
水稻心白突变体xb1的淀粉理化特性分析
2021-09-10 06:25姜秀英解文孝
核农学报 2021年10期
吕 军 姜秀英 刘 军 解文孝 韩 勇 沈 枫
(辽宁省水稻研究所,辽宁 沈阳 110101)
随着生活水平的日益提高,人们对稻米品质尤其是食味品质有了更高的要求。淀粉是稻米胚乳的主要成分,其组成、结构和理化特性决定了稻米的外观品质和食味品质[1-3]。因此,深入研究水稻淀粉特性,对改良水稻品质具有十分重要的意义。
垩白是由于籽粒灌浆不充分,胚乳中的淀粉和蛋白颗粒排列疏松,相互间存在的空隙影响了光的透射而形成的白色不透明部分。根据其在胚乳上的不同位置可分为腹白、背白和心白等多种类型[4-6]。胚乳淀粉排列不紧密,导致米粒在加工过程中易断裂,整精米率降低,蒸煮后米粒断裂,影响食味品质[7-8]。
长期以来,已利用自然突变和人工诱变获得了大量胚乳淀粉突变体材料,如垩白突变体、高直链淀粉突变体、糯性突变体、粉质突变体、糖质突变体、暗色突变体、皱缩突变体等[9-12],其中已报道的垩白突变体有gpa1、osbt1、ospk2、flo4、flo5 等。并对这些突变体开展了淀粉合成相关研究,发现这些突变体的淀粉特性均发生了一定的变化,如直链淀粉含量、淀粉粘滞特性、热力学特性、支链淀粉链长分布等。进一步通过基因克隆及功能研究,逐步明确了其在淀粉合成及调控中的作用。
Wang 等[13]发现突变体gpa1 呈现白心胚乳,胚乳中复合淀粉颗粒呈圆形且包裹松散,该突变基因OsRab5a位于第12染色体,编码一个GTP酶(GTPase),参与胚乳细胞液泡中蛋白质的转运。细胞壁转化酶基因GIF1,位于第4 染色体,参与籽粒灌浆早期的碳源分配,该基因突变后产生心白,粒重、直链淀粉和支链淀粉含量降低。Li 等[14]发现突变体osbt1呈现心白表型,总淀粉和直链淀粉含量降低,支链淀粉中短链增加,中长链减少,复合淀粉颗粒发育不正常,该突变基因OsBT1 编码ADPG 转运蛋白,在水稻种子发育过程中参与调控淀粉合成和复合淀粉颗粒的形成。Cai 等[15]发现突变体ospk2 呈现心白表型,总淀粉和直链淀粉含量显著下降,蛋白和脂肪含量增加,支链淀粉中中短链比例降低,中长链增多,该突变基因OsPK2 编码一种质体丙酮酸激酶(pyruvate kinase PK),在淀粉合成、复合颗粒形成和籽粒灌浆过程中起重要作用。Flo4 是一个T-DNA 插入突变体,胚乳呈现心白表型,粒重下降,蛋白质含量和脂肪含量增加,该突变基因OsPPDKB编码丙酮酸磷酸双激酶,在水稻胚乳淀粉代谢和结构中起重要作用[16]。flo5 突变体籽粒呈现心白表型,粒重减小,支链淀粉的长链[聚合度(degree of polymerization,DP)≥30]显著减少,快速黏度分析仪(rapid viscosity analyzer,RVA)谱发生显著变化,突变基因SSIIIa编码淀粉合成酶,主要参与支链淀粉B2-B4 链的延伸[17-18]。SSIIa和SSIIIa双突变植株中,胚乳支链淀粉的精细结构发生变化,短链和长链减少,中长链含量增加,双突植株表现出明显的垩白表型[19]。近几年,一系列水稻flo突变体被鉴定,包括flo12、flo14、flo15、flo16 等[20-24],这些突变体的突变基因有些还未被克隆,有些虽已被克隆,但调控的分子机理尚不清晰。水稻胚乳淀粉的合成与调控是非常复杂又精细的过程,因此还需要挖掘更多胚乳淀粉突变体,进一步解析淀粉的生物合成途径与调控代谢网络,为稻米品质改良提供坚实的理论基础。
本研究以经甲基磺酸乙酯(ethyl methane sulfonate,EMS)诱导粳稻品种辽星1 号获得的心白突变体xb1 为材料,对其表型特征及淀粉理化特性进行了分析,旨在了解心白突变的形成机制,为进一步的突变基因克隆和功能分析奠定基础。
1 材料与方法
