
甘蓝型油菜铵转运蛋白基因家族成员的鉴定与表达分析
2021-09-02 08:19:58岳彩鹏黄进勇华营鹏
华北农学报 2021年4期
周 婷,岳彩鹏,黄进勇,华营鹏
(郑州大学 农学院,河南 郑州 450001)



1 材料和方法
1.1 试验材料
供试材料为冬油菜品种湘油15,由湖南农业大学官春云院士提供。
1.2 试验方法
1.2.1 甘蓝型油菜AMT家族基因的鉴定 利用拟南芥AMT氨基酸序列为种子序列进行Blast分析,在白菜、甘蓝、甘蓝型油菜等植物中寻找AMT基因。在本研究中,使用了以下数据库:拟南芥信息资源数据库(TAIR)(https://www.arabidopsis.org/)用于拟南芥氨基酸序列查找,芸薹属数据库(BRAD)v.1.1(http://brassicadb.org/brad/)[32]用于白菜、甘蓝以及甘蓝型油菜氨基酸序列搜索。根据以下标准对芸薹属植物中的基因进行了命名:种名+染色体+同源基因的名称。例如,BnaA1.AMT1;4是指甘蓝型油菜A1染色体上拟南芥AMT1;4的同源基因。
1.2.2 AMT蛋白的分子特性鉴定 使用ExPASy ProtoParam(http://www.expasy.org/tools/protparam.html)[33]计算了各AMT蛋白的氨基酸数目和组成、分子质量(MW)、理论等电点(pI)、平均亲水系数(GRAVY)和不稳定性指数。利用THMMM(http://www.cbs.dtu.dk/services/TMHMM/)和TMpred(https://embnet.vital-it.ch/software/TMPRED_form.html)[34]预测氨基酸序列的跨膜结构域(TM)数目。
1.2.3 AMT家族基因进化树的构建 利用MEGA 7中的ClustalW功能,对20个甘蓝型油菜AMT蛋白序列、6个拟南芥AMT蛋白序列、9个白菜AMT蛋白序列和10个甘蓝的AMT蛋白序列进行比对,根据序列比对的结果,采用邻近法(NJ)构建系统进化树,Bootstrap值设定为1 000[35-36]。
