
蓖麻生长素外输载体PIN蛋白家族的生物信息学分析
2021-08-25 07:24郑钧伦李子彤门淑珍
聊城大学学报(自然科学版) 2021年5期
郑钧伦,罗 琼,李子彤,门淑珍
(南开大学 生命科学学院,天津 南开 300071)
0 引言
生长素(auxin)广泛存在于各种植物中,是最早在植物体被发现并进行研究的激素之一。虽然结构简单,但其能实现的功能却十分复杂,在植物生长和发育的许多过程中都发挥着至关重要的作用。生长素通过在不同的组织建立浓度梯度影响植物细胞的伸长、分裂与分化等过程,从而调节植株的向重力性和向光性生长,调控植物组织器官的发育和分化,如胚的形成、器官发生和形态建成、花芽的萌发以及果实的发育与成熟等[1,2]。一般认为,生长素在茎尖、根尖和正在发育的幼叶等生长活跃的组织中合成,通过维管系统以被动运输或主动运输的方式运送至目标组织和器官。研究表明,植物产生生长素浓度梯度主要是通过主动运输,该运输方式就是生长素所特有的极性运输(polar auxin transport,PAT)机制。
在极性运输的作用下,植物的一些发育特性开始出现,比如维管发生、顶端优势和向性生长等等[2]。一般生长素在植物中的极性运输主要指茎尖向茎基部,以及茎基向根部的运输。进一步的研究表明,生长素极性运输除了上述运输途径外,还包括在组织内部的局部定向运输,例如生长素在根的表皮细胞和侧根冠细胞中的向基(从根尖向根基部)运输及其通过皮层细胞向根尖的回流[3,4]。
生长素浓度梯度分布决定了植物的生长发育模式,而生长素的转运蛋白对于生长素的梯度分布起着关键作用。因此,对生长素极性运输的载体蛋白及其调节机制的研究是生长素研究的热点。截至现在,人们已经鉴定出几种生长素转运蛋白,包括AUXIN1/LIKE-AUX1(AUX / LAX)、PIN-FORMED(PIN)、ABCB(ATP-binding cassette transporters B sub-family)转运蛋白和类PIN蛋白(PIN-LIKES,PILS)转运蛋白[3,5]。这些生长素的载体蛋白根据亚细胞定位及功能不同可分成两类:一类定位于细胞质膜,作为生长素的输入或输出载体负责细胞间的生长素极性运输,包括AUX/LAX家族蛋白、大部分PIN家族蛋白以及ABCB转运蛋白;另一类主要定位于内质网膜,维持细胞内生长素的稳态[3],包括PILS以及PIN蛋白的短链亚族成员。
