
根系分隔方式下紫花苜蓿/燕麦间作氮素利用及种间互馈特征研究
2021-08-21 02:32汪雪刘晓静赵雅姣王静
草业学报 2021年8期
汪雪,刘晓静,赵雅姣,王静
(甘肃农业大学草业学院,草业生态系统教育部重点实验室,甘肃省草业工程实验室,中-美草地畜牧业可持续发展研究中心,甘肃兰州730070)
间作是通过不同作物在生长过程中对资源的利用差异来进行集约化种植,可实现光、肥、水、气、热等有限农业资源的高效利用[1],抑制杂草,降低病虫害[2],防止水土流失[3]等,进而提高单位土地面积的生产力,达到增产稳产,是我国传统农业中应用较为广泛的种植技术[4]。豆/禾间作在众多间作组合中具有显著优势,其很重要的一方面体现在豆/禾间作体系中存在种间氮营养竞争及互馈机制[5],种间氮素的互补利用会促进其间作体系内氮素的高效利用[6-7],而施氮水平是调控种间关系的常见措施之一[7]。植物生命活动所需的绝大部分氮素来源于其根系对土壤氮素的吸收,因此,提高根系吸收利用氮素的能力对于提高作物产量和氮素利用效率具有重要意义。在豆/禾间作群体内,不同作物调控氮素利用的生理和生态学过程,很大程度上依赖于种间根系互作[8],目前,关于根系互作对间作下氮素吸收利用影响的研究已在紫花苜蓿(Medicago sativa)/玉米(Zea mays)[9]、蚕豆(V icia faba)/玉米[10]等间作组合中进行,并认为豆/禾间作时,根系通过互利竞争可增强豆科作物的固氮能力,提高群体的氮素资源利用及减少氮肥的投入等[11]。利用根系分隔法[12-13]可真实有效地模拟间作中根系互作强度,更好地阐释种间氮营养互补及利用机制。目前此方法已被应用在紫花苜蓿/玉米[9]、大豆(Glycine max)/玉米[14-16]、蚕豆/小麦(Triticum aestivum)[17]等间作体系中来研究作物的生长特性,氮、磷养分吸收利用特性和评价作物根区土壤残留养分差异[18]等,来不断揭示间套作模式下种间营养竞争与促进的互作机理。
紫花苜蓿在我国具有悠久的种植历史,是一种优质的植物性蛋白资源[19],具有丰富的维生素和矿物质,同时其结瘤固氮能力会减少化学氮肥的使用,促进氮素在农业系统中的利用效率[20]。燕麦(Avena sativa)属粮饲兼用型作物,是中国西北干旱地区重要的饲草作物。紫花苜蓿与燕麦间作不仅使碳、氮代谢更加活跃[21],还可促进紫花苜蓿对氮素的固定,进而提高间作群体的氮素利用效率[22],此外,它们皆具有重要的经济价值,其种植面积在我国呈逐年增长的态势[23]。