
酶辅助乳化制备姜黄素脂肪乳研究
2021-08-27 09:29:26江连洲孙铭悦佟晓红成晓祎崔文玉
农业机械学报 2021年8期
江连洲 孙铭悦 佟晓红 成晓祎 崔文玉 王 欢
(东北农业大学食品学院, 哈尔滨 150030)
0 引言
姜黄素(Curcumin, CUR)是姜黄的天然多酚化合物,具有多种生物活性,例如抗氧化、抗增殖、抗炎和抗癌特性。姜黄素对pH值和温度较敏感,故易于氧化和化学降解,其生物利用度较低,导致其应用受到极大的限制[1]。因此,需要采取一些有效的手段将这种化合物应用于食品中,并实现营养递送。文献[2]制备了姜黄素纳米乳液,研究表明纳米乳液可以实现姜黄素的包埋和运载,且稳定性较好。文献[3]研究表明,在140℃下将过量的姜黄素添加到一定量的香精油中溶解,不会引起姜黄素的降解。这些为酶法辅助制备负载姜黄素的新型大豆脂肪乳提供了基础。
基于乳液的递送系统在生物活性营养素运载中起着重要作用。常规乳液是热力学不稳定的体系,存在许多物理化学机制(如奥斯特瓦尔德熟化、絮凝、乳化和聚结),随着贮藏时间的延长,其稳定性逐渐降低[4]。脂肪乳是一种将脂质以小液滴状分散在水中,形成非均相的水包油(O/W)型乳液,能够明显提高疏水生物活性物质在水溶液中的溶解度和生物利用度[5]。近年来,研究表明,将脂肪乳作为溶剂与某些活性物质组合能达到控释的目的[6]。文献[7]利用大豆卵磷脂作为乳化剂,以大豆油为油相,通过高速剪切和高压均质制备了姜黄素脂肪乳。
制备乳液低耗能方法之一的相转变温度法是利用体系内部发生相变所释放的化学能实现乳化,然而,相比于超声、高压均质和研磨法,这种相转变温度法制备的乳液稳定性差、使用局限性较大。文献[8]利用生物酶酶解大豆粉过程中得到的优质组分蛋白(或肽)和磷脂作为天然乳化剂,吸附在油、水界面,并降低界面张力,可以形成天然且稳定的水包油(O/W)型乳液体系,这种低能乳液即为相转变温度法产生的乳液。但有关利用生物酶辅助相转变温度法制备脂肪乳作为生物活性物质递送体系尚未见深入的研究。
本文选用姜黄素作为亲脂性生物活性物质,利用碱性蛋白酶辅助酶解膨化大豆,将姜黄素溶解在大豆酶解物中,利用水解后的蛋白、磷脂等作为天然乳化剂,制备自发形成、稳定性更佳的新型脂肪乳。为解决姜黄素生物利用率低等问题,本文对以酶解制备大豆乳液为原料负载姜黄素的脂肪乳进行评估,并对其包埋率随储存时间的变化进行研究,以期实现姜黄素更高的稳定性能和生物利用度。
1 材料与方法
1.1 材料与试剂
全脂膨化大豆(蛋白质量分数40%、油脂质量分数20%),山东高唐蓝山集团总公司;Alcalase 2.4 L碱性蛋白酶,诺维信(中国)生物技术有限公司;姜黄素、胃蛋白酶、胰蛋白酶、猪胆酸盐、尼罗蓝、尼罗红、1,1,3,3-四乙氧基丙烷,美国Sigma公司;其他试剂均为分析纯。
1.2 仪器与设备
Mastersize2000型激光粒度分析仪,英国伍斯特郡马尔文仪器有限公司;F-4500型荧光分光光度计,日本日立公司;722型可见分光光度计,上海光谱仪器有限公司;THZ-80型水浴恒温振荡器,天津常仪仪器设备有限公司;LGR20-W型台式高速冷冻离心机,北京京立离心机有限公司;TCS SP8型激光共聚焦显微镜,日本岛津制作公司。
