
桉树无性系组织比量特征差异分析
2021-09-06 02:20卢翠香苏远玉郭东强李昌荣李建凡陈健波
中国农学通报 2021年22期
卢翠香,苏远玉,郭东强,李昌荣,孙 熹,李建凡,陈健波
(1广西林业科学研究院/广西优良用材林资源培育重点实验室,南宁530002;2广西国有钦廉林场,广西钦州535099;3玉林市林业科学研究所,广西玉林537501)
0 引言
木材组织比量是指构成木材的各种细胞所占横截面积的比例,它是一个重要木材解剖特征,也是决定木材物理力学性质的主要因子之一[1],同时关系到木材的机械和化学加工、利用。研究木材纤维构造的数量特征是探寻树木生长的变异规律及材性预测的重要手段[2]。桉树是阔叶散孔材,其组织比量包括导管、轴向薄壁细胞、射线和纤维[3]。迄今,国内外在枫杨(Pterocarya stenoptera)、刺楸(Kalopanax septemlobus)、马 尾 松 (Pinus massoniana)、杨 树 (Populus)、落 叶 松(Larix gmelinii)、三角枫(Acer buergerianum)、铜钱树(Paliurus hemsleyanus)、尾巨桉(Eucalyptus grandis×E.urophylla)等[4-8]树种上已开展一些木材组织比量变异的研究,并取得了较为理想的效果。费本华等[9]研究表明径向上,随着树龄的增大,铜钱树纤维比量递增,导管、射线和轴向薄壁组织递减。马顺兴等[8]研究发现日本落叶松的射线比量、树脂道比量和管胞比量无性系间差异极显著。胡进波等[10]对8个尾巨桉家系木材研究发现,家系间及家系内个体间的组织比量分化非常明显,且各性状在个体水平上分布范围较广,说明开展个体选择的潜力更大。因此,对大部分树种而言,由于其材性在种内存在广泛的遗传变异,为遗传改良奠定了物质基础,并且可以通过改良获得理想效果。