目前,关于饲草间作组合的研究主要集中在紫花苜蓿/玉米[9]、燕麦/箭筈豌豆(Vicia sativa)[24]等组合,且相关研究方向主要集中在生产潜力评定[25]、土壤理化性质[26]等方面,而对紫花苜蓿/燕麦间作根系互作氮素吸收利用及种间互馈机制的研究很少。为此,本试验拟采用根系分隔法来模拟紫花苜蓿/燕麦间作下的不同根系互作关系,以研究豆/禾间套作模式下种间氮素的互补和促进作用,为减少氮肥施用量、优化间作系统的氮素利用及氮素互馈机制以及提高豆/禾间作资源利用效率提供理论依据。
1 材料与方法
1.1 供试材料
紫花苜蓿(LW6010):由北京猛犸种业有限公司提供;燕麦(海威):由克劳沃公司提供;中华根瘤菌(12531):由甘肃农业大学提供。
1.2 试验设计
于2017年3月5日在甘肃农业大学的防雨网棚内进行试验。采用桶栽土培法,即紫花苜蓿与燕麦种植于塑料桶中(直径为47 cm,高为40 cm),试验中用的土样来自甘肃农业大学牧草试验基地,土壤pH 8.23,有机质(organic matter)12.23 g·kg-1,全氮(total nitrogen)0.81 g·kg-1,碱解氮(available nitrogen)81.24 mg·kg-1,速效磷(available phosphorous)15.22 mg·kg-1,速效钾(available potassium)118.12 mg·kg-1。
采用3因素设计试验,A因素为2个氮素水平,即:1)不施氮水平(N21);2)施氮水平(N210)[0.15 g·kg-1尿素,CO(NH2)2(N≥46%),该氮素是在装桶种植前一次性加入土壤中];B因素为5种种植方式,即:1)紫花苜蓿单作(SA);2)燕麦单作(SO);3)紫花苜蓿与燕麦间作根系不分隔,简称不分隔(A-O);4)紫花苜蓿与燕麦间作根系尼龙网分隔,简称尼龙网分隔(NA-O);5)紫花苜蓿与燕麦间作根系塑料分隔,简称塑料分隔(PA-O);C因素为不同的取样时期,即:1)紫花苜蓿分枝期(Ⅰ);2)紫花苜蓿现蕾期(Ⅱ);3)紫花苜蓿初花期(Ⅲ)。每个处理重复3次,其中根系的分隔方式参考李隆等[27]的方法。先将紫花苜蓿种植于桶中,单作每桶留20株,根系分隔则种植于被分隔后桶的一侧,每桶留10株。紫花苜蓿出苗后,对燕麦进行2~3 d的催苗,随后将露白的燕麦种子均匀移入相对应的塑料桶内,单作每桶留20株,根系分隔则留10株。在紫花苜蓿出苗1周后,接种根瘤菌25 mL·桶-1。待燕麦出苗前每天3次用喷壶进行喷水,保持土壤的湿润;待燕麦出苗后,每3 d浇1次水,保持紫花苜蓿和燕麦的正常生长。分别于2017年6月15日(分枝期)、7月1日(现蕾期)、7月16日(初花期)取样,将待测样品(带桶)运至实验室,直接从桶栽植株上进行茎叶的取样,后将根系用蒸馏水冲洗干净,并用吸水纸吸干后迅速取样并测定。
