
双菌种混合发酵制备富含γ-氨基丁酸酸奶工艺优化
2021-09-02 06:44孙世鑫骆鹏飞俞兰秀莫小叶孙海燕
食品工业科技 2021年16期
孙世鑫,李 科,骆鹏飞,俞兰秀,莫小叶,孙海燕,刘 冬,
(1.深圳职业技术学院深圳市发酵精制检测系统重点实验室,广东深圳 518055;2.绿雪生物工程(深圳)有限公司,广东深圳 518055)
γ-氨基丁酸又名γ-氨酪酸、4-氨基丁酸,是一种在动植物体内广泛存在的小分子量非蛋白质氨基酸。作为哺乳动物重要的抑制性神经递质[1],GABA在日常代谢和神经调节中发挥着巨大作用:如可通过GABAA受体改变神经突触后膜对Cl-的通透性来发挥镇定安神的功效[2],或通过对血管紧张素转换酶的高效抑制来降低血压[3]等。2009年9月,我国正式批准GABA为新资源食品,允许其在除婴幼儿食品之外的饮料等食品中添加使用,且摄入量不超过500 mg/d[4]。
当前,功能性酸奶已经成为乳制品开发的主流。如具备安神助眠功效的酸奶,即可通过直接发酵法[5]制备。在生产中,向乳液内添加在乳基底物中具备较高GABA合成活性的乳酸菌,在发酵的同时利用其胞内的谷氨酸脱羧酶(Glutamate Decarboxylase,GAD,EC4.1.1.15)催化L-谷氨酸或其钠盐,经GABA支路脱羧合成GABA[6],即可直接生产出天然富含GABA的功能性酸奶。研究结果表明,经口服摄入的GABA能够由消化道进入血液循环[7],发挥舒缓焦虑[8]、抗解压力[9]和改善睡眠[10]等保健作用。此外,有研究证实,富含GABA的功能性酸奶在感官指标方面与传统酸奶无显著差异,且乳酸菌活菌数和GABA含量在酸奶保质期内能够保持相对稳定[11]。因此,筛选在乳基底物中具备较高GABA合成活性的乳酸菌并优化其GABA生产工艺具备重要的经济意义。
本研究利用课题组前期从国内市售奶酪、奶疙瘩等发酵食品中筛选得到的在乳基底物中具备高产GABA潜力的乳酸乳球菌乳酸亚种等菌种,以脱脂复原乳液为主要原料,通过单菌种发酵及双菌种发酵的单因素实验,确定了菌种在乳基环境中的最适发酵条件;利用均匀实验构建出双菌混合发酵过程的模型方程,确定了优化后的混合发酵工艺参数,以期为更多富含GABA发酵乳制品的开发应用奠定基础。
1 材料与方法
1.1 材料与仪器
GABA高产潜力菌种乳酸乳球菌乳酸亚种(Lactococcus lactissubsp.lactis,菌种编号4043)、德氏乳杆菌保加利亚亚种(Lactobacillus delbrueckiisubsp.Bulgaricus,菌种编号1060)、副干酪乳杆菌(Lactobacillus paracasei,菌种编号1134)、干酪乳杆菌(Lactobacillus casei,菌种编号1443) 实验室保藏;培养基:MRS肉汤培养基和平板培养基、M 17肉汤培养基和平板培养基 实验室自制;脱脂乳粉市售;四氢呋喃、甲醇、乙腈 色谱纯,天津大茂化学试剂厂;L-Glu-Na(生化试剂纯) 国药集团化学试剂有限公司;其他试剂 均为国产分析纯。
5810R型高速冷冻离心机 德国Eppendoff公司;LC-20AT型高效液相色谱 日本Shimadzu公司;PHS-3C型p H计 上海仪电科学仪器股份有限公司。
1.2 实验方法
1.2.1 培养基配制MRS肉汤培养基和M 17肉汤培养基:蒸馏水混匀后调节pH为7.2±0.1,121℃灭菌15 min。
发酵乳液:用纯水溶解乳粉,配制成10%(w/v)的乳液,按照实验所需质量浓度加入适量L-Glu-Na[12],混匀后分装入100 mL锥形瓶,97℃水浴灭菌15 min后冷却至室温备用。
1.2.2 单因素实验