1.3 方法
1.3.1脂肪乳制备
参照文献[9]的方法,全脂大豆经挤压膨化后按照液料比6 mL/g进行水解,水浴55℃恒温搅拌。用2 mol/L NaOH溶液调pH值至9.0。通过Alcalase 2.4 L碱性蛋白酶(质量分数1.5%)酶解90 min。随着酶解进行,每9 min加入固定量的姜黄素,使添加的姜黄素质量分数分别达到0、0.2%、0.4%、0.6%、0.8%、1.0%。经中性条件下灭酶5 min。冷却,室温(20℃)、9 000 r/min条件下离心20 min,分离得到负载姜黄素的膏状脂肪乳,记为E0、E0.2、E0.4、E0.6、E0.8、E1.0。
1.3.2脂肪乳微观结构观察
参照文献[10]的方法,采用激光共聚焦显微镜进行微观结构观察,激发光的波长分别是488 nm和633 nm。样品稀释至20 mg/mL。配制1 mg/mL的尼罗红染液和10 mg/mL的尼罗蓝染液(异丙醇作溶液),分别对油脂和蛋白染色,时间为40 min。染色后的样品置于载玻片,盖上盖玻片密封。用100倍油镜采集样品图像。
1.3.3脂肪乳粒径、电位分析
参照文献[11]的方法,采用马尔文激光粒度仪测定乳状液体积平均粒径及其分布情况,将乳状液用去离子水稀释1 000倍,充分振荡,颗粒折射率为1.520,分散剂折射率为1.333,操作温度为25℃。电位测定前采用蒸馏水将不同姜黄素添加量的脂肪乳稀释1 000倍,混匀。稀释后的样品置于Zeta池中。平衡120 s后,在25℃下测定电位,每个样品重复3次测量。
1.3.4乳化活性及乳化稳定性
乳化活性及乳化稳定性的测定参照文献[12]的方法加以修改。将不同姜黄素添加量的脂肪乳用0.1% SDS(十二烷基硫酸钠)溶液稀释到确定倍数,在波长532 nm处用紫外分光光度计测定吸光度,计算乳化活性指数(EAI)。静置30 min后测定吸光度,计算乳化稳定性指数(ESI),公式分别为
(1)
(2)
式中E——乳化活性指数,m2/g
S——乳化稳定性指数,min
N——稀释倍数,取250
C——乳化液形成前蛋白质水溶液中蛋白质质量浓度,g/mL
φ——乳化液中油相体积分数,%
A0——0 min时的吸光度
A30——30 min时的吸光度
1.3.5氧化稳定性
(1)POV值
参照文献[13]的方法,采用硫氰化铁法测定过氧化值(POV值),脂肪乳氢过氧化物含量的测定是将样品置于45℃干燥箱中加速氧化,并于0、3、5、7、14、21 d取出分装小样进行氧化实验的测定,以不添加乳液的一组为对照。
(2)TBARS值
次级氧化产物——丙二醛含量的测定采用硫代巴比妥酸反应物(TBARS)法,由文献[14]的方法稍作改动。将2 mL样品和4 mL硫代巴比妥酸试剂混合于试管中,混匀后沸水浴加热15 min,冷却至室温,4 000 r/min离心20 min,经0.22 μm水系微孔滤膜过滤,测定上清液在532 nm波长处的吸光度。样品中TBARS值可以通过1,1,3,3-四乙氧基丙烷标准曲线计算。
1.3.6荧光光谱分析