PIN蛋白由两侧的疏水区与一个位于中央的亲水区构成。研究最多的是拟南芥AtPIN家族,有8个成员(AtPIN1-AtPIN8),由于最先发现的atpin1突变体花序呈针状(pin-formed)而得名。后根据亲水环长度及亚细胞定位不同将其分为三个亚族:第一类包括AtPIN1-AtPIN4 和AtPIN7,它们都具有长的亲水环,极性定位于质膜;第二类包括AtPIN5和AtPIN8,具有短的亲水环,定位于内质网膜;AtPIN6属于第三类,其具有较长的亲水环,具有质膜和内质网膜的双重定位[5,6]。研究表明,PIN基因家族是从同一祖先基因进化而来,其成员之间的蛋白结构的差异主要来自于其中央亲水环的差异,并通过对中央亲水环不同的磷酸化位点修饰使AtPIN蛋白家族成员功能各异[6-8]。长链家族的AtPIN1-AtPIN4以及AtPIN7的功能研究比较深入,它们都极性定位于细胞膜,将胞内的生长素运输至细胞外,形成与维持组织中生长素的浓度梯度[2-5,9]。PIN1蛋白主要定位于维管组织细胞的底部细胞膜,参与生长素的极性运输,同时影响器官发生、维管分化、花芽和叶脉的形成[2-5,9];PIN2蛋白在根尖伸长区和胚中表达,也参与极性运输,调控植物的向重力生长[10]。PIN1、PIN3、PIN4和PIN7蛋白也参与根的向重力生长和茎的负向重力生长[11]。此外,PIN3和PIN4蛋白还参与控制植物的向光性生长,并与早期侧根的发育有关[12]。PIN5蛋白定位于内质网,负责生长素从胞质到内质网的运输,通过调节细胞质内的生长素浓度而参与侧根生长、早期胚的发生以及根与子叶的生长等一系列与生长素调控有关的植物发育过程[13];PIN6蛋白由于磷酸化作用的影响,使它既可以定位于细胞膜也可以定位于内质网,通过两种定位方式来同时调节胞内外的生长素极性运输与胞质间的生长素动态平衡, PIN6蛋白也参与了由生长素信号转导决定的植物发育过程,如主/侧根的发育和生长、根毛的生长以及顶端优势的形成等等[14]。PIN8蛋白定位于内质网,参与花粉、雄配子体以及孢子体的形成[15]。