1.2.2.1 单菌种最适发酵温度的确定 取活化后的4043号菌种子液,按3%(v/v)比例接种于L-Glu-Na质量浓度2 g/L的发酵乳液中,分别于28、30、34、37、40℃下恒温发酵48 h,随后测定发酵终点酸奶的酸度、活菌数、p H及GABA、L-Glu-Na含量。
1.2.2.2 单菌种最适发酵时间的确定 取活化后的4043号菌种子液,按3%(v/v)比例接种于L-Glu-Na质量浓度2 g/L的发酵乳液中,分别于最适发酵温度下恒温发酵12、24、36、48、60 h,随后测定发酵终点酸奶的酸度、活菌数、p H及GABA、L-Glu-Na含量。
1.2.2.3 单菌种最适接种量的确定 取活化后的4043号菌种子液,分别按2%、3%、4%、5%、6%(v/v)比例接种于L-Glu-Na质量浓度2 g/L的发酵乳液中,在最适发酵温度和发酵时间下恒温发酵至终点,随后测定酸奶的酸度、活菌数、p H及GABA、L-Glu-Na含量。
1.2.2.4 单菌种最适L-Glu-Na添加量的确定 取适量L-Glu-Na加入10%(w/v)乳液,分别配制L-Glu-Na质量浓度为0、1、2、3、4、5 g/L的发酵乳液。取活化后的4043号菌种子液按5%(v/v)的比例转接于上述发酵乳液中,最适发酵温度和发酵时间下发酵至终点,随后测定酸奶的酸度、活菌数、pH及GABA、L-Glu-Na含量。
1.2.2.5 双菌种复配方案及最适发酵温度的确定将活化后的1443号菌、1060号菌、1134号菌分别与4043号菌的种子液按1:1比例进行复配,并按6%(v/v)的总比接种于L-Glu-Na质量浓度2 g/L的发酵乳液中,分别在34和37℃下恒温发酵48 h,随后测定发酵终点酸奶的酸度、活菌数、p H及GABA、L-Glu-Na含量。
1.2.2.6 双菌种最适发酵时间的确定 将活化后的4043号菌和1443号菌种子液按1:1比例进行复配,并按6%(v/v)的总比接种于L-Glu-Na质量浓度2 g/L的发酵乳液中,分别于34℃恒温发酵12、24、36、48、60 h,随后测定发酵终点酸奶的酸度、活菌数、p H及GABA、L-Glu-Na含量。
1.2.2.7 双菌种最适总接种量的确定 将活化后的4043号菌和1443号菌的种子液按1:1比例进行复配,并分别按4%、6%、8%、10%、12%的总比接种于L-Glu-Na质量浓度2 g/L的发酵乳液中,在34℃下恒温发酵48 h,随后测定发酵终点酸奶的酸度、活菌数、pH及GABA、L-Glu-Na含量。
1.2.3 均匀实验 根据单因素实验结果,选择发酵时间、发酵温度、L-Glu-Na添加量、球/杆菌配比为影响因素,并设定适宜水平,进行均匀实验。使用DPS 7.05(Data Processing System 7.05)数据处理系统,制作一个U12(12×63)混合水平均匀设计表,在录入因素及水平个数后,控制最大寻优时间为5 min、最大迭代次数为1000次的优化操作,最终得到的混合水平均匀设计方案(中心化偏差CD=0.1296),如表1所示。

表1 混合水平均匀设计方案Table 1 Mixed-level uniform design scheme
1.2.4 GABA和L-Glu-Na的定量检测 使用HPLC法,在QB/T 4587-2013检测方法基础上加以改进。精密吸取适量样品液与等体积的邻苯二甲醛衍生剂混合,室温反应2 min后经0.22μm尼龙针头过滤器转移至液相进样瓶。流动相A为25 mmol/L醋酸钠,调p H至7.2;流动相B为甲醇和乙腈,混合比例为50:50,两相均经0.45μm有机系滤膜抽滤后超声脱气20 min后使用。色谱柱Agilent ZORBAX Eclipse Plus C18(4.6 mm×250 mm,5μm),柱温为40℃,流速为1 mL/min,进样量为20μL,检测波长334 nm,梯度洗脱(梯度洗脱程序如表2所示)。