脂肪乳的荧光光谱分析根据文献[15]方法并进行了修改,将脂肪乳用PBS(10 mol/L磷酸盐缓冲液、pH值7.0)稀释至蛋白质质量浓度为0.15 mg/mL,涡旋混合20 s后采用F-4500型荧光分光光度计测定荧光图谱。设置激发波长为295 nm,发射波长为300~500 nm,狭缝宽度均为5 nm,重复3次实验。
1.3.7姜黄素包埋率
测定包埋率需对不同姜黄素添加量的脂肪乳总姜黄素含量和表面姜黄素含量进行定量,本文分别对储存第0天、第7天、第14天的脂肪乳姜黄素包埋率进行测定。绘制标准曲线(R2>0.999),并计算对应姜黄素含量。
根据文献[16]进行修改,测定脂肪乳中姜黄素包埋率:将定量脂肪乳分散在DMSO(二甲基亚砜)中,并涡旋振荡2 min,静置后在432 nm处测定吸光度,根据姜黄素标准曲线计算出姜黄素的浓度和样品的表面姜黄素含量。取适量脂肪乳溶解在DMSO中,涡旋振荡2 min,在60 W条件下超声振荡萃取10 min后,以5 000 r/min离心5 min。收集上清液,洗涤沉淀3次。收集洗涤清液在432 nm处测定吸光度。根据姜黄素标准曲线计算出样品中总姜黄素含量。脂肪乳的姜黄素包埋率计算公式为
(3)
式中E′——样品的姜黄素包埋率,%
C0——表面姜黄素质量浓度,mg/mL
C′——总姜黄素质量浓度,mg/mL
1.3.8体外模拟消化分析
参照文献[17]的方法,使用胃蛋白酶和胰蛋白酶对脂肪乳进行胃肠消化模拟。用磷酸盐缓冲液(0.1 mol/L、pH值7.0)稀释样品至所需倍数,与模拟胃液(SGF)以体积比1∶1混合,并使用1 mol/L HCl将pH值调节至3.0,在消化混合物中加入胃蛋白酶使之活性达到2 000 U/mL,将混合物在水浴振荡器(37℃、120 r/min)中反应2 h。SGF消化2 h后,取出胃消化样品与模拟肠液以体积比1∶1混合,用1 mol/L NaOH将pH值增加至7.0,添加胰蛋白酶(活性为100 U/mL),混合物在相同条件下孵育2 h。调节体系pH值使之始终为6.9~7.0。分别收集胃消化和肠消化后消化样品,以测定在体外消化后粒径和姜黄素生物利用率。
1.3.9姜黄素生物利用率
参照文献[1]的方法,测定经体外模拟胃肠消化后的样品中姜黄素含量来计算姜黄素生物利用率。分别取适量胃消化和肠消化结束后的消化液于4℃用5 000 r/min低温离心10 min后取清液层。利用紫外分光光度法,测定姜黄素浓度,根据标准曲线分别计算出胃、肠消化后清液层的姜黄素含量。脂肪乳中姜黄素的生物利用率计算公式为
(4)
式中Y——姜黄素生物利用率,%
C1——清液层中姜黄素质量浓度,mg/mL
C2——消化前样品中姜黄素质量浓度,mg/mL
1.3.10数据统计及分析
所有数据均为3 次重复试验结果,表示为平均值±标准差。采用Origin 2019软件进行图表绘制,SPSS 23.0用于数据分析,显著性水平为P<0.05。
2 结果与讨论
2.1 脂肪乳微观结构观察
激光共聚焦显微镜可观察乳液中油脂和蛋白的分布情况,形成O/W乳液后可观察到圆形液滴,如图1所示。结果表明,添加姜黄素后,脂肪乳液滴变小。添加量为0.6%时,乳液中油滴和蛋白分布比较均匀,且蛋白包裹在油滴外侧,信号较强。姜黄素添加量在0.8%和1.0%时,脂肪乳平均液滴大小没有显著变化,但液滴分布不均匀,少量的油滴变大,游离在外侧,可能的原因是蛋白有少量聚集或吸附到乳液界面,影响乳液界面蛋白油脂分布情况。文献[18]的研究结果表明,蛋白质分子之间的疏水相互作用导致其他蛋白质(在水相中未吸附的)迁移到O/W界面,影响乳液界面组成。