迄今为止,人们已经通过全基因组测序在三十余种植物中鉴定出PIN基因家族,成员数目在4(地钱,Marchantiapolymorpha)至23(大豆,Glycinemax)之间[3]。遗传学分析证明,在进化过程中的基因组重复是导致PIN基因在不同物种中的数量差异的主要原因。尽管人们对PIN蛋白家族的研究在不断深入,许多植物的PIN基因也不断被发现,但一些需要受到研究关注的植物却一直没有研究。蓖麻(RicinuscommunisL.)是大戟科(Euphorbiaceae)蓖麻属(Ricinus)下的唯一物种。它原产于非洲,目前在中国、印度和巴西均有大面积栽培。作为世界十大油料作物之一,蓖麻产出的蓖麻籽可以用于提炼榨取蓖麻油,其不仅可以作为生物柴油,而且还是化工领域中重要的原材料之一。在人类对生态环境保护日益重视的今天,由于蓖麻的含油量高、油质好且具有一定的可再生性,被认为是最有希望的石油替代者[16]。
本研究以功能已基本明确的拟南芥PIN蛋白家族序列为参考,在蓖麻基因组数据库中检索蓖麻编码PIN蛋白的同源基因序列,并通过生物信息学方法系统分析目的基因的序列信息,以及该基因家族的进化关系和保守基序特征,旨在为将来开展蓖麻PIN蛋白家族的生物学功能研究提供参考信息,为蓖麻生长素的极性运输机制研究奠定基础,为蓖麻种植行业的发展提供理论依据。
1 实验方法
1.1 实验材料
蓖麻种子由南开大学生命科学学院的李秀兰老师提供,于南开大学生命科学学院试验田种植。剪取水培蓖麻幼苗的根、土中正常生长植株二叶期的茎和叶、盛花期植株的雌花、雄花和果实组织用于提取表达分析所用的总RNA。
1.2 实验方法
1.2.1 总RNA的提取和cDNA第一链的合成。采用北京全式金生物技术有限公司RNA提取试剂盒(ET121)提取蓖麻各组织材料的总RNA。采用北京全式金生物技术有限公司反转录试剂盒(AE301)合成cDNA第一链。
1.2.2RcPIN基因在蓖麻各组织的表达。以上述获得的cDNA为模板,利用RcPIN基因的特异引物进行RT-PCR。利用蓖麻的RcACTIN7基因作为内参。引物由生工生物工程(上海)股份有限公司合成,引物序列见表1。