1.3 测定指标
取样后于105℃烘箱中杀青15 min,再调至85℃烘干36 h,称其干重。采用凯氏定氮法测定植株氮含量并计算[28]。氮积累量为干物质重与氮含量的乘积。采用邹琦[29]的方法测定硝酸还原酶(nitrate reductase,NR)活性,单位:μg·g-1FW·h-1以及谷氨酰胺合成酶(glutamine synthetase,GS)活性,单位:μmol·g-1FW·h-1。利用Epson根系扫描仪对活根进行扫描并存入电脑,再用WinRHIZO根系分析系统软件获得根系总长度、根表面积、根系平均直径和根体积数据,每个处理重复3次。根部用蒸馏水清洗干净后,用吸水纸吸干,然后立刻取出结节。每株紫花苜蓿的根瘤数量为单株根瘤数、重量为单株根瘤重;每株紫花苜蓿粉色及浅粉色根瘤(主要是豆血红蛋白的颜色)数量为单株有效根瘤数。每株紫花苜蓿的有效/根瘤数为单株有效根瘤数与单株根瘤数的比[9]。采用TTC法测定根系活力[28]。
1.4 数据分析
采用Excel 2016软件整理试验数据,用SPSS 22.0软件进行单因素方差分析。
2 结果与分析
2.1 紫花苜蓿/燕麦间作下的氮素吸收
2.1.1 干物质重 紫花苜蓿地上干物质重在分枝期时,各分隔方式之间差异不显著,在现蕾期和初花期时,表现为紫花苜蓿单作及塑料分隔均显著大于不分隔(图1);燕麦地上干物质重在分枝期时,各分隔方式之间差异不显著,在现蕾期和初花期时,表现为不分隔显著大于燕麦单作(P<0.05)。N210下,紫花苜蓿地下干物质重表现为紫花苜蓿单作和塑料分隔均显著大于不分隔,N21下,紫花苜蓿地下干物质重在分枝期和初花期时,均表现为紫花苜蓿单作显著大于其他处理;燕麦地下干物质重除N210现蕾期外,均表现为不分隔显著大于塑料分隔和尼龙网分隔(P<0.05)。
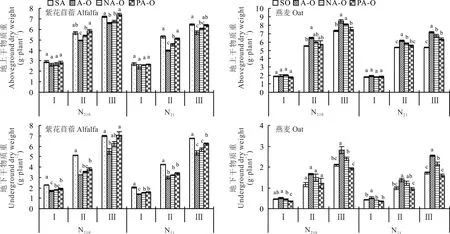
图1 不同分隔方式及氮素水平对紫花苜蓿和燕麦干物质重的影响Fig.1 Effect of differ ent r oot partitions and N level on dr y matter of alfalfa and oat
2.1.2 氮含量 N210下,紫花苜蓿地上氮含量在分枝期时,表现为塑料分隔显著大于紫花苜蓿单作,在现蕾期和初花期时表现为紫花苜蓿单作和塑料分隔均大于不分隔,N21下,除分枝期外,均表现为紫花苜蓿单作显著大于不分隔(图2);燕麦地上氮含量在每个处理下,均表现为不分隔显著大于燕麦单作(P<0.05)。紫花苜蓿地下氮含量在现蕾期和初花期时,表现为紫花苜蓿单作及塑料分隔均显著大于不分隔;燕麦地下氮含量在N21现蕾期和初花期时,表现为不分隔及尼龙网分隔均显著大于燕麦单作和塑料分隔(P<0.05)。