2.2 脂肪乳粒径、电位分析
流体动力学半径分布是评价乳液稳定性的重要指标,可以直观地评价液滴的分散状态。本文利用动态光散射技术探究不同姜黄素添加量下脂肪乳液滴分布情况及体积平均粒径D4,3。酶法辅助制备的大豆脂肪乳粒径分布如图2a所示。结果表明,脂肪乳粒径主要呈现单峰分布,分布较为均匀。从图2b(图中不同字母表示数据之间差异显著(P<0.05))可知,未添加姜黄素的脂肪乳粒径最大,添加姜黄素后,随添加量的增加,粒径逐渐增大,这可能是因为在油相中所含的姜黄素导致乳液粒径变大[19]。同时,在添加量为0.6%时脂肪乳平均粒径显著增加,达到1 127.42 nm。此添加量下峰宽较窄,表明此时脂肪乳中颗粒大小均匀。姜黄素添加量0.8%和1.0%的脂肪乳平均粒径增加不显著,分别为1 145.29 nm和1 159.23 nm,峰宽较大,脂肪乳颗粒大小不均一。文献[19]的研究也发现,包埋姜黄素的乳液粒径随着姜黄素含量的增加而增加,达到一定浓度时乳液平均粒径逐渐趋于稳定。
电位绝对值可间接评价乳液的稳定程度。由图2c可知,在添加姜黄素后,电位绝对值随着姜黄素添加量增加先增加再减小。其中,在添加量为0.6%时达到38.67 mV,此时乳液分子间静电斥力较大,更易保持结构稳定[20]。
2.3 乳化活性及乳化稳定性
乳化活性及乳化稳定性可评价乳液的乳化性能[21],结果表明,由于蛋白(多肽)和磷脂的共存体系作为乳化剂形成的液滴较稳定,利用酶辅助法制备大豆脂肪乳的乳化稳定性较高。如图3(图中不同大写字母表示脂肪乳乳化活性指数之间差异显著(P<0.05),不同小写字母表示脂肪乳乳化稳定性指数之间差异显著(P<0.05))所示,随着姜黄素添加量的增加,乳化活性指数和乳化稳定性指数均呈显著降低的趋势,可能的原因是加入姜黄素会对乳化剂吸附到O/W界面的能力产生负面影响。在添加量为0.6%时,EAI和ESI分别为92.39 m2/g和189.54 min,当添加量在0.8%和1.0%时乳化稳定性指数和乳化活性指数有显著降低(P<0.05),表明添加过多的姜黄素会影响乳液的乳化性能,可能的原因是其他蛋白质迁移到O/W界面,与CLSM观察到的现象相吻合。文献[22]指出,在弱酸性或接近中性pH值条件下,过量蛋白质吸附在界面层会导致乳化活性的降低。
2.4 氧化稳定性分析
本文采用测定乳液在45℃干燥箱中经0、3、5、7、14、21 d储存后的氧化产物含量来评价不同姜黄素添加量脂肪乳的氧化稳定性,分别测定了脂肪乳的初级氧化产物含量(POV值)和次级氧化产物含量(TBARS值)。
由图4a可知,所有样品在第0天至第7天时的POV值没有显著增加,且不同姜黄素添加量的脂肪乳之间没有显著差异。第7天后,所有样品POV值显著增加,第14天添加姜黄素0、0.2%、0.4%、0.6%、0.8%和1.0%的乳液POV值分别增加至6.215、5.56、3.99、3.15、2.71、3.05 mmol/kg。添加量为0.6%的脂肪乳在第14天时的POV值较低,仅为3.15 mmol/kg,表明此条件下脂肪乳稳定性好,氧化产物较少,不易受到氧气的破坏。姜黄素添加量达到0.8%和1.0%时与0.6%样品相比POV值差异不显著,表明较高姜黄素添加量脂肪乳氧化程度均较低,且随着储藏时间增加,受到姜黄素添加量的影响较小。