桉树是中国南方重要的工业用材和多功能高效益人工林树种,具有速生、丰产、高效益、用途广等特点[11]。桉树无性系造林是南方桉树造林的主要手段,在国内桉树主产区,无性系人工林的比例占90%以上[12]。现阶段桉树无性系材性遗传改良研究主要集中在生长性状[13]、木材密度[14]、制浆造纸性能[15]、物理力学性能[16]和纤维形态[17]方面,取得了一定进展,但对桉树无性系组织比量及其变异规律的研究较少,而这类研究对无性系选育、木材的合理高效利用具有重要意义。笔者以11个10年生的桉树无性系为材料,研究组织比量在无性系间、树龄间的遗传变异模式,探讨桉树无性系组织比量改良的潜力,以期为确定桉树无性系生长和材质性状的联合遗传改良策略及无性系的早期选择和人工林的定向培育提供理论依据。
1 材料与方法
1.1 试验地与材料
试验材料取自桉树无性系试验林,试验林位于广西容县玉林市林科所(110°09′E,22°39′N)。2004年5月造林,共有11个桉树无性系参试(表1)。试验采用随机完全区组设计,每小区3行,每行4株,4个重复,株行距2.0 m×3.5 m。整地方式为穴状整地,常规抚育管理。造林后连续2年抚育施肥,每年抚育1次,追肥结合除草进行,追肥为桉树专用复合肥,总有效养分为30%,N:P:K=15:8:7,300 g/(株·次)。
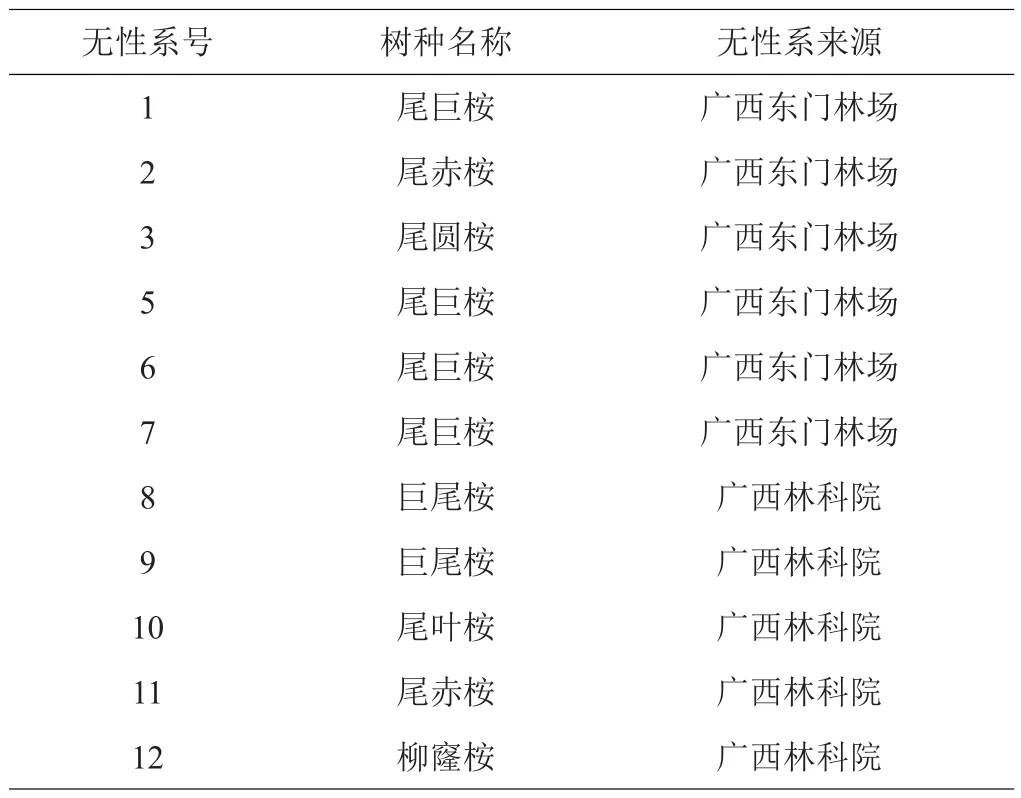
表1 参试桉树无性系概况
1.2 试验方法
2014年调查各无性系的生长性状,每个无性系抽取2株标准样木,伐倒取胸径处5 cm的圆盘供研究。
采用交叉定年法定年,在圆盘上沿髓心向树皮方向分别切取截面为2 cm×2 cm的木样。采用徒手切片切取木样横切面,经番红染色、酒精(30%、50%、70%、100%)脱水、TO溶液透明后,制成临时玻片。采用Nikon 80I数码显微图像电脑分析系统测定,分别计落在网格的交叉点上的导管、轴向薄壁细胞和射线的个数。每个试样测定30次。
1.3 数据处理与分析方法
用Excel软件处理各指标的平均值、标准差和变异系数,采用SPSS 19.0软件[18]对各指标进行方差分析、回归分析,并根据数量遗传学理论知识计算各性状遗传参数[19],无性系重复力计算如式(1),个体重复力计算如式(2),表型变异系数计算如式(3)。