1.1 突变体材料
辽星1 号经EMS 诱变处理,在其后代对胚乳垩白性状进行筛选,从中发现一个能够稳定遗传的、表型明显的心白突变体,命名为xb1。野生型辽星1 号和突变体xb1 种植于辽宁省水稻研究所试验地,成熟期收获籽粒,自然晾干,并在室温下保存。
1.2 突变体表型特征
利用Laser Jet M1136 扫描仪(惠普中国)对野生型和突变体xb1 的籽粒、糙米和糙米横截面进行拍照。利用游标卡尺测量10 粒种子的粒长、粒宽和粒厚,求平均值,重复3 次。随机选取1 000 粒种子称重,求平均值,重复3 次。
1.3 淀粉粒结构的扫描电镜观察
分别选取野生型和突变体xb1 的完整精米样品,用刀片将米粒横向切开,使横断面向上,利用真空镀膜仪对横断面镀金后,在JSM-7500F 扫描电镜(日本电子)上观察和拍照。
1.4 淀粉粒的粒径分布
淀粉提取:将精米用3 倍体积的亚硫酸钠水溶液浸泡,精米充分吸收膨胀后,转移至匀浆机中匀浆粉碎,随后将匀浆液转移至200 目筛网中过滤,并用亚硫酸钠水溶液充分清洗滤渣,收集滤液,静置过夜,倒掉上清液,用新的亚硫酸钠溶液清洗,重复5~8 次,待淀粉呈白色,上清液透明时,彻底倒掉上清液,将淀粉自然晾干,转移至恒温箱中水分平衡1 周,研钵研磨分散,过200 目样品筛。
取100 mg 淀粉于干净EP 管中,加入1 mL 75%酒精,涡旋混匀后超声混匀,利用Matersizer 3000 激光粒度分析仪(Malvern Instruments Ltd,Worcestershire,英国)进行粒度分布测定,重复测定3 次。
1.5 总淀粉、直链淀粉和蛋白质含量测定
总淀粉含量测定:称取100 mg 淀粉于15 mL 试管中,加入4 mL 80%乙醇,70℃放置2 h,期间漩涡混匀。12 000 r·min-1离心10 min,加4 mL 80%乙醇,重复清洗3 次。小心倒出上清,加入0.2 mL 80%乙醇,涡旋混匀,再加入3 mL 耐热α-淀粉酶,沸水浴孵育6 min,期间间断涡旋混匀。将试管放置于50℃的水浴锅中,然后加入0.1 mL 淀粉葡糖苷酶,涡旋混匀,50℃孵育30 min。用蒸馏水调节体积。取0.1 mL 至新的试管中,加入3 mL GOPOG 试剂,涡旋混匀,于50℃孵育20 min,用1510-04201 多功能酶标仪(美国赛默飞公司)于510 nm 波长下测定吸光度值。
直链淀粉含量测定:称取10 mg 淀粉于EP 管中,加入100 μL 乙醇,900 μL NaOH 溶液,涡旋混匀。沸水煮10 min,待冷却后定容至10 mL。取0.5 mL 上清于离心管中,加入0.1 mL 乙酸,0.2 mL 碘化钾溶液,定容至10 mL,室温放置10 min 显色,用1510-04201 型多功能酶标仪于620 nm 处测定吸光度值。
蛋白质含量测定: 采用凯氏定氮法,利用KJELTEC2300 凯氏定氮仪(丹麦FOSS 公司)测定野生型和突变体的蛋白质含量,重复3 次,取平均值。
1.6 支链淀粉链长分布
取已纯化淀粉5 mg,重悬于5 mL 双蒸水中,沸水浴60 min,间断涡旋混匀。取2.5 mL 的糊化样品,加入125 μL 醋酸钠,5 μL 三氮化钠和5 μL 异淀粉酶,38℃放置24 h。加入395 μL 硼氢化钠,室温放置24 h。取600 μL 于离心管中,室温氮吹干燥。溶于600 μL 流动相中,12 000 r·min-1离心5 min,取上清,用ICS5000 离子色谱系统进行检测,采用电化学检测器及DionexTMCarboPacTMPA200(3.0 mm×250 mm)离子色谱柱。流动相A(100%水溶液),流动相B(100 mmol·L-1NaOH、1 mol·L-1NaAC),流动相C(100 mmol·L-1NaOH),流速为0.4 mL·min-1,柱温为30℃。每个样品重复测定3 次,取平均值。