次级氧化产物丙二醛生成物(TBARS值)的结果如图4b所示,在氧化初期所有样品的TBARS值没有显著差异。在经过7 d后,部分初级氧化产物转化为次级氧化产物,乳液的TBARS值有所增加,但均不超过0.50 mmol/kg。文献[23]采用高速剪切制备姜黄素MCT/Pickering乳剂,在第7天的TBARS值约为0.65 mmol/kg,由此可见,酶法辅助制备的大豆姜黄素乳液具有较传统高速剪切制备的乳液更好的氧化稳定性。酶法辅助制备过程中蛋白质(多肽)吸附在油滴表面后形成一层黏弹性蛋白膜,可以有效阻止脂质氧化的引发剂扩散到油滴内部,同时限制水相中的过渡金属与油相相互作用,从而起到抑制油脂氧化的作用[13]。脂肪乳E0、E0.2、E0.4、E0.6、E0.8和E1.0在第14天的TBARS值分别增加至0.70、0.55、0.45、0.32、0.43、0.37 mmol/kg,表明在姜黄素添加量较少时,乳液的氧化稳定性受到姜黄素添加量的影响,且添加量为0.6%的脂肪乳TBARS值最低,与POV值的结果一致。在储存21 d时,姜黄素添加量为0.6%脂肪乳的TBARS值仅为0.56 mmol/kg,产生较少的次级氧化产物。当姜黄素添加量在0.6%以上时,脂肪乳的TBARS值没有明显变化,表明过多的姜黄素添加量不会显著抑制脂肪乳的次级氧化产物的生成。
2.5 荧光光谱分析
脂肪乳外层水相中的大豆蛋白含有疏水性氨基酸残基(色氨酸和酪氨酸),是蛋白质的主要荧光基团,蛋白质具有的内源荧光对疏水性的色氨酸和酪氨酸极性变化敏感[24]。本研究测定了姜黄素添加量在0、0.2%、0.4%、0.6%、0.8%和1.0%的脂肪乳内源荧光光谱。
如图5所示,脂肪乳水相中的大豆蛋白在发射波长332 nm处具有最大荧光强度,说明色氨酸所处的微环境具有较高的疏水性。由图5可知,未添加姜黄素的脂肪乳荧光强度达到3 300.32,加入姜黄素后,E0.2、E0.4和E0.6的最大荧光强度分别为2 880.85、2 514.10和2 013.24,表明姜黄素的添加使蛋白质的色氨酸微环境疏水性增加,此时荧光强度稍有降低。然而,姜黄素添加量增加至0.8%和1.0%时,脂肪乳水相中的蛋白质最大荧光强度显著降低至1 787.17和1 710.64,表明姜黄素含量的增加使大豆蛋白发生内源荧光猝灭现象。文献[25]的研究结果表明,荧光光谱显示蛋白质和酚类物质的相互作用可导致荧光猝灭。同样地,文献[26]的研究表明,姜黄素添加量的增加会促进大豆蛋白与姜黄素的相互作用,发生静态荧光淬灭。因此,在姜黄素添加量大于0.6%时,未包埋在油相中的游离姜黄素可能会在酶法制备脂肪乳的过程中与大豆蛋白通过非共价作用相结合,从而间接降低其包埋率和生物利用率。
2.6 姜黄素包埋率