式中,R无表示无性系重复力,R个表示个体重复力,V1表示组间方差,V2为组内方差,N为每一无性系测量个体量,PCV为表型变异系数,系、无性系×区组、环境的方差分量,为性状均值。分别为无性
2 结果与分析
2.1 桉树组织比量无性系间的变异
表2为参试11个桉树无性系木材组织比量数值统计,导管比量均值为11.80%,变异范围10.49%~ 13.45%,5号无性系的值最大,6号无性系的值最小,最大的无性比最小无性系高28.21%。轴向薄壁组织比量均值为0.61%,变异范围0.42%~ 0.89%,1号无性系的值最大,7号无性系的值最小。射线比量均值为8.02%,变异范围6.29%~ 9.79%,5号无性系值最大,4号无性系值最小,最大的无性比最小无性系高52.78%。纤维比量均值为79.53%,变异范围76.37%~ 81.80%,4号无性系的值最大,5号无性系的值最小,最大的无性比最小无性系高6.61%。从变异系数看,除纤维比量的变异系数小于10%外,其余3个指标的变异系数均大于10%。11个无性系中,1号无性系的导管比量变异系数值最大,为31.08%,6号无性系的变异系数值最小,为20.30%;7号无性系的轴向薄壁组织比量变异系数值最大,为108.87%,1号无性系的变异系数值最小,为75.42%。3号无性系的射线比量变异系数值最大,为37.00%,7号无性系的变异系数值最小,为17.90%。由此可见,不同无性系木材组织比量不同,而且同一无性系内也有差异,这是无性系间遗传差异和生长差异所致。
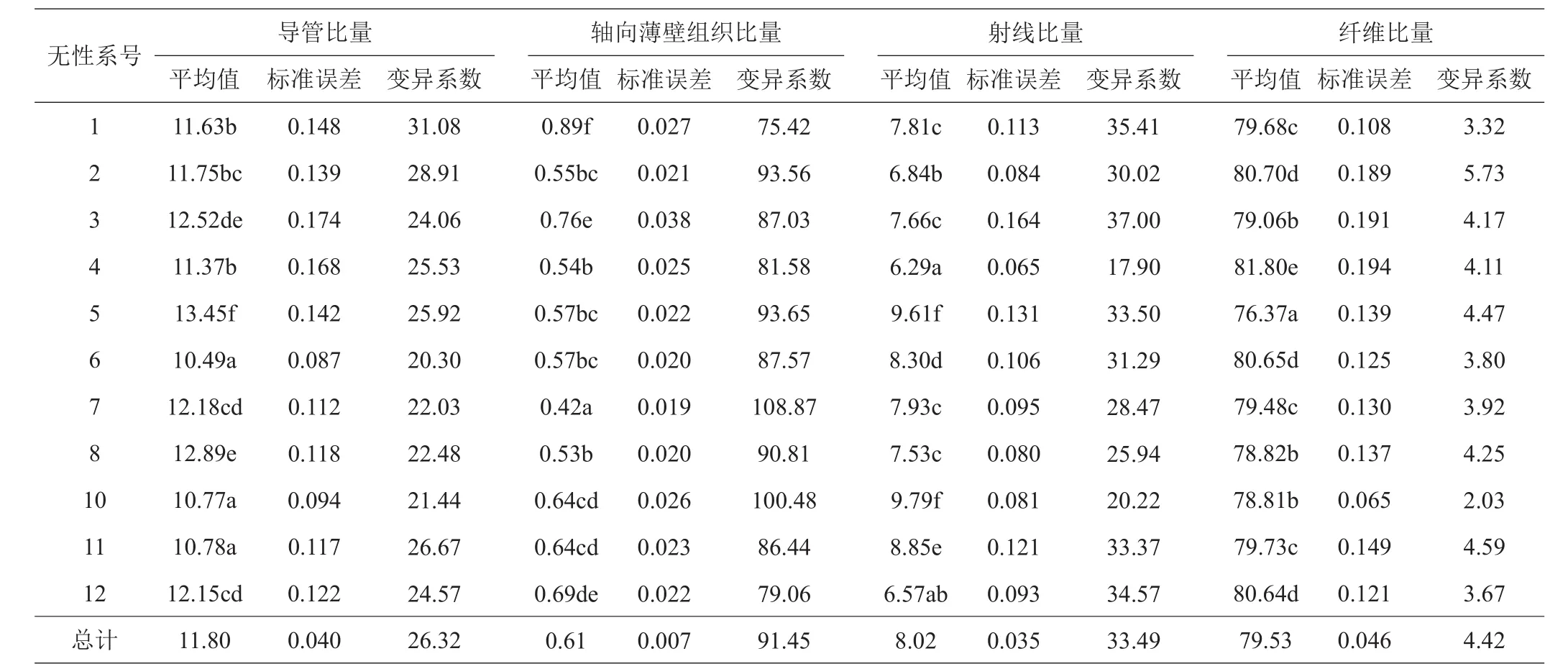
表2 11个桉树无性系木材组织比量及其变异系数 %
方差分析结果(表3)表明,导管比量、轴向薄壁组织比量、射线比量和纤维比量在无性系间存在极显著差异。邓肯多重比较显示,这种差异的具体表现是6号无性系导管比量显著小于1号、2号、3号、5号、7号、8号和11号无性系,而与4号无性系和10号无性系比较无显著差异;7号无性系轴向薄壁组织比量显著小于其他10个无性系;4号无性系射线比量与12号无性系比较无显著差异,而与1号等其他9个无性系比较有显著差异;4号无性系纤维比量显著高于其他无性系,与其他10个无性比较有显著差异。