1.7 淀粉的粘滞特性
采用3D 型RVA 快速黏度分析仪(澳大利亚Newport Scientific 公司)测定。仪器自动读出的一级参数有:热浆黏度(hot paste viscosity,HPV)、峰值黏度(peak paste viscosity,PKV)、冷胶黏度(cool paste viscosity,CPV)、糊化温度(pasting temperature,PaT),二级参数有:崩解值(breakdown viscosity,BDV =PKVHPV)、消减值(setback viscosity,SBV =CPV-PKV)、回复值(consistency viscosity,CSV =CPV-HPV)。每个样品重复测定3 次,取平均值。
1.8 淀粉的热力学特性
采用200-F3 差示扫描量热仪(differential scanning calorimetry,DSC,德国Netzsch 公司)测量淀粉的热力学特性。称取5 mg 米粉于铝盘中,加入10 μL 超纯水,密封2 h。升温速率为10℃·min-1,温度范围为20~120℃。测定后根据测定曲线的吸热峰边界和面积,分析得到糊化起始温度(onset temperature,To)、峰值温度(peak temperature,Tp)、终止温度(conclusion temperature,Tc)热焓值(enthalpy of gelatinization,△H)和糊化距离(gelatinization range,Tr =Tc-To)。每个样品重复测定3 次,取平均值。
2 结果与分析
2.1 突变体的表型特征
突变体xb1 是粳稻品种辽星1 号经EMS 诱变后筛选得到的能稳定遗传的胚乳垩白突变体,其种子表型与野生型(WT)相似(图1-A),而突变体xb1 的糙米较野生型相比,中心呈现白色粉质不透明,边缘一圈呈现透明,即心白表型(图1-B、C)。在籽粒表型性状上,突变体xb1 种子的粒长与野生型无显著差异(图1-D),但粒厚显著低于野生型(图2-F),粒宽和千粒重极显著降低(图1-E、G)。
2.2 突变体的淀粉粒结构和淀粉粒径分布
利用扫描电镜对野生型和突变体xb1 糙米的横切面进行观察,野生型胚乳中的淀粉颗粒大小均一,排列较紧密,相互嵌合挤在一起呈现规则的多面体结构(图2-A、B、C),而突变体xb1 糙米的不透明部位胚乳中的淀粉颗粒大小不一,排列比较疏松,颗粒与颗粒之间存在较大的间隙,形成卵圆形或正圆形(图2-D、E、F),说明突变体xb1 的淀粉合成过程中出现了异常,导致淀粉结构的改变。
利用激光粒度分析仪对淀粉粒的粒径值和粒径分布进行测定,结果显示(图3),突变体xb1 与野生型的淀粉粒大小分布相似,均呈单峰分布,但其分布曲线的峰值明显高于野生型,且淀粉颗粒的大小分布范围也相对集中。突变体xb1 的平均粒径(11.05 μm)略小于野生型的平均粒径(11.57 μm),但淀粉粒粒径值大于13 左右的淀粉粒数量明显低于野生型。说明淀粉颗粒的粒径分布受到心白突变的影响。
2.3 突变体的种子成分分析
对突变体和野生型种子成分分析结果显示(图4),突变体的蛋白质含量极显著升高,野生型种子的蛋白质含量为8.8%,突变体升高至11.2%。突变体的总淀粉含量极显著减少,由81.5%减少到78.2%,而直链淀粉含量无显著变化,说明该突变并未影响到直链淀粉的合成,而与蛋白质和淀粉中支链淀粉的积累存在密切的联系。
2.4 突变体的支链淀粉链长分布
为进一步分析突变体中支链淀粉结构,对突变体xb1 和野生型的支链淀粉的链长分布进行测定,结果表明,突变体种子的支链淀粉链长分布发生了明显变化(图5),与野生型相比,突变体xb1 的聚合度(DP)在6~9 之间的短链及25~35 之间的中长链比例有所增加,而DP 值在10~24 之间的中短链及36~50 之间的长链比例有所减少,说明突变体xb1 中支链淀粉的精细结构发生了改变。