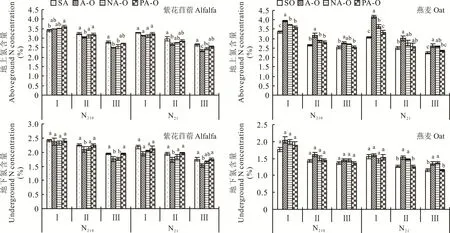
图2 不同分隔方式及氮素水平对紫花苜蓿和燕麦氮含量的影响Fig.2 Effect of different root partitions and N level on N concentration of alfalfa and oat
2.1.3 氮积累量 紫花苜蓿地上氮积累量在现蕾期和初花期时,表现为紫花苜蓿单作及塑料分隔均显著大于不分隔;燕麦地上氮积累量均表现为不分隔显著大于燕麦单作和塑料分隔(P<0.05)。N210下,紫花苜蓿地下氮积累量在现蕾期和初花期时,表现为紫花苜蓿单作显著大于不分隔,在N21下,均表现为紫花苜蓿单作显著大于不分隔;燕麦地下氮积累量除N21分枝期外,均表现为不分隔显著大于燕麦单作(P<0.05)(表1)。
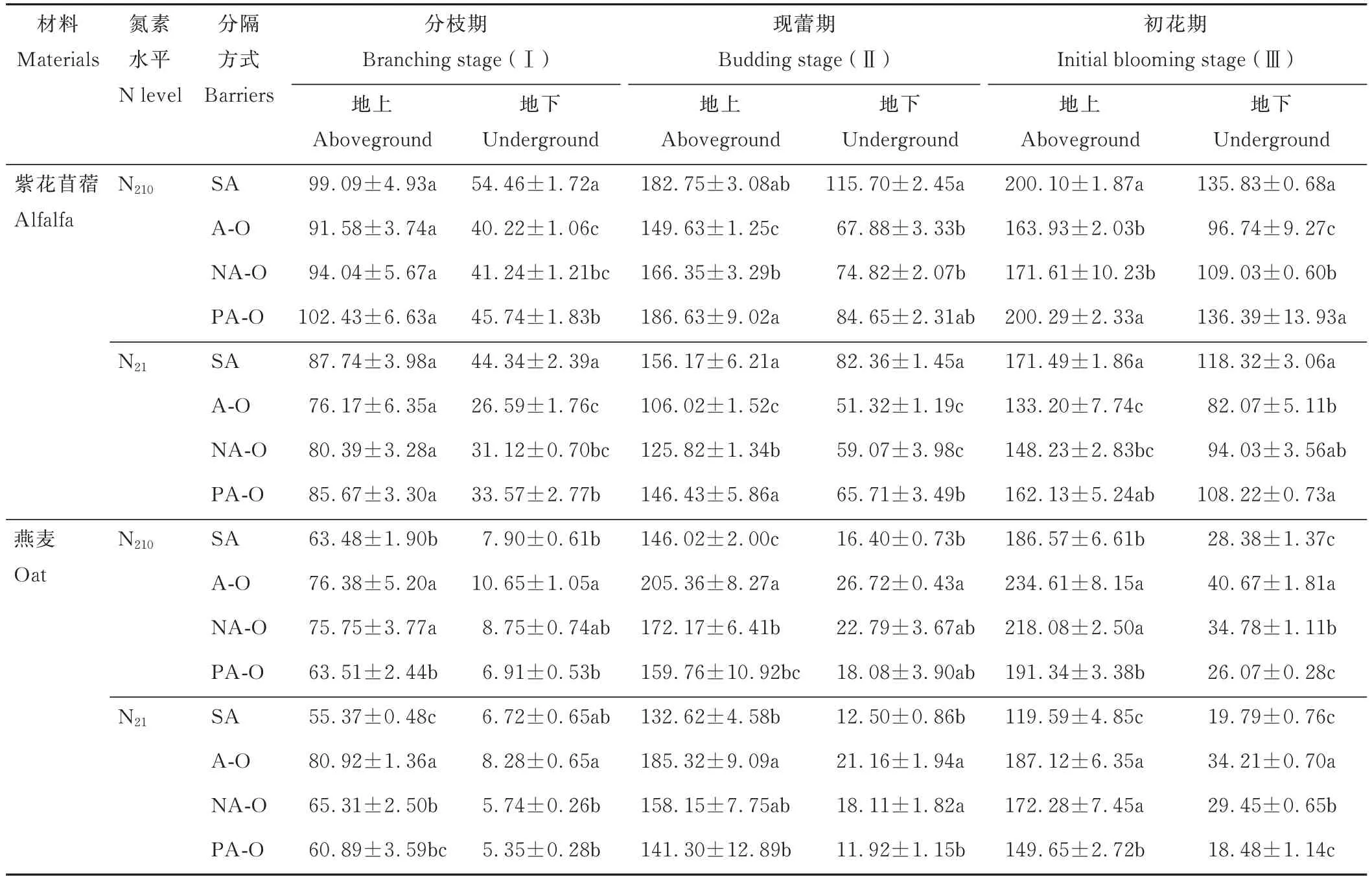
表1 不同分隔方式及氮素水平对紫花苜蓿和燕麦氮积累量的影响Table 1 Effect of different root partitions and N level on N accumulating of alfalfa and oat(mg·plant-1)
2.2 紫花苜蓿/燕麦间作下的氮代谢关键酶活性
2.2.1 硝酸还原酶(NR) 紫花苜蓿地上NR活性在每个处理下,均表现为紫花苜蓿单作显著大于尼龙网分隔(图3);燕麦地上NR活性在分枝期和初花期时,均表现为不分隔显著大于塑料分隔和燕麦单作,在现蕾期时,表现为不分隔及尼龙网分隔均显著大于燕麦单作和塑料分隔(P<0.05)。紫花苜蓿地下NR活性在每个处理下,均表现为紫花苜蓿单作显著大于不分隔和尼龙网分隔;燕麦地下NR活性在每个处理下,均表现为不分隔显著大于尼龙网分隔和燕麦单作(P<0.05)。
2.2.2 谷氨酰胺合成酶(GS) N210下,紫花苜蓿地上GS活性在分枝期时,表现为紫花苜蓿单作及尼龙网分隔显著大于塑料分隔,在现蕾期和初花期时,表现为紫花苜蓿单作显著大于不分隔,N21下,紫花苜蓿地上GS活性在分枝期时,表现为尼龙网分隔显著大于塑料分隔,现蕾期和初花期时,表现为紫花苜蓿单作显著大于塑料分隔(图4);燕麦地上GS活性除N210分枝期外,均表现为不分隔显著大于燕麦单作(P<0.05)。紫花苜蓿地下GS活性除N210分枝期外,均表现为紫花苜蓿单作显著大于尼龙网分隔;燕麦地下GS活性在初花期时,表现为不分隔显著大于燕麦单作和尼龙网分隔(P<0.05)。
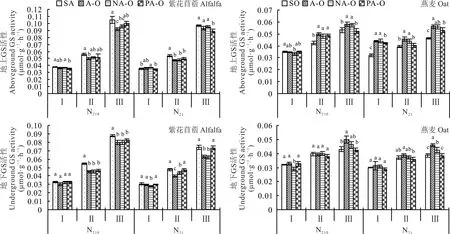