表1 RT-PCR引物序列
1.2.3 生物信息学分析工具。实验使用的序列数据来自于TAIR数据库、NCBI数据库和Phytozome数据库。通过生物信息学手段分析蓖麻中编码PIN蛋白的同源基因,使用的生物信息学分析工具如表2。

表2 生物信息学分析工具
2 实验结果
2.1 蓖麻PIN蛋白家族的序列分析
为了检索蓖麻中编码PIN蛋白的同源序列,以拟南芥的8条PIN蛋白序列为模板,在蓖麻基因组数据库中进行比对,删除掉得分较低的亚型,共获得7条蓖麻PIN蛋白序列,对其逐一进行分析。
理化性质分析结果显示,蓖麻PIN蛋白平均长度约为537个氨基酸残基,平均分子量为58.64 ku。编码氨基酸最长的是RcPIN3蛋白,长度为646个氨基酸残基,预测分子量为70.39 ku;编码氨基酸最短的是RcPIN8蛋白,长度为356个氨基酸,预测分子量为39.05 ku。蓖麻PIN蛋白的理论等电点变化范围在8.17(RcPIN3)至9.52(RcPIN8)之间,均大于7,平均理论等电点为8.84,结果如表3所示。不稳定系数分析结果显示,RcPIN1-1、RcPIN1-2、RcPIN2、RcPIN3、RcPIN5、RcPIN6、RcPIN8蛋白不稳定系数均小于40,为稳定蛋白。它们的脂溶指数都较高,证明它们脂溶性较好。亲水性系数分析结果显示,根据亲水性系数正值为亲水性,负值为疏水性,介于-0.5至+0.5之间为两性蛋白的原则[39],RcPIN5、RcPIN8蛋白为疏水性蛋白,其余蛋白均为两性蛋白。保守结构域分析结果显示,除了RcPIN5、RcPIN8只有一个Mem_trans保守结构域外,其余PIN蛋白均具有两个Mem_trans保守结构域,结果如表3所示。

表3 蓖麻PIN蛋白家族成员信息