表3 桉树无性系木材组织比量方差分析
2.2 桉树无性系组织比量的径向变异
桉树无性系组织比量径向变异因构造分子而异,导管比量随树龄的增加先呈递增趋势,第5年开始略有降低,之后基本保持平稳,最小值出现在第1年轮(10.95%),最大值出现在第5年轮(12.34%)(图1a)。导管比量的这种径向变异规律从生物学上分析,可能是木质部母细胞分裂必需的生长因子差异分布的结果。在幼年期,细胞分裂快,导管分子所需的生长因子受到抑制,导管分子的分化较弱,比量较小;到成熟期后,细胞分裂趋于稳定,良好的细胞迅速扩展,并分化为导管分子,故导管比量相对较多[10]。轴向薄壁组织比量随树龄的增加开始迅速下降,第4年后缓慢波动下降,第7年后又迅速上升,波动性较大,表明桉树无性系轴向薄壁组织比量的变异性大,生长速度对桉树无性系轴向薄壁组织比量的影响不大,最小值出现在第7年轮(0.56%),最大值出现在第1年轮(0.68%)(图1b)。射线比量随树龄的增加呈逐渐增加趋势,中间有小幅波动,最小值出现在第1年(7.68%),最大值出现在第10年(8.62%)(图1c)。纤维比量随树龄的增加呈减小趋势,中间有小幅波动,最小值出现在第5年(78.80%),最大值出现在第1年(80.69%)(图1d)。这种结果的产生,可能是由于幼年期形成层原始细胞快速分裂形成的导管细胞较少,形成层后增长作用较弱,促使纤维细胞比量相对多一些;进入成熟期后,形成层原始细胞分裂趋于稳定,管孔的形成层后增长作用加强,促使纤维细胞比量相对要少一些[7]。方差分析结果表明,导管比量、轴向薄壁组织比量、射线比量和纤维比量在树龄间存在极显著差异(表4)。
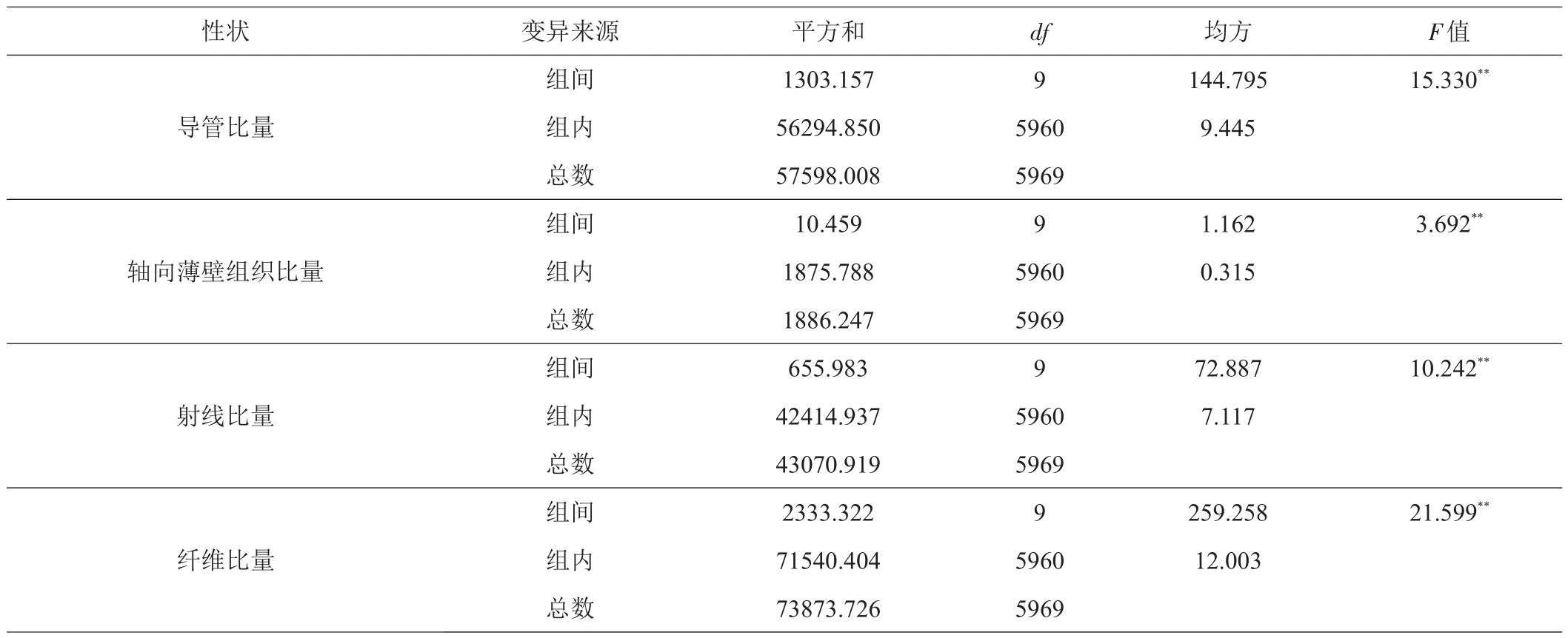
表4 桉树无性系树龄间木材组织比量方差分析
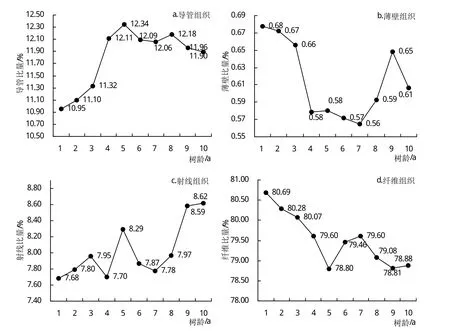
图1 桉树无性系组织比量的径向变异
2.3 桉树无性系的组织比量与年轮的回归