1.2.4 AMT家族基因的结构分析 为了研究拟南芥和芸薹属作物AMT蛋白的结构差异,将所有的AMT蛋白序列置于MEME v5.1.1[37](http://meme-suite.org/tools/meme)中比对,以表征保守结构域。除了下列参数外,其余参数使用默认设置:最佳图案宽度设置为6~50 bp,最大保守结构域数量设置为10。使用2种不同的工具分析蛋白质的二级结构:GOR4[38](https://npsa-prabi.ibcp.fr/cgibin/page=npsagor4.html/)和PSIPRED[39](http://bioinf.cs.ucl.ac.uk/PSIPRED/)。利用在线网站Phyre2[40](http://www.sbg.bio.ic.ac.uk/phyre2/html)进行同源建模,以构建蛋白质的三维结构。
1.2.5 AMT家族基因的染色体定位分析 为了确定AMT基因在染色体上的位置和方向,将完整的AMT基因核苷酸序列进行BlastTn搜索,从而确定了AMT基因在每个染色体上的起始位置。使用MapInspect v.2010(http://www.soft.com/./MapInspect.html)作图,描绘AMT基因的基因组位置。串联重复基因是指100 kb基因组区域内的2个或更多基因的阵列[41]。

1.2.7 高通量转录组测序 对上述胁迫处理(1.2.6)的样品进行转录组测序,测序平台为Illumina Hiseq 2000,每个样品生成6.0 Gb测序数据,测序方式为双末端(PE)测序,读取长度为150 bp。以错误发现率(FDR)≤0.05和表达变化倍数≥2为鉴定差异表达基因的阈值。
1.2.8 组织表达分析 组织表达数据来源于甘蓝型油菜转录组信息资源数据库(BnTIR,http://yanglab.hzau.edu.cn/BnTIR)。数据包括甘蓝型油菜中双11的90个不同样品,覆盖了油菜生长过程不同阶段的不同器官。利用GraphPad Prism软件绘制箱型图。
2 结果与分析
2.1 二倍体白菜与甘蓝以及四倍体甘蓝型油菜AMT家族基因的鉴定
为了鉴定甘蓝型油菜中的AMT基因,使用拟南芥AMT的氨基酸序列作为参照,从芸薹属数据库(BRAD)中提取基因,并用油菜转录组数据检验这些基因的注释。进一步用SMART[43]软件对所有潜在基因所编码蛋白质的保守结构域进行校验,以保证每个蛋白质中都含有完整的AMT保守结构域,经过筛选,最终鉴定出20个BnaAMT基因:其中14个为AMT1家族成员:3个BnaAMT1;1s,2个BnaAMT1;2s、3个BnaAMT1;3s、3个BnaAMT1;4s、3个BnaAMT1;5s,其余6个为AMT2亚家族成员BnaAMT2;1s;并基于芸薹属植物基因命名标准进行了命名:BnaA5.AMT1;1~BnaCnn.AMT2;1。