通过比对拟南芥和蓖麻的PIN蛋白序列,构建进化树,然后进行基因结构分析,结果显示,拟南芥与蓖麻聚集到一起的基因均有较为相似的基因结构,如图1所示。RcPIN1-1、RcPIN1-2与AtPIN1均有5个内含子,6个外显子。AtPIN2有8个内含子,9个外显子,而RcPIN2有5个内含子,6个外显子。RcPIN3、AtPIN3、AtPIN4和AtPIN7均有5个内含子,6个外显子。RcPIN5和AtPIN5均有4个内含子,5个外显子。RcPIN6与AtPIN6均有6个内含子,7个外显子。AtPIN8与RcPIN8均有5个内含子,6个外显子。

图1 拟南芥与蓖麻PIN蛋白家族的进化关系与基因结构
对蓖麻PIN蛋白的二级结构进行分析,结果如表4所示。从二级结构分析结果可以看出,它们的二级结构主要由α螺旋、延伸链、β转角和无规则卷曲组成。其中RcPIN1-1、RcPIN1-2、RcPIN2、RcPIN3和RcPIN6的二级结构的数量表现为无规则卷曲>α螺旋>延伸链>β转角,而RcPIN5和RcPIN8则表现为α螺旋>无规则卷曲>延伸链>β转角,β转角是这些蛋白中数量最少的结构。
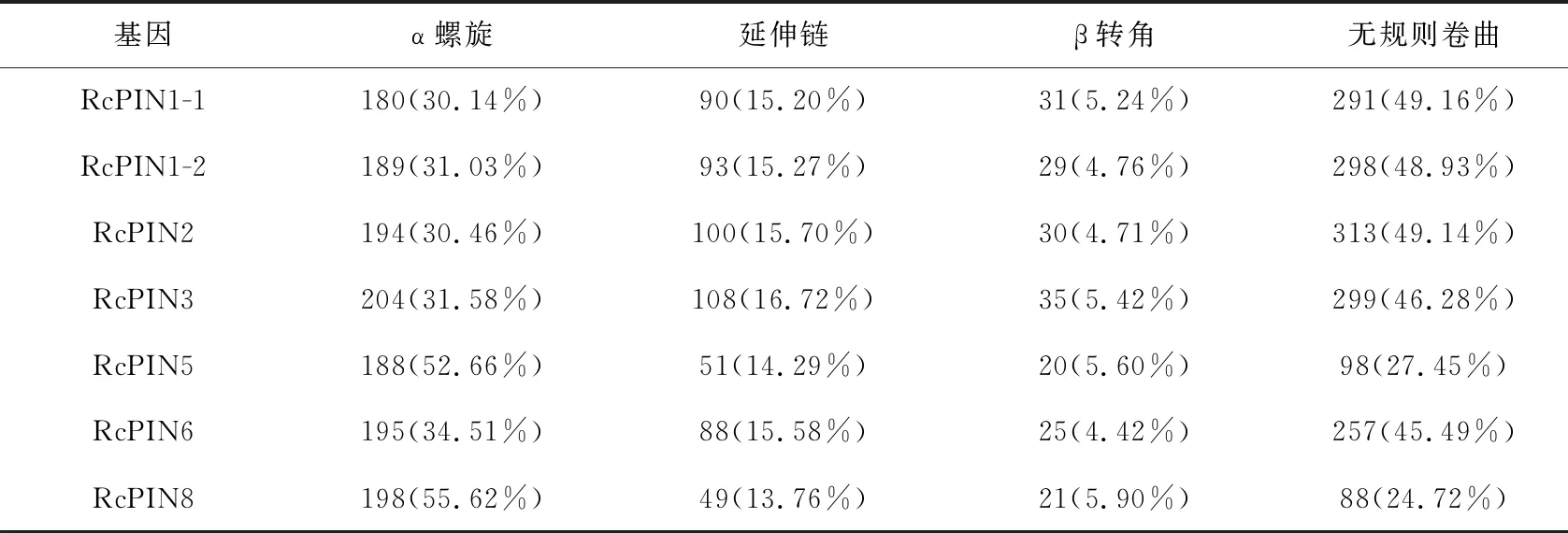
表4 蓖麻PIN蛋白家族的二级结构
PIN蛋白主要负责将生长素转运至胞外,是植物细胞重要的跨膜蛋白,因此跨膜结构域是其重要特征。跨膜结构域分析结果显示,所有AtPIN家族与RcPIN家族成员均有N端和C端两个疏水区域(5次跨膜)和一个中央的亲水环(图2)。亲水环结构位于细胞内。RcPIN1-1、RcPIN1-2、RcPIN2、RcPIN3以及RcPIN6蛋白的亲水环较长,而RcPIN5和RcPIN8蛋白的亲水环较短。分别与AtPIN1、AtPIN2、AtPIN3、AtPIN6、AtPIN5以及AtPIN8对应,拓扑结构相似性非常高(图2)。
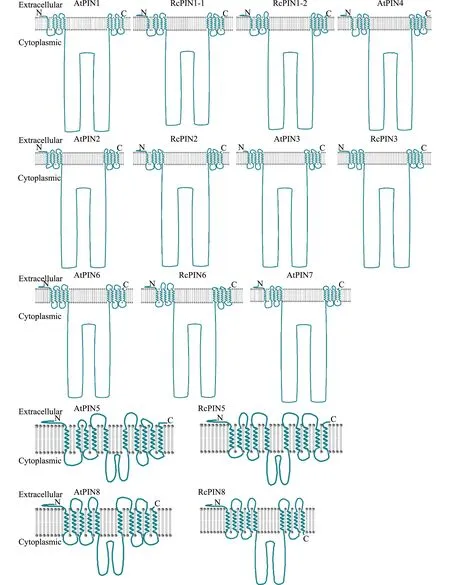
图2 拟南芥与蓖麻PIN蛋白家族的跨膜结构域
使用SignalP 5.0在线工具预测拟南芥与蓖麻PIN蛋白的信号肽,结果发现这15个PIN蛋白均不存在信号肽(分析结果未展示),暗示PIN蛋白不会向细胞外表达,为非分泌性蛋白。PIN蛋白的磷酸化对其细胞膜极性定位和转运活性有重要影响,因此,使用NetPhos 3.1在线工具分析了拟南芥与蓖麻PIN蛋白的磷酸化位点。结果显示,与拟南芥类似,蓖麻PIN蛋白的磷酸化位点也大多分布于蛋白序列中间的亲水环区域,潜在的磷酸化位点多数为丝氨酸残基和苏氨酸残基,少数为酪氨酸残基(图3)。