姜黄素的包埋率以及生物利用率是乳液作为生物活性物质递送体系的关键评价指标。姜黄素的包埋率会随着乳液的凝聚、聚集和氧化分解等因素发生变化。本研究测定了不同姜黄素添加量脂肪乳储存第0天、第7天、第14天的姜黄素包埋率,如图6(图中不同大写字母表示不同姜黄素添加量乳液原始包埋率之间的差异显著(P<0.05),不同小写字母表示储存过程中姜黄素包埋率之间的差异显著(P<0.05))所示。
结果表明,原始(第0天)乳液E0.2、E0.4和E0.6的姜黄素包埋率分别为66.62%、68.30%和72.58%,表明酶法辅助制备的大豆脂肪乳可以较好地负载姜黄素,且随着姜黄素的增加,包埋率增大。文献[27]中开发了用于包裹姜黄素的羧甲基玉米醇溶蛋白纳米粒子,包埋率为66.70%,可见酶法制备的大豆脂肪乳相较于传统纳米乳液体系有较高的包埋率。然而,当继续添加姜黄素至添加量0.8%和1.0%时,姜黄素包埋率显著降低至56.70%和30.54%,表明过量添加姜黄素并不会使姜黄素包埋率增加,反而会对包埋效果产生较大负面影响。可能的原因是制备的大豆脂肪乳中蛋白和磷脂吸附的界面层十分稳定,过量的游离姜黄素无法进入并包埋在乳液的油相中。文献[28]中表示,不同的芯壁比在固定的乳化体系中会造成表面电荷的变化,从而影响乳化体系中颗粒的大小和分布,间接影响包埋率和稳定性,并推测在芯壁比增加到某一值后,乳化体系固定且表面电荷的变化不明显。
在乳液储存第7天时,E0.2、E0.4和E0.6的姜黄素包埋率由66.62%、68.30%和72.58%下降到65.87%、67.23%和70.34%,并没有大幅度降低,表明此时乳液发生了轻微的氧化,间接影响了对姜黄素的包埋效果。而姜黄素添加量为0.8%和1.0%的乳液包埋率有显著降低,可能的原因是在贮存过程中,脂肪乳内部有少量姜黄素暴露于乳液表面,且游离姜黄素随添加量的增加而增多,导致包埋率有显著降低。
当储存第14天时,E0.2、E0.4和E0.6的姜黄素包埋率分别为63.56%、64.37%和68.45%,没有显著降低,且总体姜黄素包埋率均在60%以上;E0.8和E1.0的姜黄素包埋率分别降低至47.61%和20.60%,表明随着储存时间的增加乳液发生的聚集、凝聚和氧化分解使姜黄素的包埋率显著降低。因此,当姜黄素添加量大于0.6%时,添加过量姜黄素的脂肪乳不能在储存第14天后保留更多的姜黄素,反而会降低脂肪乳对姜黄素的保护作用。文献[29]的研究表明,大豆蛋白-姜黄素-大豆多糖复合物中在20 h后有56%~60%的姜黄素释放。同样地,文献[30]报道了随着储存时间的延长,姜黄素在蛋白质水解产物纳米乳剂中的流失率达到36.26%~44.66%。对比可知,在姜黄素添加量合适的情况下,酶法辅助制备的大豆脂肪乳相比常规的姜黄素负载方法有较好的包埋效果。
2.7 姜黄素生物利用率
生物利用率可衡量乳液运载体系经胃肠消化后的递送效果,生物活性物质会不断在胰脂肪酶和猪胆酸盐作用下水解,并被载入到由胆盐和脂肪酸组成的混合胶束中,进而被小肠上皮细胞吸收[31]。因此可以通过测定胶束中姜黄素的浓度得到生物利用率。本文测定了不同姜黄素添加量脂肪乳在胃肠消化后的姜黄素生物利用率。由表1可知,姜黄素在胃肠消化后的生物利用率与脂肪乳粒径呈负相关。
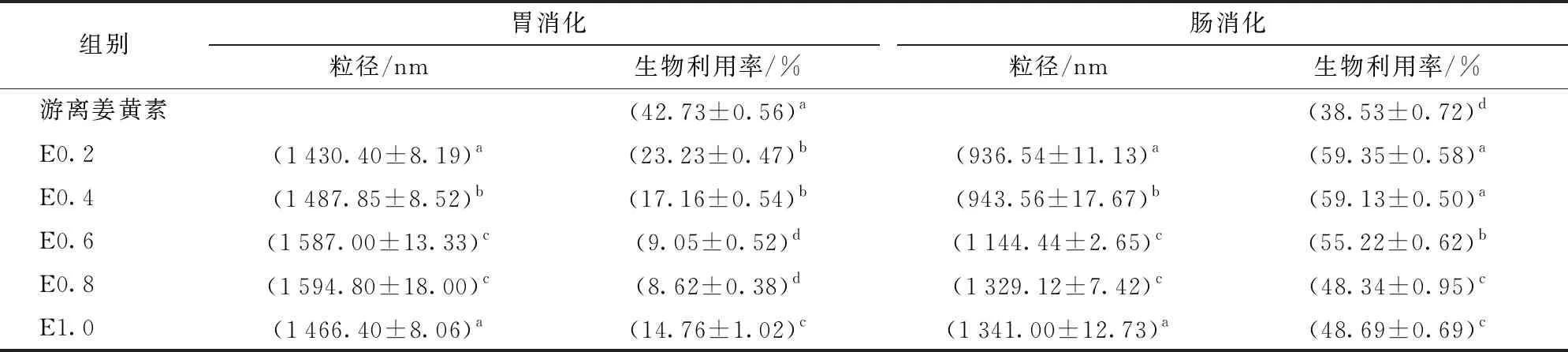
表1 不同姜黄素添加量脂肪乳经胃肠消化后的粒径、生物利用率