其中,Cnn表示该基因锚在C亚基因组上,但具体染色体信息未知;Unn表示该基因在油菜基因组的染色体信息完全未知。用同样的方法找到了白菜和甘蓝的AMT家族基因,白菜中共有9个AMT成员,甘蓝中共有10个AMT成员。分析了这39个基因所编码蛋白质的氨基酸数量、分子质量、等电点、不稳定系数、脂肪指数以及平均亲水系数。所有AMT基因编码蛋白质的氨基酸数量为330~669,其中33个介于475~512,差异较小(表1)。不稳定性指数>40表明蛋白质不稳定[44],分析表明这39个AMT蛋白均为稳定蛋白。

表 1 甘蓝型油菜、白菜、甘蓝AMT家族基因信息Tab.1 Basic information of AMT family genes in Brassica napus,Brassica rapa and Brassica oleracea
2.2 AMT家族基因系统发育分析
用20个甘蓝型油菜BnaAMTs蛋白序列、9个白菜BraAMTs蛋白序列、10个甘蓝BolAMTs蛋白序列和6个拟南芥AtAMTs蛋白序列构建系统进化树。AMT的每个亚家族成员都与拟南芥中的相应同源基因紧密聚集(图1),暗示甘蓝型油菜AMT家族基因与拟南芥相应同源基因具有相似功能。同时,甘蓝型油菜BnaAMTs均与白菜或者甘蓝的AMT成簇出现,拟南芥单独成一个分支(图1),这说明甘蓝型油菜与白菜和甘蓝基因编码的蛋白质在序列上的相似程度高于拟南芥,亲缘关系更近。
2.3 AMT家族基因的染色体定位
通过下载甘蓝型油菜AMT基因相关染色体定位信息,利用软件MapInspect进行染色体定位并进行可视化分析(图2)。结果显示,甘蓝型油菜的20个AMT基因不均匀地分布在12条染色体上:C4染色体上分布3个(BnaC4.AMT2;1a、BnaC4.AMT2;1b、BnaC4.AMT2;1c),A1、A5和C3染色体上均分布2个(BnaA1.AMT1;3、BnaA1.AMT1;4、BnaA5.AMT1;1、BnaA5.AMT2;1、BnaC3.AMT1;5a、BnaC3.AMT1;5b),A4、A7、A9、C1、C6、C7和C8染色体上均分布1个AMT基因(分别是:BnaA4.AMT2;1、BnaA7.AMT1;3、BnaA9.AMT1;2、BnaC1.AMT1;4、BnaC6.AMT1;1、BnaC7.AMT1;4、BnaC8.AMT1;1)。