2.5 突变体淀粉的热力学特性
由表1 和图6 可知,突变体xb1 的峰值温度(Tp)与野生型相近,起始温度(To)和糊化距离(Tr)均略高于野生型,但差异不显著,而终止温度(Tc)显著高于野生型,热焓值(△H)极显著高于野生型。表明该突变不会对糊化温度产生太大影响,但会造成热能值显著提高。
2.6 突变体淀粉的粘滞特性
利用快速黏度分析仪RVA 对突变体与野生型的黏度特性进行了测定,结果显示,两者的淀粉黏度存在明显差异(表2 和图7)。从RVA 谱特征值看,突变体xb1除了糊化温度与野生型相近外,其他参数均发生了极显著的变化,突变体xb1 的热浆黏度(HPV)、峰值黏度(PKV)、冷胶黏度(CPV)和消减值(SBV)分别较野生型升高了55.9%、27.0%、33.1%和111.7%。崩解值(BDV)和回复值(CSV)分别较野生型降低了34.6%和5.8%。
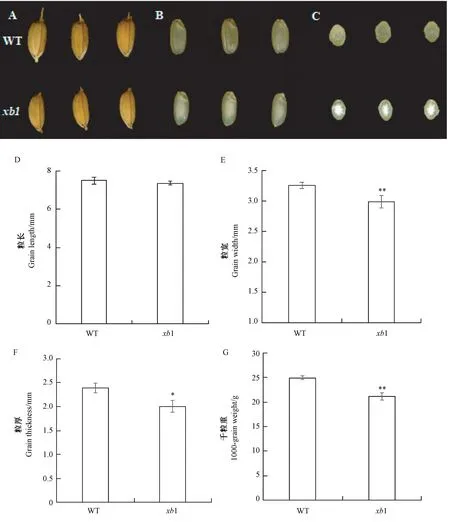
图1 野生型和突变体xb1 的表型分析Fig.1 Phenotypic analysis of wild type and xb1 mutant

表1 野生型与突变体xb1 淀粉的DSC 热力学参数Table 1 DSC thermal parameters of wild type and xb1 mutant

图2 野生型和突变体xb1 的扫描电镜观察Fig.2 Observation of scanning electron micrographs of the wild type and xb1 mutant
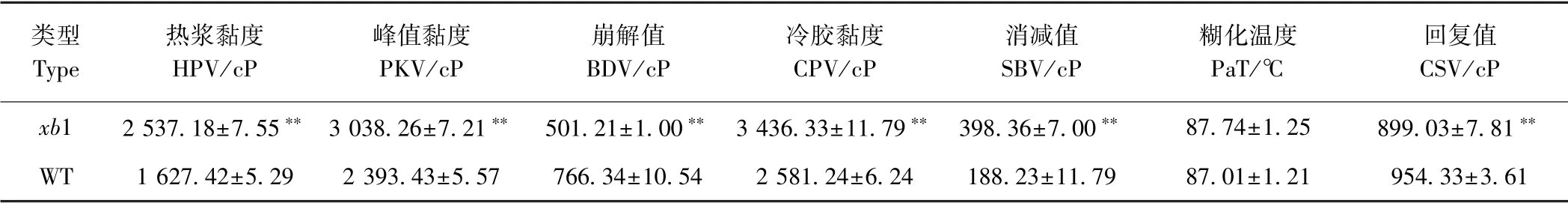
表2 野生型与突变体xb1 的RVA 谱主要参数Table 2 Major parameters of RVA profiles in wild type and xb1 mutant

图3 野生型与突变体xb1 胚乳淀粉粒的粒径值和粒径分布Fig.3 Starch particle size and distribution of the wild type and xb1 mutant
3 讨论
垩白不仅影响稻米的外观品质,而且影响稻米的加工品质、营养品质和蒸煮食味品质,因而直接决定了稻米的市场价格和受欢迎程度[25-26]。而垩白的形成机制较为复杂,与水稻植株内“源库流”的协调、籽粒灌浆动态及胚乳内淀粉的形成和积累密切相关,同时还易受环境条件的影响,因此,垩白性状的研究是当前水稻品质研究的热点和难点[27-31]。本研究筛选到一个稳定遗传的心白突变体xb1,其籽粒的粒长未发生明显改变,但粒宽、粒厚和粒重显著低于野生型。电镜扫描结果显示突变体xb1 的淀粉颗粒大小不一,排列比较疏松,呈现卵圆形或正圆形。激光粒度仪测定结果显示突变体xb1 的淀粉颗粒的粒径分布发生了改变。说明突变体xb1 的淀粉合成过程发生了异常,导致淀粉粒结构和分布发生改变。
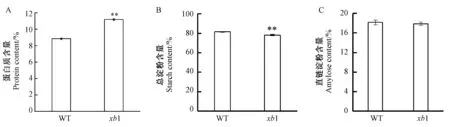
图4 野生型与突变体xb1 的理化性质Fig.4 Physicochemical properties of wild type and xb1 mutant

图5 野生型与突变体xb1 支链淀粉链长分布Fig.5 Chain length distribution profiles of amylopectin in wild type and xb1 mutant