表2 梯度洗脱程序Table 2 Gradient elution program
配制10、50、100、150、250μg/mL的GABA标准工作液并绘制标准曲线。以峰面积(y)对GABA浓度(x)做线性回归,得线性回归方程y=14179x−59007,线性决定系数R2=0.9998。
配制10、100、500、1000、1500 μg/mL的L-Glu-Na脱脂乳溶液并绘制标准曲线。以峰面积(y)对LGlu-Na浓度(x)做线性回归,得线性回归方程y=5392x+534801,线性决定系数R2=0.9989。
1.2.5 酸度和p H的测定 酸奶酸度参照国标GB 5009.239-2016《食品酸度的测定》,采用乳与乳制品酸度的测定方法。取酸奶样品10 g于盛有20 mL蒸馏水的烧杯中,滴加适量酚酞混匀后,用0.1 mol/L的标准NaOH溶液滴定至淡粉色且30 s不褪色,即为滴定终点。记录消耗的NaOH溶液体积(mL),计算酸度值(°T)。
酸奶pH采用pH计测定。取酸奶样品10 g于烧杯中,按照1:1添加去离子水混匀,用PHS-3C型p H计测定稀释后样品的pH。
1.2.6 乳酸菌活菌数的测定 参照国标GB4789.35-2016《食品微生物学检验乳酸菌检验》,采用平板计数法。取酸奶样品25 g于盛有225 mL灭菌生理盐水的烧杯中混匀,后取10 mL于90 mL灭菌生理盐水试管中混匀,依次操作进行10倍稀释后选取适合的梯度,吸取100μL的样品匀液涂布于MRS平板培养基,在37℃下厌氧培养48 h,选取菌落数为30~300的平板计数,以lg CFU/mL表示活菌数。
1.2.7 感官评定 酸奶由10名食品专业学生作为感官鉴评员分别评分,取平均值。感官评分指标及标准参考食品安全国家标准GB 19302-2010并做适当修改,如表3所示。

表3 酸奶感官评分指标及标准Table 3 Sensory evaluation index and standards of yogurt
1.3 数据处理
每次试验均做3次平行,数据结果以Mean±SD表示。实验数据的差异显著性用SPSS 22.0软件中one-way ANOVA法检验,P<0.05为有显著差异,并用GraphPad Prism 8软件作图。正交试验和二次正交旋转组合实验使用DPS7.05软件进行实验设计和数据分析。
2 结果与分析
2.1 单菌种发酵单因素实验结果
2.1.1 最适发酵温度的确定 温度是影响菌株生长代谢的重要因素。发酵温度对单菌种发酵结果的影响如图1所示。由图1可知,酸奶中GABA产量随发酵温度升高呈现先升后降的趋势。由于L-Glu-Na是乳酸菌GAD转化合成GABA的底物,故LGlu-Na残留量即表现为相反的先降后升。比较可见,GABA产量与活菌数两者变化趋势并不同步,究其原因,应与温度对4043号菌种的生长特性及其GAD的酶学性质的影响不同有关。随着发酵温度升高,酸奶中活菌数显著增长,至30℃达到最高值,表明30℃为4043号菌种的最适生长温度。随着温度继续升高,4043号菌种的生长受到抑制,导致活菌数迅速下降。而有研究显示,乳酸菌GAD的最适反应温度分布在30~50℃区间[5],在40℃左右活力最高[13]。因此在30~40℃范围内,4043号菌种的GAD活力应随温度升高而持续增强。