BnaCnn.AMT1;2(仅知定位在C亚基因组,具体染色体信息未知)、BnaUnn.AMT1;3(所在亚基因组和染色体信息未知)和BnaCnn.AMT2;1(仅知定位在C亚基因组,具体染色体信息未知)由于具体位置不详,无法进行染色体定位。其中,BnaC4.AMT2;1b和BnaC4.AMT2;1c在染色体上的相对位置仅为4 412 bp,推断它们为串联重复基因簇。
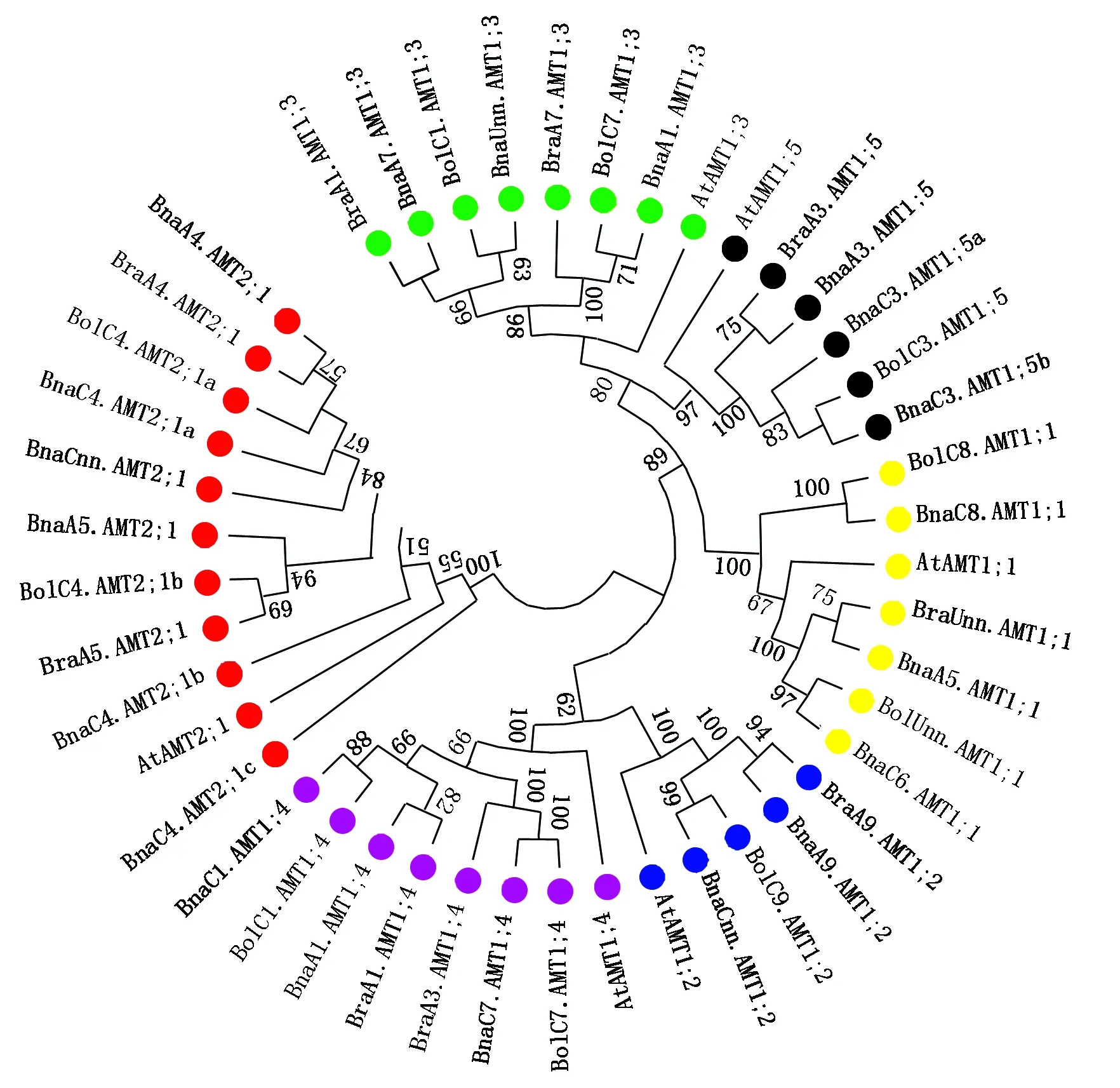
图1 甘蓝型油菜、白菜、甘蓝和拟南芥AMT家族蛋白质的系统进化分析Fig.1 Phylogenetic tree of AMT proteins in Brassica napus, Brassica rapa, Brassica oleracea and Arabidopsis

图2 甘蓝型油菜AMT家族基因在染色体上的分布Fig.2 Chromosomal location of Brassica napus AMT family genes
2.4 多序列比对和保守结构域分析
从分子进化的角度来看,如果氨基酸残基进化保守,则认为它们在功能上或结构上具有重要意义。序列同源性分析表明,甘蓝型油菜与拟南芥AMT蛋白具有高度相似性,表明AMT在异源多倍化过程中是保守的。为了揭示芸薹属植物和拟南芥AMT家族基因之间的内在联系,将45个AMT基因的氨基酸序列提交到MEME网站进行保守基序预测和注释。MEME分析表明,BnaAMT家族基因编码的氨基酸序列在芸薹属植物中基本含有相同的保守结构域(图3),表明它们高度保守。Motif是蛋白质分子中具有特定空间构象和特定功能的结构成分。它是结构域的亚单位,与特定的功能有关,推测具有不同Motif的AMT成员可能在进化和功能上存在差异。如,所有的BnaAMT2;1s基因均没有Motif1、Motif5和Motif6(图3),这可能是AMT2家族功能不同于AMT1家族的结构上的原因。AMT1和AMT2亚家族结构差异巨大,进化的过程中可能分开较早。