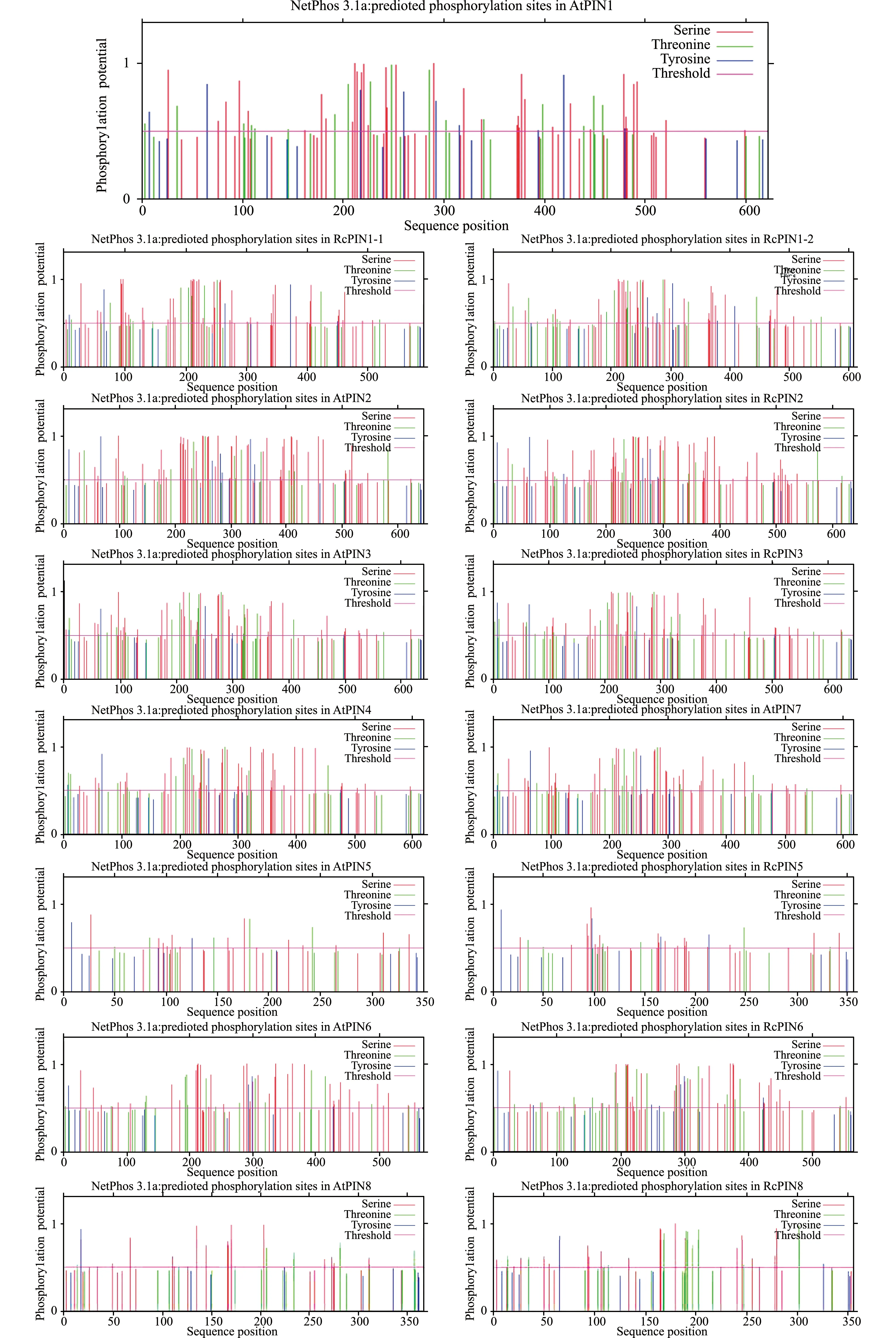
图3 拟南芥与蓖麻PIN蛋白家族的磷酸化位点预测
使用Mafft Version 7在线工具进行序列比对后用Jalview软件绘制出序列比对分析图,结果如图4所示。结果表明,PIN家族蛋白的两端跨膜结构域较为保守,而构成中央亲水环的氨基酸序列差异较大。
2.2 蓖麻PIN基因家族的系统进化分析
选择双子叶植物烟草,大戟科的木薯、橡胶树和麻风树,单子叶植物水稻和玉米作为分析对象,与蓖麻和拟南芥PIN蛋白序列一起置于MEGA7.0软件中使用邻接法构建进化树。结果显示,蓖麻与同科的木薯、橡胶树和麻风树聚在一起,表示PIN蛋白在大戟科植物中序列比较保守。拟南芥和烟草等双子叶植物聚拢于一起,而单子叶植物水稻和玉米聚拢于一起。此结果与植物分类学一致,结果如图5所示。
2.3 蓖麻PIN基因的表达分析
为了初步研究所筛选出的PIN蛋白的功能,我们首先利用蓖麻基因表达数据库(RcDB,https:∥woodyoilplants.iflora.cn/),对这些PIN基因的表达模式进行了分析。结果显示,RcPIN1-1和RcPIN1-2基因在胚和根中的表达水平较高,而且RcPIN1-1基因在胚中的表达水平明显强于其它组织(图6),暗示其在胚发育中有重要的功能。RcPIN2基因在根中特异表达。RcPIN3基因在所有检测的组织中均有表达,且在幼苗、茎、叶片和幼嫩的种子中表达水平较高。RcPIN5基因主要在叶、根和茎中表达,而且在叶片中表达水平明显强于其它组织。在该数据库中未检索到RcPIN6基因的表达。而相对于其它的蓖麻PIN基因,RcPIN8在所有组织中的表达均较弱(图6)。
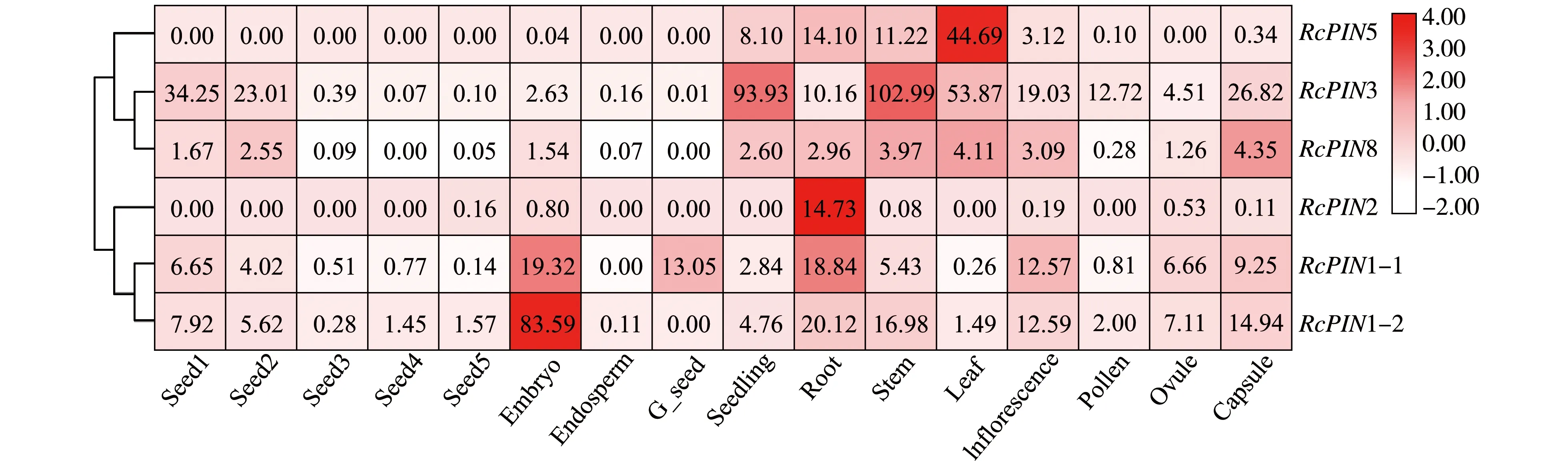
注:比例尺表示标准化后的相对表达水平,无色框表示较低的表达水平,有色框表示基因的转录水平较高;种子1-5表示种子的不同发育阶段(10-50天)。