当发酵温度在30~34℃范围,活菌数开始减少,GAD活力的增强抵消了GAD总量减少的消极影响,使L-Glu-Na的转化效率在总体上提升,GABA产量继续增加。但随着温度升高,活菌数进一步减少,使GAD总量显著降低,即便活力进一步增强也无法提升转化速率,导致GABA产量开始下降。4043号菌种GABA产量随温度升高先升后降的现象与干酪乳杆菌NFRI7415的GABA产量在37~43℃发酵温度区间内的变化[14]类似。因此,当发酵温度为34℃时,GABA产量即为最高值,约0.52±0.02 g/L。此外,研究显示,L-Glu-Na在清水中的呈鲜阈值为0.30 g/L[15],在酸奶中的呈鲜阈值约为0.13 g/L[16]。若L-Glu-Na残留量高于0.13 g/L,则酸奶会呈现出较为明显的咸、鲜等味道,不利于保持酸奶的良好风味,因此应尽量减少发酵终点L-Glu-Na的残留。L-Glu-Na残留量在34℃时为0.50±0.03 g/L,虽高于0.13 g/L,但已是最低值,且较其他发酵温度组均具备显著性差异(P<0.05)。故确定34℃为最适发酵温度。

图1 发酵温度对单菌种发酵结果的影响Fig.1 Influenceof fermentation temperature on result offermentation with singlestrain
2.1.2 最适发酵时间的确定 对发酵时间的合理控制有助于提高效率,降低成本。由图2可知,发酵时间为12~24 h时,GABA产量增长缓慢;进入24~48 h阶段,GABA产量随时间延长增加显著(P<0.05);当发酵时间进一步延长至48~60 h阶段,GABA产量趋于稳定。当发酵时间为48 h时,GABA产量为0.68±0.02 g/L,已达到最大产量的95%左右。此外,活菌数表现为在12~24 h阶段显著(P<0.05)增长,在24~60 h阶段持续下降(P<0.05)。结合菌株生长曲线和GABA产量曲线,可以发现GABA的生产过程可以大致分为两个阶段:12~24 h为延滞期,24~60 h为积累期。推测可能是因为:在延滞期,发酵乳液中营养丰富,菌株快速生长并大量产酸,导致酸度显著增加(P<0.05),pH降低,形成酸性环境。在此阶段,GAD活性活性增加较慢[5]。随后酸性环境激活并诱导GAD表达[17],L-Glu-Na脱羧反应得以顺利进行,GABA开始生成并进入积累期。随着发酵时间延长,发酵乳液内营养物质逐渐消耗殆尽,菌株个体间竞争加剧[18],活菌数减少,导致反应速度和产物积累速度逐渐减缓。4043号菌种对应的GABA产量变化滞后于活菌数变化的现象,与植物乳杆菌WZ011(Lactobacillllus plantarumWZ011)在GYP培养基中的发酵动力学曲线(优化前后)类似[19]。当发酵时间为60 h时,虽然L-Glu-Na残留量显著小于48 h时的残留量(P<0.05),但GABA产量几乎没有增加,加之发酵时间过长会导致酸奶乳清析出增多,影响酸奶表观和感观评价。综合考虑生产成本和发酵周期,确定48 h为最适发酵时间。
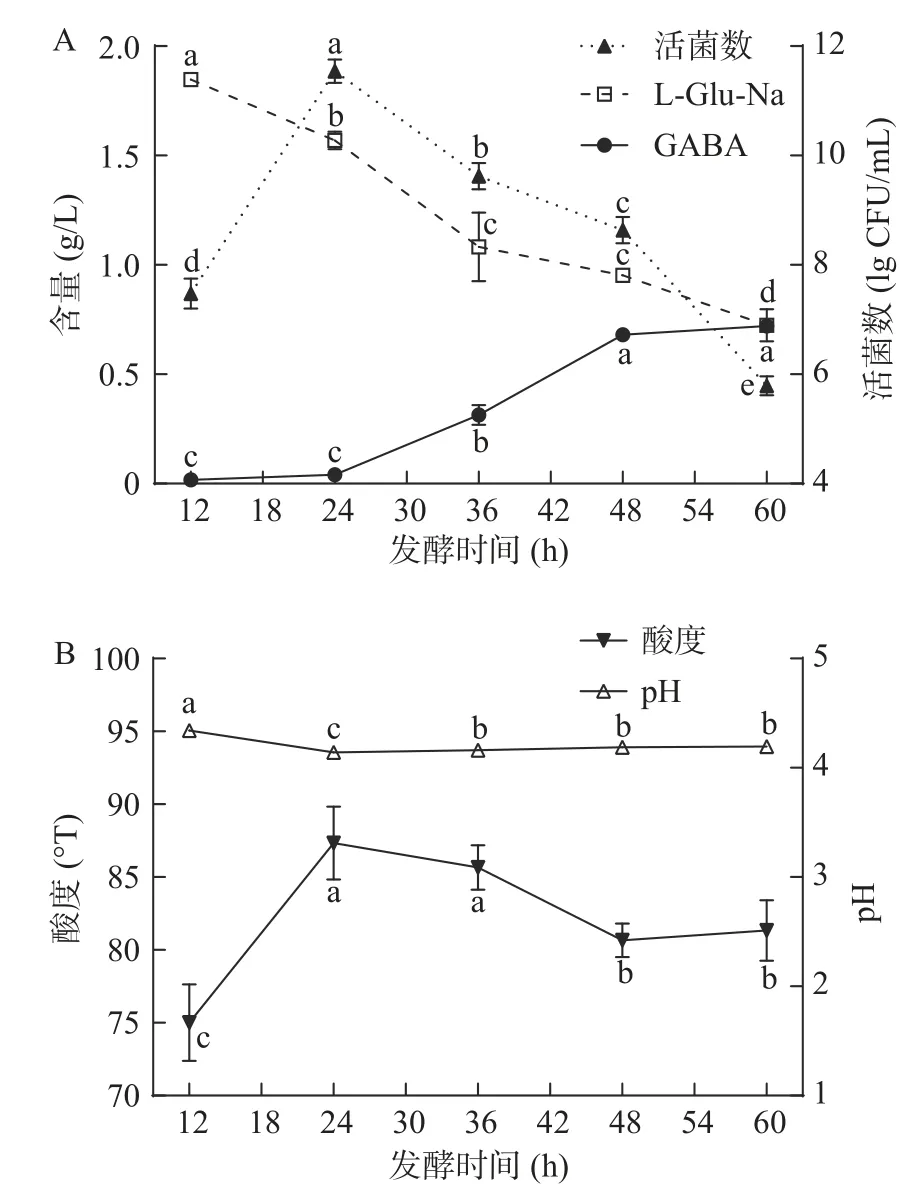
图2 发酵时间对单菌种发酵结果的影响Fig.2 Influence of fermentation timeon result of fermentation with single strain