图4 不同分隔方式及氮素水平对紫花苜蓿和燕麦GS活性的影响Fig.4 Effect of differ ent r oot partitions and N level on GSactivity of alfalfa and oat
2.3 紫花苜蓿/燕麦间作下的根系特性
2.3.1 根系形态 紫花苜蓿总根长均表现为不分隔显著大于尼龙网分隔和塑料分隔;燕麦总根长除N21分枝期外,均表现为不分隔显著大于塑料分隔(P<0.05),除N21的现蕾期外,燕麦单作与尼龙网分隔之间无显著差异。紫花苜蓿根表面积在N210现蕾期和初花期时,表现为紫花苜蓿单作显著大于不分隔和尼龙网分隔,在N21分枝期和现蕾期时,表现为紫花苜蓿单作显著大于其余分隔方式;燕麦根表面积均表现为不分隔显著大于塑料分隔(P<0.05)。紫花苜蓿根平均直径在N210下,均表现为紫花苜蓿单作及塑料分隔都显著大于不分隔和尼龙网分隔,在N21下,均表现为紫花苜蓿单作显著大于不分隔;燕麦根平均直径均表现为不分隔显著大于塑料分隔(P<0.05)。紫花苜蓿根体积均表现为紫花苜蓿单作显著大于不分隔;燕麦根体积除N21分枝期外,均表现为不分隔显著大于尼龙网分隔和塑料分隔(P<0.05)(表2)。
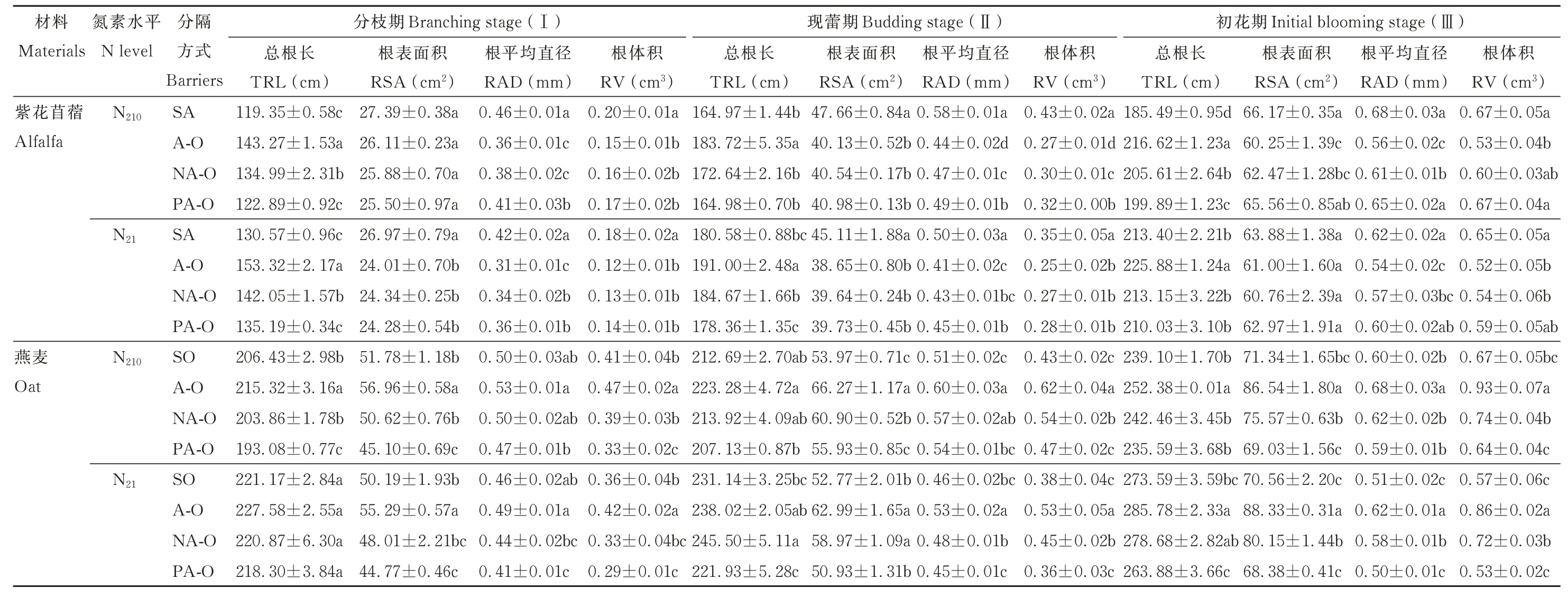
表2 不同分隔方式及氮素水平对紫花苜蓿和燕麦根系形态的影响Table 2 Effect of different root partitions and N level on root morphology of alfalfa and oat
2.3.2 根系活力 N210下,紫花苜蓿根系活力在现蕾期和初花期时,表现为不分隔显著大于尼龙网分隔和塑料分隔,N21下,紫花苜蓿根系活力在分枝期和初花期时,表现为不分隔显著大于尼龙网分隔,紫花苜蓿单作和塑料分隔(图5A);燕麦除N21分枝期外,均表现为不分隔显著大于尼龙网分隔,塑料分隔和燕麦单作(P<0.05)(图5B)。
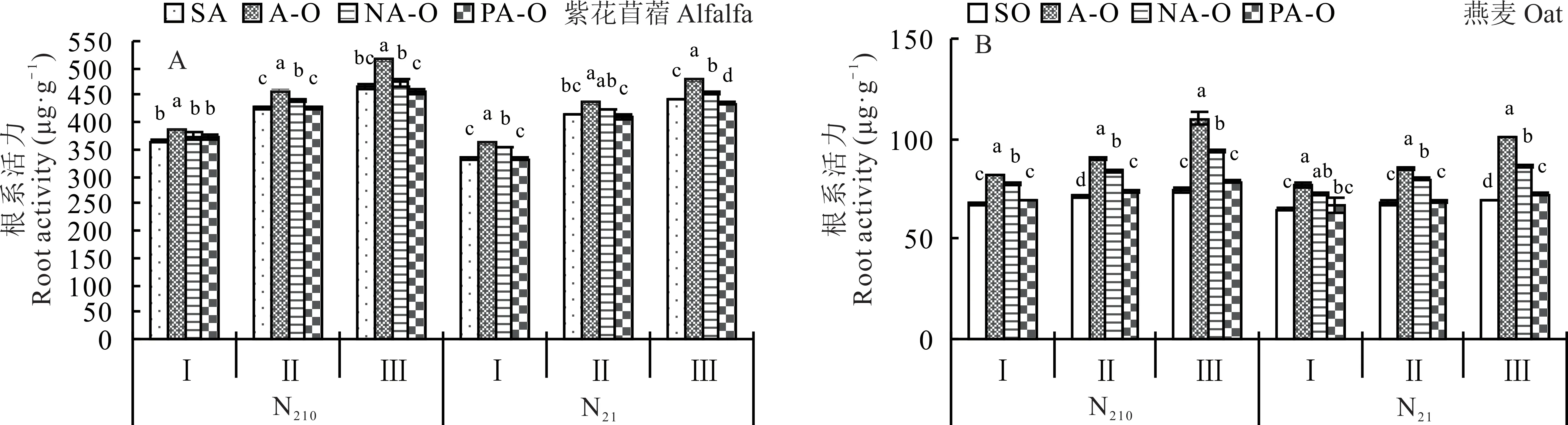
图5 不同分隔方式及氮素水平对紫花苜蓿和燕麦根系活力的影响Fig.5 Effect of different root partitions and N level on root activity of alfalfa and oat
2.4 紫花苜蓿结瘤特性
N210下,紫花苜蓿总根瘤数在各时期内,均表现为不分隔显著大于单作,尼龙网分隔与塑料分隔之间差异不显著;N21下,除分枝期外,均表现为不分隔显著大于单作及塑料分隔(P<0.05)。单株有效根瘤数均表现为不分隔显著大于单作及塑料分隔(P<0.05)。有效/总根瘤数、总根瘤重在N210下均表现为不分隔显著大于尼龙网分隔,单作及塑料分隔;在N21下,均表现为不分隔显著大于单作及塑料分隔(P<0.05)(表3)。
