中mRNA 的丰度是mRNA 长距离运输的关键因素[26-27]。后期研究需要明确BoSVPmRNA 运输的分子机制。BoSVP的mRNA 从接穗甘蓝运输到砧木菜心中,通过比较砧木菜心中异源BoSVPmRNA 的reads 和内源BrSVPmRNA 的read 数量,显示运输到砧木菜心中的BoSVPmRNA 的read 占内源BrSVPread 的不足1%(图4)。BoSVP的mRNA 运输并不能明显改变砧木菜心中SVP的表达量。SVP基因在甘蓝/菜心嫁接体的菜心花序轴和菜心实生苗花序轴中的转录表达量没有差异,说明BoSVPmRNA 的运输几乎不影响砧木菜心中SVP的表达水平,推测BoSVPmRNA 的运输不会影响菜心中SVP在花序轴中的功能。砧木菜心中异源BoSVPmRNA read 全部来自于接穗甘蓝,理论上将会导致茎尖中甘蓝BoSVP的mRNA 含量减少。甘蓝BoSVP是抑制开花基因,接穗甘蓝中SVPmRNA 的持续减少,可能促进甘蓝提早开花。前期利用甘蓝/菜心的异源嫁接促进了接穗甘蓝的提前开花[28-29]。在甘蓝/菜心嫁接体中,发现砧木菜心的开花促进基因AGL24 的mRNA 从砧木运输到接穗甘蓝中[13]。甘蓝/菜心嫁接促进接穗甘蓝提早开花可能是大量开花促进基因mRNA 向上运输和开花抑制基因mRNA 向下运输综合导致的结果。一些研究认为植物中一些基因的mRNA 可作为信号在穗砧中进行长距离运输并参与生长发育调控[30-31],SVP和AGL24 均为开花调控转录因子基因,SVP和AGL24 运输的mRNA 有可能在甘蓝/菜心嫁接体中发挥调控开花的作用。
4 结论
本研究结果表明,甘蓝BoSVPmRNA 可从接穗甘蓝运输到砧木菜心中,但不影响砧木菜心中SVP基因的转录表达量,同时证明其旁系同源基因SVPmRNA也可长距离运输。由于未提出与BoSVPmRNA 运输直接相关的序列和结构,后期应将mRNA 运输属性与基因序列和结构进行关联分析。本研究为植物基因mRNA 运输机制研究提供了基础数据。
猜你喜欢
草业科学(2022年6期)2022-06-29
中国农业科学(2022年10期)2022-06-28
广东蚕业(2021年12期)2022-01-18
中学生理科应试(2021年10期)2021-12-07
猪业科学(2021年3期)2021-05-21
山西农业科学(2021年1期)2021-01-18
中华诗词(2019年1期)2019-11-14
上海故事(2017年10期)2017-10-19
饮食与健康·下旬刊(2016年9期)2016-05-14
西湖(2016年9期)2016-05-14