由木材组织比量变异情况可以看出,生理年龄是影响组织比量的一个主要因素,因此采用曲线拟合的方法,对11个桉树的无性系组织比量的平均值(y)与年龄(x)进行回归分析,得到回归方程。从表5可以看出,导管比量采用多项式方程和乘幂式方程拟合效果较好,其相关系数分别达到0.8655和0.6878。轴向薄壁组织比量和射线比量采用多项式方程拟合效果较好,其相关系数分别达到0.7263、0.6130。纤维比量采用对数方程和乘幂式方程拟合效果较好,其相关系数分别达到0.8172和0.8154。

表5 组织比量与年轮的回归关系
2.4 桉树无性系组织比量的遗传参数
由表6可知,桉树木材组织比量的表型变异系数差异较大,变幅4.42%~ 91.45%,说明无性系间存在极大变异。无性系重复力和单株重复力均大于0.90,说明这些性状受到较强的遗传控制。纤维比量无性系重复力值为0.990,而表型变异系数较小,表明这个性状稳定性较好。基于重复力高、表型变异系数小的选择基础,该试验可基于纤维比量、导管比量、射线比量、薄壁组织比量的顺序进行无性系选择,部分性状还应考虑单株效应。

表6 11个桉树无性系材性性状遗传参数分析结果
3 结论与讨论
(1)桉树无性系纤维比量最大,平均79.53%;导管比量次之,平均11.80%;射线比量次之,平均8.02%;轴向薄壁组织比量最小,平均0.61%。这与尾巨桉家系[10]的纤维比量52.86%、轴向薄壁组织比量15.31%、射线比量17.14%、导管比量14.70%不一致,可能是因为尾巨桉家系木材为散孔材,管孔小而疏,而参试的11个桉树无性系均为散孔材,但管孔较尾巨桉大且少;与楸树[20]纤维比量78.06%、导管比量15.75%、射线比量6.19%相一致。轴向薄壁组织的变异性较大,表明生长速度对桉树无性系木材轴向薄壁组织比量的影响不大。方差分析表明,导管比量、轴向薄壁组织比量、射线比量和纤维比量在无性系间差异极显著,说明对桉树无性系的组织比量作遗传改良是可行的。已有研究表明,日本落叶松无性系间木射线比量、树脂道比量和管胞比量差异极显著[8];楸树无性系间纤维比量、导管比量和射线比量具有显著差异[20];杉木不同立地类型间的薄壁组织比量、木射线比量、管胞比量差异不显著[21];巨桉8个无性系株内组织比量在径向上差异均不显著,变异也无一定规律[22];本试验测得的结果与前人测得的阔叶材和针叶材的组织比量变异一致。