2.1.3 最适发酵接种量的确定 接种量大小对活菌数和GAD酶总量有直接的影响。由图3可知,当接种量从2%(v/v)增至5%(v/v)时,GABA产量随着接种量增加而增大,至5%(v/v)时达到最高值约0.85±0.03 g/L。如接种量继续增加,GABA产量则显著降低(P<0.05)。究其原因,和活菌数、GAD与L-Glu-Na的相对量(浓度)变化有关。在发酵乳液中,底物即L-Glu-Na总量及浓度保持不变。当接种量处于2%~5%的范围内,活菌数和GAD总量相对较少,底物L-Glu-Na总量相对较高。此时增大接种量能够加快转化效率,提升代谢产物即GABA的产量。从图3A可以看出,该阶段又可分为两个区间:当接种量小于3.4%(v/v)时,GAD对L-Glu-Na的转化比例低于50%,导致L-Glu-Na残留量大于GABA产量,可称为“次优区间”;当接种量超过3.5%(v/v)时,GAD对L-Glu-Na的转化效率高于50%,GABA产量开始高于L-Glu-Na残留量,可称为“最优区间”。而当接种量超过5%(v/v)时,菌体浓度过大,无法与底物有效结合,菌株个体间竞争抑制了GAD的转化效率[20]。这导致GABA产量下降,L-Glu-Na残留量升高,加之发酵乳液内有限的营养物质不足以支持全体菌株生长,活菌数最终也会减少。可见,适宜的接种量更利于GABA产量积累。当接种量为5%(v/v)时,GABA产量最高,且L-Glu-Na残留量显著优于其余各接种量组(P<0.05)。故确定5%(v/v)为最适发酵接种量。
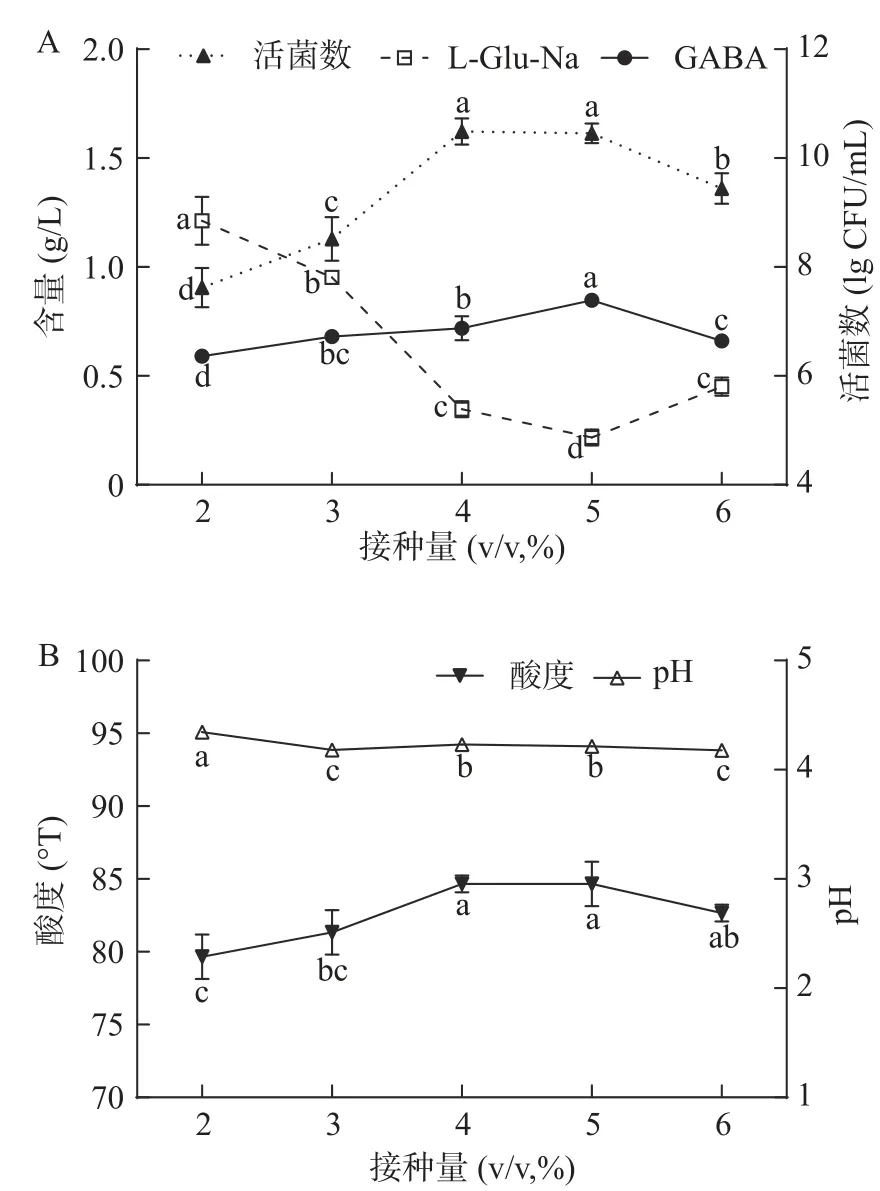
图3 接种量对单菌种发酵结果的影响Fig.3 Influence of inoculation quantity on result of fermentation with singlestrain
2.1.4 最适L-Glu-Na添加量的确定L-Glu-Na添加量与GABA产量、GAD活性及酸奶口感均关系密切。由图4可知,当L-Glu-Na添加量从0 g/L增大至4 g/L,GABA产量同步增加,至4 g/L时GABA产量达到最大值约1.03±0.03 g/L。而后随着L-Glu-Na添加量继续增大,GABA产量出现显著下降(P<0.05)。推测可能与GAD活性变化有关。由图4可知,L-Glu-Na添加量的变化对活菌数无显著影响(P>0.05),即GAD总量基本保持不变。而L-Glu-Na作为GABA的底物,不但可以影响GABA的生成量,还可通过调节GABA代谢支路中碳流量的变化,调节GAD活性[21]。当L-Glu-Na添加量介于0~4 g/L时,底物浓度相对较低,转化反应相当于酶促反应中一级反应或混合反应阶段,加之此阶段GAD活性随L-Glu-Na浓度升高而升高[5],GABA产量随之显著增加(P<0.05)。当L-Glu-Na添加量超过4 g/L时,底物浓度相对过高,将GAD饱和,转化反应受到反馈抑制,GAD酶活降低[14],导致GABA产量随之下降,L-Glu-Na残留量也因未能充分反应而迅速增加至约2.10±0.15 g/L,严重影响酸奶风味。综合GABA产量和感官评价,确定最适L-Glu-Na添加量为4 g/L。

图4 L-Glu-Na添加量对单菌种发酵结果的影响Fig.4 Influence of L-Glu-Na quantity on result of fermentation with single strain