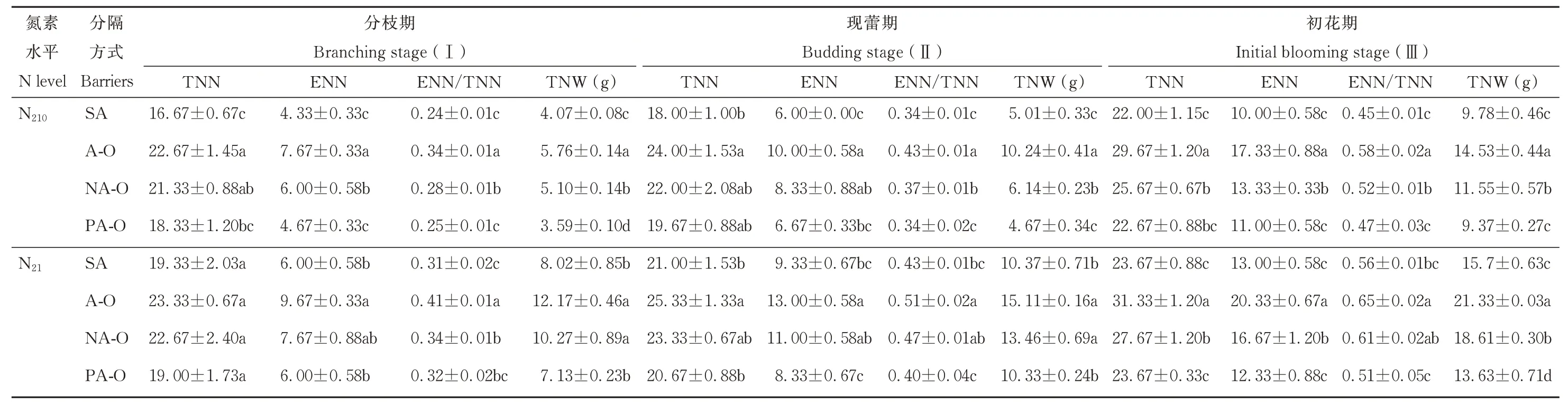
表3 不同分隔方式及氮素水平对紫花苜蓿结瘤特性的影响Table 3 Effect of different root partitions and N level on nodule
3 讨论
氮素代谢受NR和GS等活性的调节[30],NR作为氮代谢途径中的关键酶与限速酶,是氮同化的最初反应催化者[31],可调节NO3--N的还原,从而调节氮代谢,GS主要参与NH4+-N同化过程,可通过GS/GOGAT循环转化利用95%左右铵态氮[32]。本研究发现,NR和GS活性在不同根系互作强度(3种分隔方式)下表现不同,紫花苜蓿的NR和GS活性均表现为单作>塑料分隔>尼龙网分隔>不分隔,而燕麦相反,这说明根系互作越紧密越有利于燕麦NR和GS活性的提 高。在塑料分隔时,根系和养分被隔开,地下无任何互作,种间氮素竞争最弱;尼龙网分隔时,根系具有半交互作用,根系之间无交互影响,但水分和养分可以交互流动,菌根可以通过,菌根菌丝能够间接帮助植物根系吸收其所形成的营养匮乏区,间接提高植物的氮素利用率,燕麦具有较强的养分竞争能力,故燕麦能获得来自紫花苜蓿周围少量的氮素,这与张宇亭等[33]在玉米/油菜(Brassica napus)间作体系中的研究结果一致;不分隔时,燕麦与紫花苜蓿根系交互重叠,菌根侵染率升高,从而加强了燕麦从紫花苜蓿一侧竞争性地吸收氮素,使得紫花苜蓿根系环境中的氮素浓度降低,肖焱波等[34]的研究中也有此发现,这是由于紫花苜蓿根系环境中氮素的降低刺激并加强了紫花苜蓿结瘤固氮和向燕麦转移,使得燕麦NR和GS活性均有显著的提高,且两者变化趋于一致,唐秀梅等[35]在木薯(Manihot esculenta)/花生(Arachis hypogaea)间作中发现,间作花生的NR和GS活性均低于单作,郭峰等[36]与宋艳霞等[37]在麦套花生及套作大豆的研究中也有类似发现。此外,吴雅薇等[38]和赵吉平等[39]在小麦和玉米的研究中发现,氮代谢酶活性与氮素浓度和生育期呈正相关,并影响氮代谢积累和生物量,在本研究中,不分隔时的燕麦和单作时的紫花苜蓿干物质重及氮积累量均高于其余分隔方式,且随着环境氮水平的提高和生育期的推进而提高,这是由于不分隔时,根系互作紧密,提高了燕麦的氮代谢酶活性,促进其对氮素的积累和利用,进而提高其生物量,且随着适宜氮素的添加和生育期的推进,燕麦的氮代谢水平不断增强,而紫花苜蓿无论在何种分隔方式下都处于劣势地位。