(2)11个桉树无性系组织比量在不同树龄间存在极显著差异。其径向变异规律为:导管比量随树龄的增加呈递增趋势,第5年开始略有降低;轴向薄壁组织比量随树龄的增加开始迅速下降,第4年后缓慢波动下降,第7年后迅速上升,然后又降低;射线比量随树龄的增加呈逐渐增加趋势,中间有小幅波动;纤维比量随树龄的增加呈减小趋势,中间有小幅波动。这与尾巨桉家系[10]、日本落叶松[8]、枫杨[4]、刺楸[5]、三角枫[2]、马尾松[6]、马占相思(Acacia mangium)[23]等树种组织比量有着相同径向变异规律。
(3)回归分析结果表明,导管比量采用多项式方程和乘幂式方程拟合效果较好,其相关系数分别达到0.8655和0.6878。轴向薄壁组织比量和射线比量采用多项式方程拟合效果较好,其相关系数分别达到0.7263、0.6130。纤维比量采用对数方程和乘幂式方程拟合效果较好,其相关系数分别达到0.8172和0.8154。
(4)桉树无性系组织比量受到较强的遗传控制,各指标重复力均为0.90以上,开展无性系组织比量选择改良是可行的。
(5)重复力、遗传相关参数随着空间和时间在无性系间变动[12],无性系选择测定需要在空间和时间上同时进行。由于该试验只有一个地点的试验,后续的研究需考虑扩大试验范围,增加试验点,利用近红外光谱、连锁图谱和QTL定位等手段解析桉树生长与材性性状间复杂的遗传背景。
猜你喜欢
中学生数理化·七年级数学人教版(2020年12期)2021-01-18
果树资源学报(2020年3期)2020-05-13
新疆农业科学(2019年11期)2019-12-31
中学生数理化·七年级数学人教版(2019年12期)2019-05-21
儿童故事画报·发现号趣味百科(2019年9期)2019-02-02
环球时报(2019-01-03)2019-01-03
中成药(2017年3期)2017-05-17
天然产物研究与开发(2016年11期)2016-06-15
中国塑料(2016年9期)2016-06-13
广西林业科学(2016年2期)2016-03-20