根据课题组前期研究的结果,优化发酵条件前,将4043号菌以3%(v/v)接种于含L-Glu-Na 2 g/L的10%(w/v)脱脂复原乳中,在30℃下单菌种发酵48 h,GABA产量约为0.39±0.07 g/L。优化发酵条件后,当L-Glu-Na添加量为4 g/L、接种量为5%(v/v)时,4043号菌在34℃下于10%(w/v)的脱脂复原乳中恒温发酵48 h,GABA产量最高可达1.03±0.03 g/L,是优化前的约2.64倍。相比之下,赵树平等[22]从国产酸马奶中筛选的瑞士乳杆菌ND01(Lactobacillus helveticusND01),按3%(v/v)接种于11%(w/v)的灭菌复原乳中,在37℃下单菌发酵30 h,GABA产量最高约0.17 g/L。贤乾隆等[23]从国内酸菜和酸奶中分离得到的干酪乳杆菌QL-20(Lactobacillus caseiQL-20),按3%(v/v)接种至含L-Glu-Na 1%(w/v)的12%(w/v)的脱脂乳培养基中,37℃下单菌发酵48 h后,发酵乳中GABA产量约0.29 g/L。韩啸等[24]将从新鲜粪便中分离得到的短乳杆菌DL1-11(Lactobacillus brevisDL1-11)接种于(接种量6×107CFU/mL)蔗糖添加量5%(w/v)、L-Glu-Na添加量0.15%(w/v)的11%(w/v)的脱脂复原乳中,在35℃下单菌发酵9 h后,GABA产量最高达到101.20±2.48 mg/100 g。4043号菌在当前工艺条件下的GABA产量优于上述文献报道的产量,但L-Glu-Na残留量约0.69±0.09 g/L,残留量过高,导致酸奶综合感官评分很低。此外,课题组前期动物实验结果表明,GABA的有效剂量应不低于300 mg/d。若产品的摄入量为200 mL/d,则产品中GABA浓度应高于1.50 g/L。
为解决上述问题,可通过多种乳酸菌的复配和混合发酵,充分发挥菌种各自的发酵性能[25]。如薛玉清等[26]将从内蒙古传统酸奶中筛选的植物乳杆菌与德氏乳杆菌保加利亚亚种和嗜热链球菌(Streptococcus thermophilus)按照0.5:0.5:1.5的比例、总接种量为2%(v/v)接种至灭菌乳中,在43℃恒温混合发酵3.6 h,GABA产量超过了1.51 g/L。韩梅等[16]将光明乳业实验室保藏的嗜酸乳杆菌gm-11(Lactobacillus acidophilusgm-11)和嗜热链球菌gm-12在均质后的牛奶中混合发酵,在总接种量为1×106CFU/mL时,不同接种比例组在发酵终点的GABA产量均明显高于两种菌各自单菌发酵的产量,且当gm-11:gm-12为5:1时,GABA产量最高达5.62 g/L。Han等[27]将一株产GABA的嗜热链球菌与鼠李糖乳杆菌(Lactobacillus rhamnosus)按1:1(接种量均为1×106CFU/mL)接种于含L-Glu-Na为15 g/L的脱脂乳液中,在37℃下混合发酵48 h后,GABA产量达到8.3 g/L,是其单菌发酵时的2.96倍。可见,复配多菌种混合发酵有助于进一步优化工艺条件,显著提升发酵效率和GABA产量。
2.2 双菌种混合发酵单因素实验结果
2.2.1 双菌种复配方案及最适发酵温度的确定 课题组前期的研究结果表明,1060号菌、1134号菌和1443号菌均不具备GABA合成能力。将三种菌接种于发酵乳液中,在各自最适生长温度下(1060号菌的最适生长温度为37~45℃,1134号菌和1443号菌均为37℃[28])恒温发酵48 h后,酸奶的乳清析出控制均能满足GB 19302-2010《食品安全国家标准发酵乳》中的感官要求。参照前期预实验的结果,将三种菌分别与4043号菌按照1:1复配,进行双菌种混合发酵实验。基于2.1.1的实验结果及三种复配菌的最适生长温度,选择34和37℃以研究不同发酵温度对混合发酵GABA产量的影响,结果如表4所示。