我们进一步利用RT-PCR对这些RcPIN基因的组织表达水平进行了验证,实验结果与上述数据库的结果基本吻合(图7)。RcPIN1-1和RcPIN1-2基因在根、叶、雌花和幼嫩的种子中表达水平较高;RcPIN2基因主要在根中表达;RcPIN3、5、6和8基因在所有检测的组织均有表达,其中RcPIN5基因的表达水平较低。
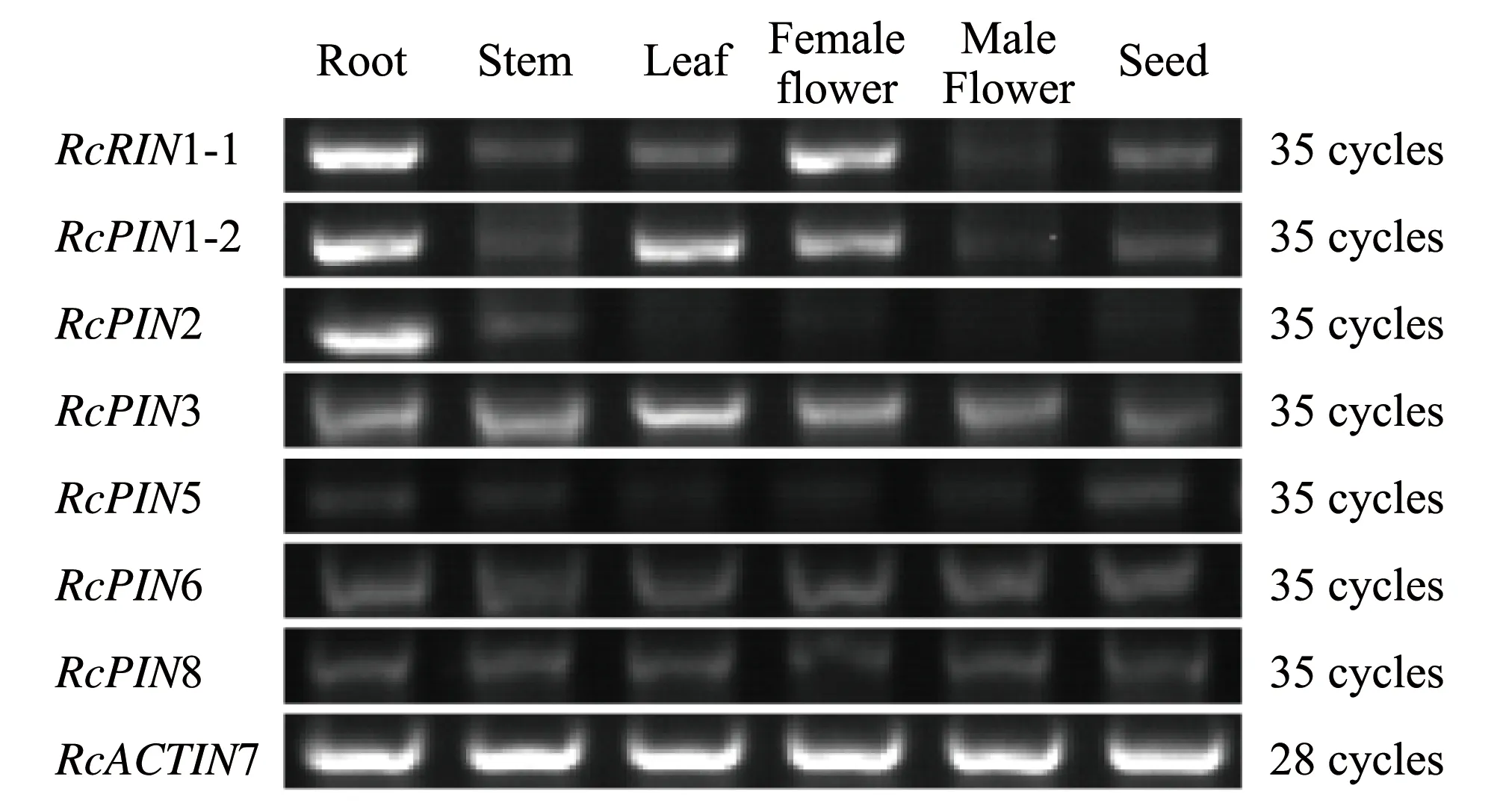
图7 蓖麻PIN基因家族在不同组织中的转录水平
3 讨论与分析
本研究共检索出7条蓖麻PIN基因序列,根据其与拟南芥PIN基因序列相似程度分别命名RcPIN1-1、RcPIN1-2、RcPIN2、RcPIN3、RcPIN5、RcPIN6、RcPIN8。这些蛋白均呈碱性,序列长度为356-646个氨基酸残基,分子量大小为38.89-70.39 ku。其中RcPIN5、RcPIN6和RcPIN8的氨基酸长度小于RcPIN1-1、RcPIN1-2、RcPIN3,符合上述亚族分类规律。这7种RcPIN蛋白均为稳定蛋白,但RcPIN2和RcPIN3不稳定系数接近40,无法确切判断。除RcPIN5和RcPIN8蛋白为疏水性蛋白外,其余RcPIN蛋白均为两性蛋白。另外,RcPIN5、RcPIN6和RcPIN8有相比于其他蛋白更高的脂溶指数。研究认为,脂溶指数更高说明PIN5、PIN6和PIN8所在亚族的蛋白质热稳定性更高[8]。推测这两个特性与其亲水环长度较小有关。信号肽预测结果显示,蓖麻PIN蛋白与拟南芥PIN蛋白一样都为非分泌性蛋白。从二级结构和跨膜结构域分析来看,与其他蛋白相比,RcPIN5和RcPIN8的二级结构的构成不同,且跨膜结构域的亲水环结构较不明显,由于其氨基酸长度较短,这可能导致RcPIN5和RcPIN8跨膜结构域模型与其他成员有差别。序列比对结果表示,PIN蛋白序列两端的序列较为保守,而中间的亲水环区域则差异较大。据此推测,其不同的生物学功能可能与其亲水环的差异有关。这些结果进一步从生物信息学的角度证明了拟南芥典型PIN的中心胞质亲水环(HL结构域)是导致它们功能差异的原因之一[7,39]。系统进化分析显示,蓖麻和与其同属大戟科的木薯、橡胶树和麻风树进化关系较为密切,与双子叶模式植物拟南芥和烟草的进化关系也较为亲密,可以参照这些植物PIN蛋白已知的生物学功能对蓖麻PIN蛋白进行下一步的研究。