也有研究证明[40],作物产量的上升完全来源于地下种间的互惠作用,这种互惠作用主要来源于根系分泌物所产生的非直接接触,当紫花苜蓿和燕麦间作时,两者的根系分泌物会发生化感促进作用,提高土壤养分的有效性,又因燕麦是优势主导作物,进而在不分隔时表现出较高的生物量。可见,根系互作和氮水平对NR和GS活性的影响程度较大,不分隔和施氮促进了燕麦对土壤氮素的吸收,提高了燕麦氮代谢酶活性和氮代谢水平,加速了蛋白质的合成和氮积累量,提高其生物量,且随着生育期的推进,影响不断增大。综上,间作下紫花苜蓿和燕麦的根系互作关系越紧密,氮素浓度越高,越有利于燕麦对土壤氮素的竞争和吸收,进而可降低紫花苜蓿根区有效氮含量,刺激紫花苜蓿提高结瘤固氮能力,促进间作中氮的互补利用,提高燕麦的氮代谢水平和氮代谢酶活性,增加燕麦氮代谢产物积累,提高其生物量。
不同分隔方式下根系的空间生态位体现了根系对水分和养分吸收的有效空间[10]。根系形态的差异和根系活力的作用是造成间作作物养分吸收不同的主要原因[41]。本研究发现,在不同根系分隔方式下,紫花苜蓿根表面积、根平均直径和根体积均表现为单作>塑料分隔>尼龙网分隔>不分隔,而燕麦相反,这是由于紫花苜蓿和燕麦在不分隔时地下互作最强,燕麦会侵占紫花苜蓿的根系空间并具有较高的养分竞争优势,可竞争到紫花苜蓿根系周围更多的氮素,从而改善其根系特性,而处于空间分布和氮素竞争劣势的紫花苜蓿根体积和根平均直径会减小,张义凯等[42]和李玉英等[10,43]在蚕豆/玉米体系的研究中也发现了相同的规律,蚕豆与玉米种间互作扩展了根系空间生态位,增加了两者在横向和纵向两个尺度上的根长密度、根表面积、根系体积,且根长密度和根表面积与两种作物氮素吸收呈正相关。本研究中,紫花苜蓿和燕麦的根系活力在不分隔时明显提高,这是由于间作提高了作物的根系活力,促进其释放根系分泌物,影响根际微生物的群落分布和生长,活化了根际土壤中难溶性的磷,促进根际土壤磷的有效性[44],加强作物对土壤氮、磷营养元素的吸收,这也说明,氮素吸收的能力还与根系活力呈正相关[45-46],可见,根系互作关系越紧密,越有利于紫花苜蓿和燕麦根系活力的提高,促进其对氮素的吸收。此外,随着环境氮水平的提高和生育期的推进,不同分隔方式之间的差距变得明显,紫花苜蓿与燕麦的根系形态、根系活力均随之改善,从而增强种间氮营养竞争互补利用机制,本团队的前期研究中也发现,间作中种间强烈的氮素竞争会刺激紫花苜蓿增加其根瘤皮层内O2的扩散,促使根系分泌物中类黄酮物质的组分和含量发生变化,最终促进根瘤的生长发育,提高其生物固氮能