由表4中可见,各复配组在34℃下的GABA产量和L-Glu-Na残留量均优于37℃下的结果,且组间均存在显著性差异(P<0.05),故确定34℃为多菌混合发酵的最适发酵温度。在34℃下,1134号复配组和1443号复配组的GABA产量均略优于4043号菌单菌发酵时的最大产量1.03±0.03 g/L,而L-Glu-Na残留量均远远小于4043号菌的0.69±0.09 g/L。推测是复配菌种间的共生作用所致[29]。如传统酸奶的发酵剂多由德氏乳杆菌保加利亚亚种和嗜热链球菌复配而成。在混合发酵时,嗜热链球菌生长性能好,产酸速率高,能够在发酵初期迅速创造酸性环境[30],并为德氏乳杆菌保加利亚亚种提供生长所需的叶酸[31]、甲酸[32]和CO2[33]等物质。而德氏乳杆菌保加利亚亚种的蛋白水解能力较强,能够生成多肽和氨基酸来促进嗜热链球菌的生长[34]。乳酸乳球菌乳酸亚种同样具备较强的利用乳糖酵解乳酸的能力[35],副干酪乳杆菌和干酪乳杆菌均具备良好的耐酸能力[36−37],复配组球菌、杆菌间具备互惠共生的基础,同时也应该具备增强菌种活力、提升L-Glu-Na向GABA的转化效率等效果。因1443号复配组在发酵终点的乳清析出控制优于1134号复配组,故确定1443号菌作为双菌种混合发酵的复配菌种。

表4 菌种复配和发酵温度的影响Table 4 Effects of strain collocation and fermentation temperature
2.2.2 最适发酵时间的确定 由图5可知,对于双菌种混合发酵,在12~48 h时阶段,GABA产量随发酵时间延长显著增加(P<0.05);随着发酵时间进一步延长,GABA产量反而有所降低。结合2.1.2可知,发酵温度对双菌种混合发酵GABA产量的影响与对单菌种发酵GABA产量的影响类似。当发酵时间为48 h时,GABA产量达到最高值,约1.05±0.02 g/L;L-Glu-Na残留量约为0.08±0.01 g/L,低于其在酸奶中的呈鲜阈值,且与其他发酵时间组间存在显著性差异(P<0.05)。综合考虑GABA产量与生产周期,选择48 h为多菌混合发酵的最适发酵时间。值得注意的是,在L-Glu-Na添加量和4043号菌种接种量相同的前提下,双菌种混合发酵的活菌数、酸奶酸度、GABA产量和产率均明显高于单菌种发酵相同发酵时段的各对应结果,而酸奶pH和L-Glu-Na残留量均明显低于各对应结果。这证明了2.2.1中有关双菌种混合发酵能够显著增强菌种活力、产酸能力及提升L-Glu-Na向GABA的转化效率等推论。

图5 发酵时间对双菌种混合发酵结果的影响Fig.5 Influence of fermentation time on result of mixing fermentation with double strains
2.2.3 最适总接种量的确定 由图6可知,总体而言,总接种量增大对双菌种混合发酵的GABA产量影响不大:当总接种量由4%(v/v)增至6%(v/v)时,GABA产量有显著增长(P<0.05);但在6~12%(v/v)区间,GABA产量基本稳定,略有波动。值得注意的是,自初始总接种量起,双菌种混合发酵始终处于“最优区间”内,GABA产量始终明显高于L-Glu-Na残留量。这表明在发酵乳液中,活菌和GAD的浓度始终低于L-Glu-Na的浓度。而GABA产量未跟随总接种量的增大而提升,应为乳酸乳球菌乳酸亚种GAD的pH活性范围较窄、pH依赖性较强所致[38]。双菌种混合发酵显著增强了菌种的产酸能力,使酸奶酸度明显升高,pH明显降低。当总接种量大于6%(v/v)时,酸奶pH已降至4.0以下,明显低于GAD的最适pH4.7[39−40],使其相对酶活低于50%[38]。此时继续增大总接种量,虽能提升GAD总量,但会使酸奶pH和GAD相对酶活进一步降低,导致总体转化效率和GABA产量变化不大。从生产成本的角度出发,总接种量6%(v/v)为宜。此时GABA产量约1.82±0.22 g/L,L-Glu-Na残留量约0.52±0.15 g/L。