我们注意到,蓖麻中似乎缺少与AtPIN4以及AtPIN7类似的蛋白。AtPIN3、AtPIN4和AtPIN7蛋白属同一亚族,在系统进化树中可以看到,AtPIN3、AtPIN4和AtPIN7蛋白聚成一个枝,进化关系十分密切。与蓖麻同属大戟科的植物同样缺少PIN3、PIN4和PIN7中的一个或两个,这些植物的生物信息学研究亦出现类似的结果。研究表明,AtPIN3、AtPIN4和AtPIN7的进化关系亲密且生物学功能较为冗余,原因是拟南芥中的AtPIN1、AtPIN3、AtPIN4和AtPIN7基因来源于进化中最近发生的一次染色体重复[7,39,40]。
蓖麻具有适应能力强、含油量高和经济效益高等特点,获取的蓖麻油粘度较高,摩擦系数较低,适合作为工业用油。获得产油率高、生长周期短且可用于大规模种植的优良转基因植株是蓖麻育种的首要目标,对蓖麻的市场地位与国家的新能源战略布局均有较大提升。生长素作为植物中重要的内源性激素,对植株的生长发育影响较大。生长素相关基因对蓖麻生长发育及抗逆性的研究,可为蓖麻育种提供有益的线索。PIN蛋白是生长素极性运输的主要承担者,目前对其在拟南芥等植物生长发育过程中功能的研究已有许多进展。若能以此为依据,探明蓖麻中PIN蛋白的生物学功能,则可以为其生长素极性运输机制的研究奠定基础,为选育蓖麻新品种和蓖麻种植行业的发展提供理论依据。
猜你喜欢
橡塑技术与装备(2022年6期)2022-06-02
中国蔬菜(2021年3期)2021-12-11
源流(2021年5期)2021-07-30
小天使·六年级语数英综合(2020年3期)2020-12-15
老友(2020年11期)2020-11-28
三农资讯半月报(2020年14期)2020-08-09
中国诗歌(2018年3期)2018-11-14
科技创新与应用(2017年12期)2017-05-08
创新作文(3-4年级)(2015年8期)2015-09-15
农家科技(2011年3期)2011-06-13