淀粉是胚乳的主要成分,由直链淀粉和支链淀粉两部分组成。本研究结果表明,突变体xb1 在直链淀粉含量没有发生明显变化的前提下,总淀粉含量极显著降低,说明淀粉中支链淀粉含量极显著减少。同时支链淀粉的链长分布也发生了显著变化,相较于野生型,突变体xb1 的聚合度(DP)在6~9 之间的短链及25~35 之间的中长链比例增加,而DP 值在10~24 之间的中短链及36~50 之间的长链比例减少。前人研究表明,支链淀粉的结构被认为是决定淀粉颗粒结构及其功能特性的关键,也是造成直链淀粉含量相近的水稻品种间蒸煮食味品质差异的主要原因[32-33]。支链淀粉的生物合成较为复杂,主要由可溶性淀粉合成酶(SSS),淀粉分支酶(SBE)和淀粉脱支酶(DBE)共同催化合成。Satoh 等[34]研究发现,突变体sbe1 的种子具有正常的表型,但中链(DP12~21)和长链(DP>37)减少,而短链(DP<10)和中长链(DP24~34)有所增加。本研究中突变体xb1 的支链淀粉链长的变化特点,与突变体sbe1 的变异特点一致,但突变体xb1 是否含有sbe1 的等位基因突变型,尚需要进一步验证。

图6 突变体xb1 与野生型的淀粉热力学曲线Fig.6 DSC thermograms of starch in wild type and xb1 mutant
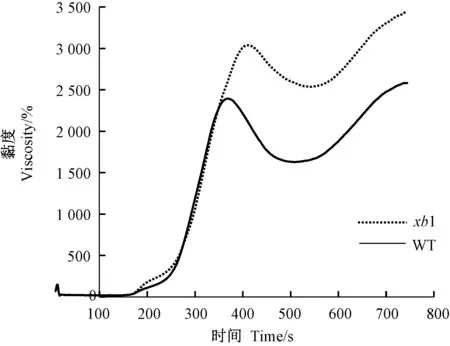
图7 野生型与突变体xb1 的RVA 谱Fig.7 RVA profiles of wild type and xb1 mutan
淀粉结构的改变会导致相应的理化指标的改变。目前关于稻米淀粉结构与淀粉理化特性的关系,不同研究者的研究结论不尽相同。Vandeputte 等[35]认为DP6~9 的短链相对数量与稻米淀粉的糊化起始温度(To)、最高温度(Tp)、终结温度(Tc)及起始成糊温度(pasting temperature,PT)呈负相关,而DP12~22 链长呈现相反结果。贺晓鹏等[36]研究指出,支链淀粉不同链长范围的支链数量比例主要与淀粉的糊化温度相关,而与淀粉的胶稠度和RVA 特征值关系不密切。而突变体xb1 淀粉的糊化起始温度(To)、峰值温度(Tp)、终止温度(Tc)和糊化距离(Tr)均未发生明显改变,这与RVA 谱参数中的糊化温度较野生型没有明显差异的结论一致。这一结果可能是直链淀粉含量未发生改变导致的,即突变体xb1 淀粉的糊化特性与直链淀粉含量关系较为密切。与此同时,突变体xb1 的RVA 谱特征值均发生了显著改变,DP6~9、DP25~35的增加和DP10~24、DP36~50 的减少,使热浆黏度(HPV)、峰值黏度(PKV)、冷胶黏度(CPV)和消减值(SBV)极显著提高,崩解值(BDV)和回复值(CSV)极显著降低。
4 结论
水稻心白突变体xb1 的直链淀粉含量相对野生型未有明显改变,其糊化温度也没有明显差异,进一步证实淀粉的糊化特性与直链淀粉含量密切相关。突变体xb1 的支链淀粉分支结构中,DP6~9(短链)、DP25~35(中长链)的增加和DP10~24(中短链)、DP36~50(长链)的减少,使RVA 谱特征值均发生显著改变,说明淀粉的粘滞特性受支链淀粉分支结构的影响。本研究通过分析心白突变体xb1 的淀粉理化性质,有助于更深入了解淀粉合成、垩白发生机理,为进一步克隆该突变基因及稻米品质改良奠定坚实基础。
猜你喜欢
食品工业科技(2022年21期)2022-10-27
包装工程(2022年1期)2022-01-26
三农资讯半月报(2021年2期)2021-03-22
辽宁农业科学(2021年1期)2021-03-17
航空学报(2019年6期)2019-07-18
北京航空航天大学学报(2019年3期)2019-04-08
小学阅读指南·高年级版(2016年9期)2016-10-31
食品工业科技(2014年13期)2014-12-16
现代营销·经营版(2010年11期)2010-05-14
中学生数理化·高二版(2008年2期)2008-10-19