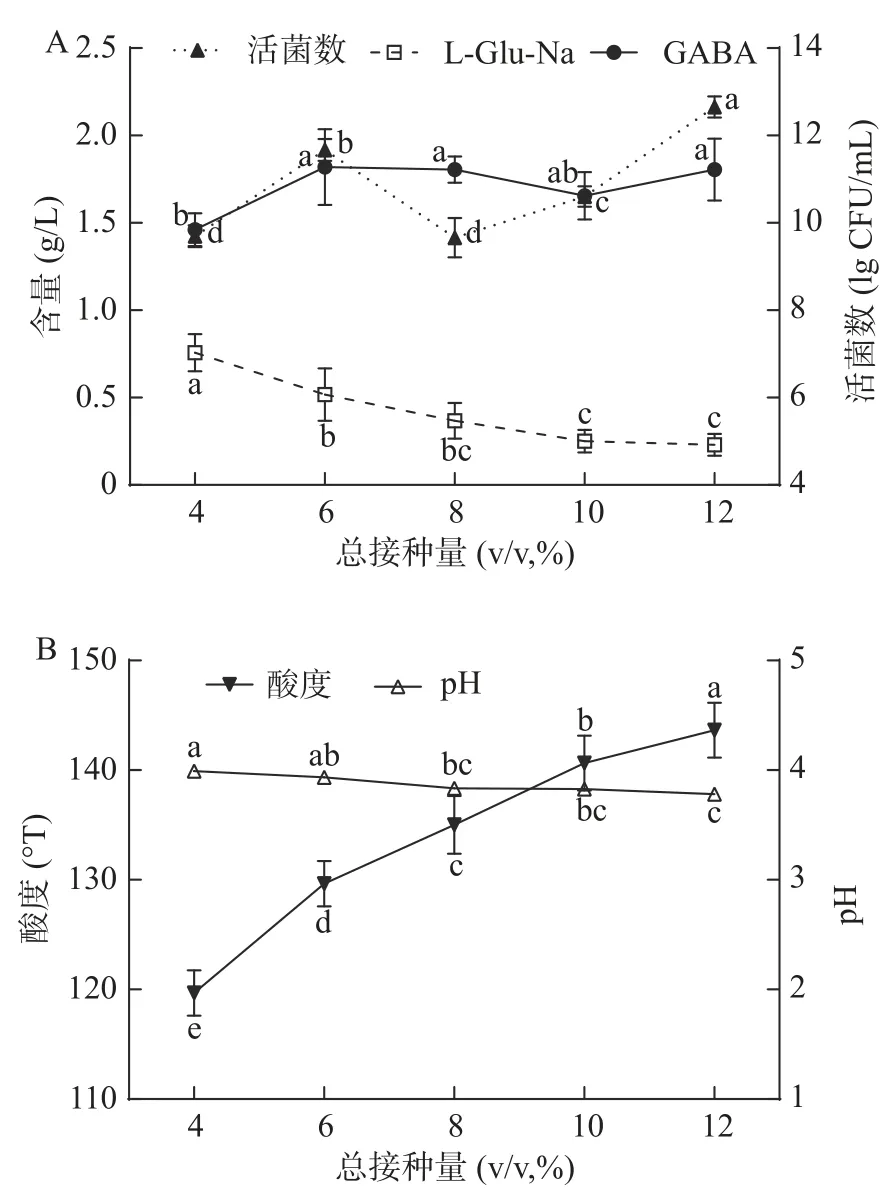
图6 总接种量对双菌种混合发酵结果的影响Fig.6 Influence of total inoculation quantity on result of mixing fermentation with double strains
2.3 均匀实验结果
U12(12×63)混合水平均匀设计实验的结果如表5所示。利用DPS 7.05软件对各指标进行二次多项式逐步回归分析,结果如表6所示。

表5 均匀实验结果Table 5 Results of uniform experiment

表6 指标模型Table 6 Model of varies parameters
其中,各因子对GABA影响主次顺序为X3(P=0.0004)> X32(P=0.0004)>X1X2(P=0.0055)> X42(P=0.0293)>X3X4(P=0.1273)>X2X4(P=0.1986)。选择Y2即GABA产量最大值对模型方程进行最值参数优化,计算可得,当发酵时间为48.42 h、L-Glu-Na添加量为8 g/L、发酵温度为32.23℃、球菌与杆菌配比为1:3时,发酵终点GABA理论产量约3.84 g/L。
2.4 验证性实验结果
根据模型方程最值参数优化结果,选择如下发酵工艺参数进行验证性实验:发酵时间为48 h、发酵温度为32℃、L-Glu-Na添加量为8 g/L、球菌与杆菌配比为1:3。进行3次平行试验,测得发酵终点酸奶中GABA实际产量为4.10±0.10 g/L,是优化前的3.78倍。L-Glu-Na残留量约0.11±0.01 g/L,低于其酸奶中的呈鲜阈值;活菌数为3.03×1010±1.75×1010CFU/g,酸度值为129.67±2.08°T,且酸奶各项指标均能够满足GB19302-2010《食品安全国家标准 发酵乳》中的相关规定,略优于模型预测值。不足是口感偏酸,综合感官评分约78.42±4.63分,属中上水平。该模型方程能够较为准确地分析各因素对双菌种混合发酵酸奶中GABA产量的影响。
3 结论
本研究利用课题组前期筛选得到的在乳基底物中具备高产GABA潜力的乳酸乳球菌乳酸亚种菌种与干酪乳杆菌复配,以脱脂复原乳液为主要原料,进行双菌种混合发酵实验。经单因素实验和均匀实验,确定最优发酵工艺参数为:发酵时间48 h、发酵温度32℃、L-Glu-Na添加量8 g/L、乳酸乳球菌乳酸亚种与干酪乳杆菌配比1:3。最终得到酸奶中GABA产量约4.10±0.10 g/L,是优化前的3.78倍,酸奶各项指标满足GB 19302-2010《食品安全国家标准发酵乳》中的相关规定,综合感官评分良好。该工艺简化了流程,降低了成本,GABA生产能力突出,可应用于富含GABA功能性酸奶的生产开发中。
猜你喜欢
化工管理(2022年13期)2022-12-02
中国典型病例大全(2022年7期)2022-04-22
河南畜牧兽医(2021年9期)2021-12-10
探索科学(学术版)(2021年2期)2021-04-22
中华养生保健(2020年2期)2020-11-16
农药科学与管理(2019年12期)2019-05-20
安徽科技(2018年9期)2018-10-24
中成药(2018年1期)2018-02-02
中国乳品工业(2015年12期)2015-12-16
中国药业(2014年24期)2014-05-26