图3 甘蓝型油菜AMT家族基因及保守基序分析Fig.3 Diagram of structure of Brassica napus AMT family genes and their conserved Motifs
2.5 AMT家族蛋白的二级与三维结构的预测
BnaAMT蛋白的二级结构预测见图4。由于多重序列比对显示,AMT1亚家族和AMT2亚家族序列差异较大(图3),分别以BnaA5.AMT1;1和BnaA4.AMT2;1为例进行了二级与三级结构预测。GOR4和PSIPRED的二维结构预测结果相似。AMT家族主要由α-螺旋、延伸链与随机螺旋构成,AMT1家族α螺旋比例高于AMT2家族,而AMT2家族的延伸链与随机螺旋比例高于AMT1家族(图4)。3D轮廓突出了AMT蛋白的保守结构域,可以直观看出AMT蛋白结构中的α-螺旋、延伸链与随机螺旋(图5)。这些结果表明,随机螺旋和α-螺旋是这些AMT蛋白结构的重要组成部分。
2.6 BnaAMT基因的表达特性分析
2.6.1 BnaAMT基因的组织特异性表达 通过对甘蓝型油菜转录组信息资源库中AMT基因的分析,得到17个AMT基因(缺少BnaC3.AMT1;5b、BnaC4.AMT2;1c和BnaCnn.AMT2;1基因,在BnPIR(http://cbi.hzau.edu.cn/bnapus/)甘蓝型油菜泛基因组数据库中检索发现,这些基因在中双11油菜基因组中缺失)在8个组织中的表达数据。结果显示,不同基因在甘蓝型油菜不同器官的表达存在差异,进化树中处于同一分支的基因表达情况相似。例如,BnaA5.AMT1;1和BnaC6.AMT1;1基因在甘蓝型油菜除籽粒外的其他器官中表达量都相对较高,而BnaC8.AMT1;1基因在甘蓝型油菜所有器官都处于低表达状态(图 6)。根器官中表达较高的基因有BnaA5.AMT1;1、BnaC6.AMT1;1、BnaA9.AMT1;2、BnaA7.AMT1;3和BnaC3.AMT1;5a;茎器官中表达较高的基因有BnaA5.AMT1;1、BnaC6.AMT1;1、BnaA9.AMT1;2、BnaA1.AMT1;4和BnaC7.AMT1;4;叶器官中表达较高的基因有BnaA5.AMT1;1、BnaC6.AMT1;1、BnaA9.AMT1;2、BnaA1.AMT1;4、BnaC7.AMT1;4和BnaA5.AMT2;1;蕾器官中表达较高的基因有BnaA1.AMT1;4、BnaC1.AMT1;4、BnaC7.AMT1;4和BnaC4.AMT2;1b;花器官中表达较高的基因有BnaA5.AMT1;1、BnaC6.AMT1;1、BnaA1.AMT1;4、BnaC1.AMT1;4、BnaC7.AMT1;4、BraA3.AMT1;5和BnaC4.AMT2;1b;角果器官中表达较高的基因有BnaA5.AMT1;1、BnaC6.AMT1;1、BnaA1.AMT1;4和BnaC7.AMT1;4;角果壳器官中表达较高的基因有BnaA5.AMT1;1、BnaC6.AMT1;1、BnaA1.AMT1;4、BnaC7.AMT1;4、BnaA5.AMT2;1和BnaC4.AMT2;1b,籽粒中所有基因均呈低表达状态(图 6)。
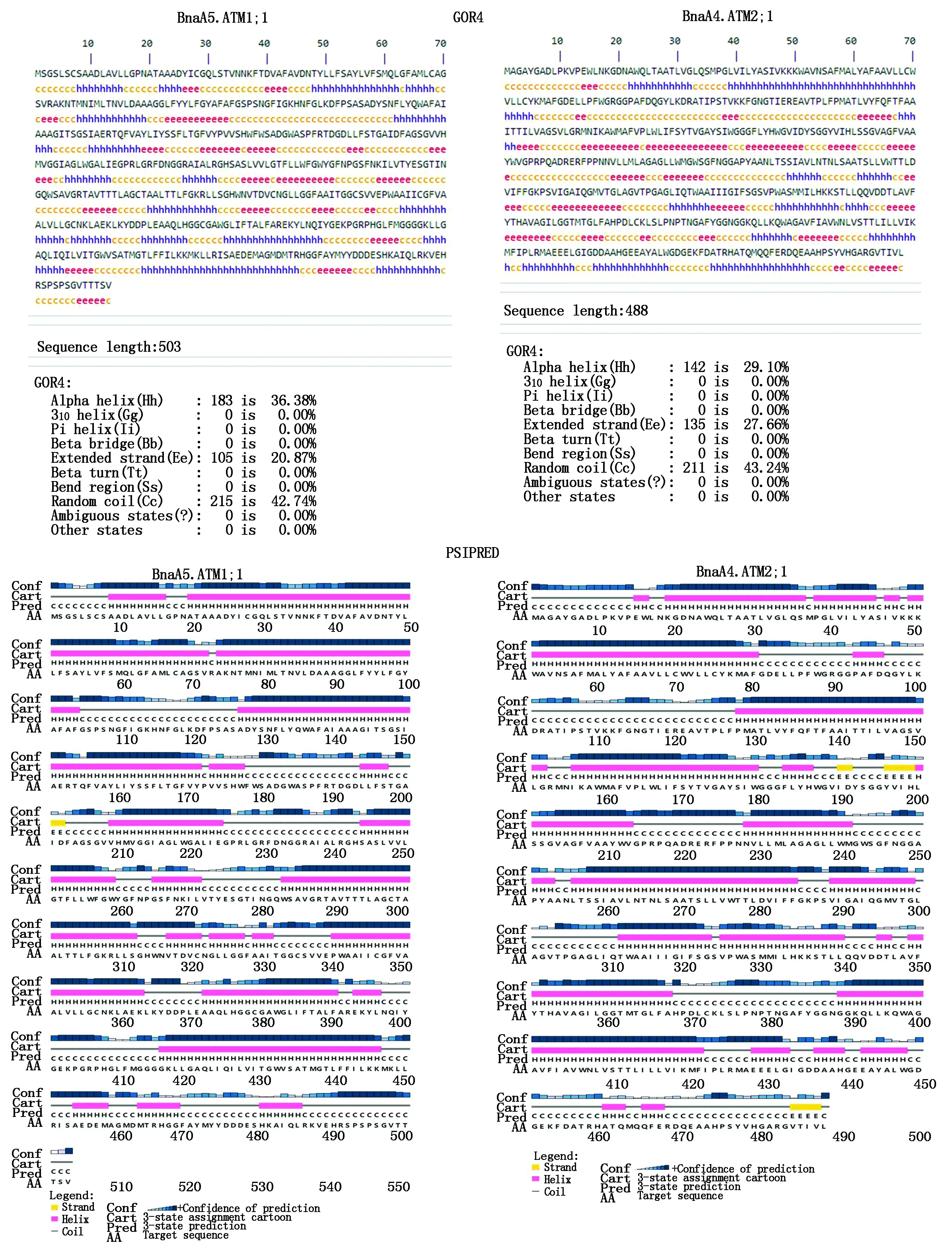
图4 甘蓝型油菜BnaA5.AMT1;1和BnaA4.AMT2;1蛋白二级结构分析Fig.4 Secondary structure analysis of BnaA5.AMT1;1 and BnaA4.AMT2;1 proteins

图5 甘蓝型油菜BnaA5.AMT1;1和BnaA4.AMT2;1蛋白三级结构预测Fig.5 Prediction of three-dimensional structure of BnaA5.AMT1;1 and BnaA4.AMT2;1 proteins
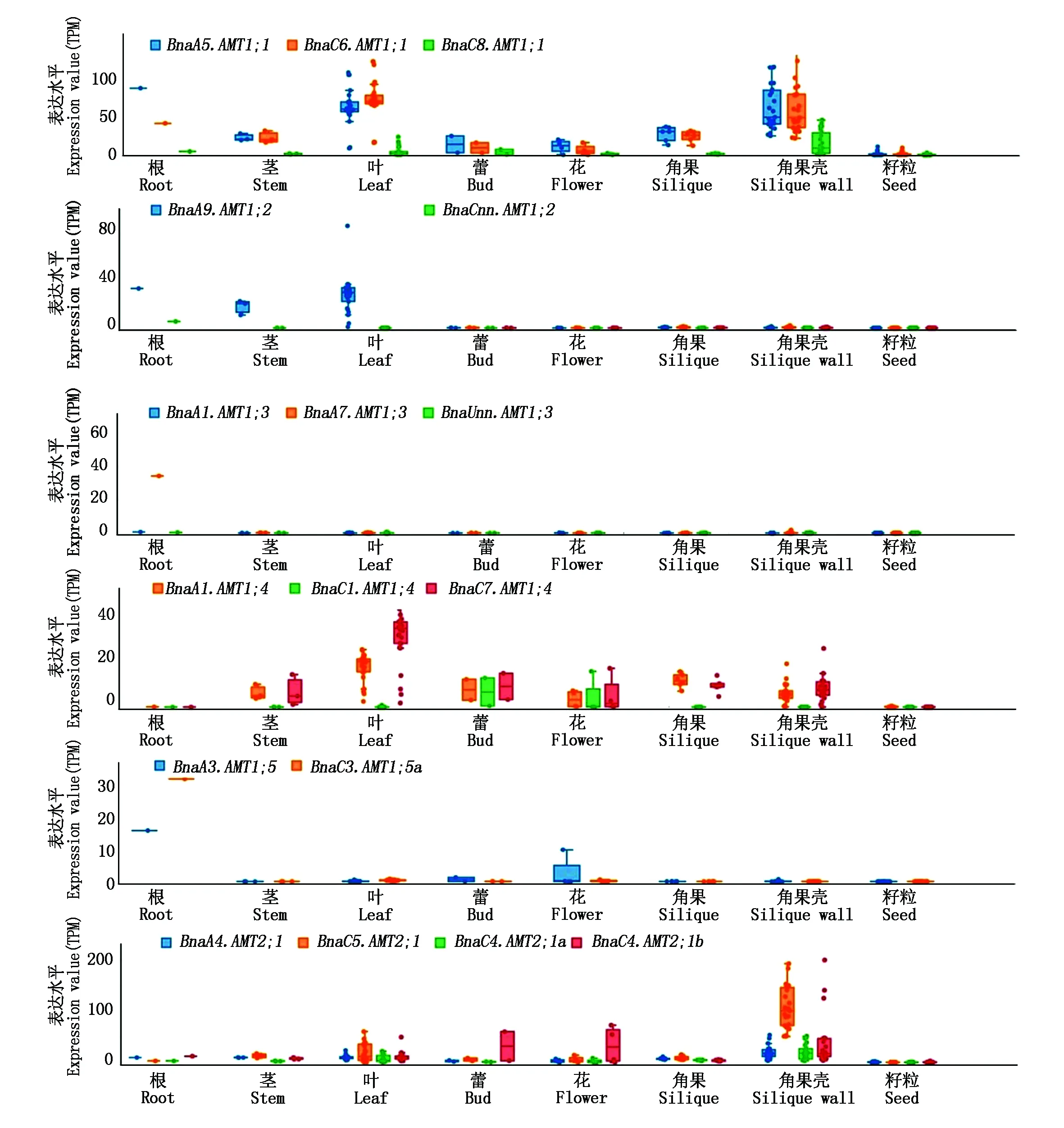
图6 甘蓝型油菜AMT家族基因在不同组织中的表达情况Fig.6 The expression of BnaAMT family genes in different tissues


S.地上部;R.根;差异显著性分析采用单因素方差分析,Tukey检验,图中不同字母表示处理之间差异显著(P<0.05)。图8同。S.Shoot;R.Root;The significance of difference was evaluated using one-way ANOVA,Tukey test,different letters in the same group indicate significant differences among treatments(P<0.05).The same as Fig.8.