经SGF消化2 h后,胃蛋白酶水解诱导疏水域的暴露,这可能导致脂肪乳聚集,平均液滴大小显著增加[3]。未经脂肪乳包裹的姜黄素释放消化了42.73%,E0.2、E0.4和E1.0分别消化了23.23%、17.16%和14.76%的姜黄素,表明经过脂肪乳包埋的姜黄素释放量显著降低。E0.6仅消化利用了9.05%的姜黄素,表明此姜黄素添加量可以将姜黄素在胃消化液中的释放控制在较低水平。文献[32]提出,当在pH值7.2的PBS中孵育时,超过30%的游离姜黄素仅需30 min即可降解。
经SIF(模拟肠液)孵育4 h,胰蛋白酶分解了来自胃消化液的大液滴导致粒径减小。游离姜黄素在肠液中的生物利用率较低,仅为38.53%,而脂肪乳中的姜黄素利用率显著增加,E0.2、E0.4、E0.6、E0.8和E1.0分别消化利用了剩余的59.35%、59.13%、55.22%、48.34%和48.69%的姜黄素,表明经肠道消化液中胰蛋白酶的作用打破了原有的乳液结构,消化了油相中的姜黄素。文献[33]的研究结果也表明,蛋白质水解产物乳化剂可以显著促进姜黄素在纳米乳液中的释放,生物利用率达到53.24%。综合胃肠消化情况,姜黄素添加量在0.6%时,酶法制备的新型大豆脂肪乳可以较好地包埋姜黄素,且在SGF消化液中有较低的释放量,在SIF中有较大的生物利用率,表明该包封达到了控制释放的效果。
3 结束语
以膨化大豆为原料,利用酶法辅助制备了姜黄素添加量0、0.2%、0.4%、0.6%、0.8%、1.0%的脂肪乳。研究表明,添加姜黄素的脂肪乳粒径随着姜黄素含量增加而增大,添加量达到0.6%时乳液粒径不再显著增加,此时电位绝对值最大。乳液在姜黄素添加量达到0.6%时,乳液蛋白包裹油滴外侧,没有游离的油滴,此时脂肪乳的乳化稳定性和乳化活性较高。姜黄素的添加会降低脂肪乳的氧化程度,在添加量大于0.6%时对脂肪乳的氧化程度影响微弱。荧光光谱结果表明,添加过量姜黄素会使未包埋的姜黄素以游离状态与大豆蛋白结合,发生荧光淬灭,导致包埋率和生物利用率降低。在添加量为0.6%时,脂肪乳对姜黄素的包埋率高达72.58%,高于传统纳米乳液。体外消化实验表明,酶法辅助制备的姜黄素脂肪乳运载体系具有高效的缓释效果。因此,酶法辅助制备的大豆脂肪乳构建的姜黄素运载体系可实现对姜黄素更好的包埋及利用,具有广阔的应用前景。
猜你喜欢
中国药科大学学报(2022年3期)2022-07-02 06:10:40
实用癌症杂志(2019年9期)2019-09-07 01:26:16
Medical Data Mining(2019年2期)2019-07-16 04:36:08
中成药(2018年2期)2018-05-09 07:19:43
中成药(2018年3期)2018-05-07 13:34:37
中国果菜(2016年9期)2016-03-01 01:28:39
中国洗涤用品工业(2015年11期)2015-02-28 19:03:09
实用中西医结合临床(2015年7期)2015-02-28 16:30:41
西南军医(2015年1期)2015-01-22 09:08:25
济宁医学院学报(2014年4期)2014-08-16 13:44:19