2.6.3 BnaAMT对其他非生物胁迫的响应 随后,进一步分析了甘蓝型油菜AMT家族基因对其他营养胁迫与重金属胁迫,以及盐胁迫的转录响应情况,其中包括磷胁迫(缺磷)、硼胁迫(缺硼)、镉胁迫(镉毒害)以及盐胁迫(盐毒害)。在缺磷条件下,BnaC6.AMT1;1和BnaA9.AMT1;2在油菜地上部和根系中的表达量均显著上升,地上部BnaA7.AMT1;3、BnaA5.AMT2;1和BnaC4.AMT2;1c表达量显著上升,只有BnaC7.AMT1;4表达量显著下降,其他基因的表达量无显著变化(图8-A)。在缺硼条件下,根系中的BnaAMT1s 3个成员的表达水平均显著上升;地上部BnaC6.AMT1;1、BnaA9.AMT1;2的表达量显著上升,只有BnaC7.AMT1;4表达量显著下降(图8-B)。无论根部还是地上部,镉毒对AMT家族基因的表达影响较小(图8-C)。油菜受到盐胁迫时,地上部BnaA9.AMT1;2和BnaC4.AMT2;1c表达量显著上升,BnaC4.AMT2;1和BnaCnn.AMT2;1表达量显著下降,其他基因表达无显著变化;根部AMT家族基因整体上则呈现显著下降的趋势(图8-D)。总之,甘蓝型油菜在遭受缺硼或者缺磷胁迫时,AMT家族部分基因呈现上升表达趋势;在盐胁迫条件下,根系AMT家族基因整体表达量下降;镉毒对AMT家族基因的表达无明显影响。

图8 磷、硼、镉、盐胁迫下甘蓝型油菜BnaAMT家族基因的转录组分析Fig.8 Transcriptome responses of BnaAMT family genes to phosphorus,boron,cadmium and salt stresses
3 结论与讨论
本研究通过数据库比对,在甘蓝型油菜全基因组中发现了20个铵转运蛋白,可分为2个亚家族。其中14个为AMT1家族成员,其余6个为AMT2亚家族成员。由保守基序预测结果可知,甘蓝型油菜铵转运蛋白亚家族内的蛋白质序列保守性强,但亚家族间的差异较大。油菜在进化过程中,基因组发生了多次多倍化事件进而导致油菜内部普遍存在多拷贝基因家族[26-27],类似作物包括小麦(TriticumaestivumL.)[46]和棉花 (GossypiumhirsutumL.)[47]。因此,拟南芥中的每个基因在油菜中应有6个拷贝[48]。一个基因的多个拷贝的存在释放了任何单个拷贝以及自然和人为的选择压力,这促进了新基因功能的出现[49]。由此产生的变异的增加可以用于培育具有理想表型的作物品种[50]。因此,异源四倍体油菜AMT家族成员是否具有不同于拟南芥的新基因功能还有待进一步研究。

综上,本研究在甘蓝型油菜基因组中共发现了20个铵转运蛋白,其中14个为AMT1亚家族成员,6个为AMT2亚家族成员。由保守基序预测结果可知,铵转运蛋白亚家族内的蛋白质序列保守性强,但亚家族之间的差异较大。甘蓝型油菜AMT基因与模式植物拟南芥AMT的同源基因的表达模式既有相似之处,又存在差异。这表明在基因组多倍化过程中,AMT基因发生了功能分化,产生了适应油菜的新的基因功能。根部BnaA5.AMT1;1和BnaA7.AMT1;3,叶片BnaC7.AMT1;4和BnaA5.AMT2;1,角果壳BnaA5.AMT2;1表达量高,同时对不同氮水平响应显著,可以利用这些基因进行遗传操作以培育具有理想表型的油菜品种。在缺硼或者缺磷胁迫时,BnaAMT家族部分基因呈现上升表达的趋势;在盐毒害胁迫下,根系AMT家族基因整体表达量下降;镉毒对AMT家族基因的表达无明显影响。这些结果为进一步深入解析甘蓝型油菜AMT家族基因的分子功能奠定了基础。
猜你喜欢
中学生天地(A版)(2023年1期)2023-02-17 00:33:04
西藏农业科技(2019年3期)2019-11-04 00:35:14
西藏农业科技(2019年1期)2019-07-25 00:37:02
广州大学学报(自然科学版)(2019年1期)2019-05-07 01:33:26
生命科学研究(2018年1期)2018-05-29 01:12:47
西藏农业科技(2018年4期)2018-04-25 06:39:28
上海农业学报(2017年3期)2017-04-10 12:39:14
山东农业工程学院学报(2016年6期)2016-12-01 05:38:19
西南农业学报(2016年6期)2016-04-16 05:12:46
天津科技大学学报(2016年1期)2016-02-28 16:59:45