力来补充自身所缺的氮素营养[9]。另外,在本研究中,紫花苜蓿的总根瘤数、有效根瘤数、有效/总根瘤数、总根瘤重均在不分隔时显著大于其余分隔方式,可见根系互作关系越紧密,越有利于紫花苜蓿增加根瘤的数目和有效性,提高其结瘤固氮能力。再者,在本研究中,紫花苜蓿在各时期的同一分隔方式下结瘤特性均表现为施氮小于不施氮,这充分证明了根系互作中环境氮素浓度的降低有利于豆科作物结瘤与固氮,这一研究结果与王立波等[47]的研究结果相同,但与赵雅姣等[9]在紫花苜蓿/玉米间作中发现有效根瘤数、有效/总根瘤数、氮根瘤重在分枝期的各分隔方式下,其随氮素浓度的增加而变大的结果不同,造成此结果的原因可能是作物组合及种植密度的不同造成了对养分竞争的不同。本研究中还表明,随着生育期的推进,紫花苜蓿结瘤特性在2个氮水平下均表现为各分隔方式之间差距不断增大,这种表现在初花期最明显,其趋势仍表现为不分隔大于尼龙网分隔和塑料分隔,可见,无论何种氮素水平和生育时期,根系互作程度是影响紫花苜蓿/燕麦间作中紫花苜蓿结瘤固氮的主要因素,且根系互作关系越紧密、环境氮素越低越有利于紫花苜蓿结瘤固氮。综上所述,间作下紫花苜蓿/燕麦的根系相互作用会随着共生期逐渐增强,且根系互作关系越紧密,越有利于燕麦根系空间生态位的扩展,增强其根系活力以竞争到更多氮素,土壤氮素的降低促进紫花苜蓿结瘤固氮,并将部分氮素转移给燕麦,可提高间作群体氮营养的互补利用。
4 结论
在紫花苜蓿/燕麦间作体系中,根系互作越紧密,越有利于燕麦根系的生长和根系活力的提高,增强燕麦氮代谢酶活性和氮代谢产物积累,提高燕麦氮素吸收和氮代谢能力,促进紫花苜蓿结瘤固氮并将由此产生的部分氮素继续转移给燕麦,增强间作系统中种间氮营养的互补利用,以达到紫花苜蓿/燕麦间作体系内的种间氮营 养高效回馈,从而有效提高了间作体系中氮素的利用效率;且适宜氮素的添加和生育期的推进,使得种间根系互作不断加强,加强了种间氮素的互补利用,促进其间作体系内氮素的高效利用。
综上所述,紫花苜蓿/燕麦间作体系中,燕麦和紫花苜蓿根系互作越紧密,其种间氮营养互馈就越活跃,间作体系的氮素利用就越高效,因此,紫花苜蓿/燕麦间作是一种氮素高效利用的牧草种植方式,当其投入生产时,适当添加氮素,并在两种作物进入生殖生长前进行收割可获得较高的生产效益。
猜你喜欢
中国农学通报(2022年28期)2022-11-23
商洛学院学报(2022年2期)2022-05-11
广西林业科学(2022年1期)2022-03-03
大众标准化(2022年17期)2022-02-06
中国南方果树(2022年1期)2022-01-28
农技服务(2021年7期)2021-09-24
中国饲料(2020年21期)2020-12-23
防护林科技(2020年6期)2020-08-12
河北果树(2020年1期)2020-02-09
植物营养与肥料学报(2019年11期)2019-12-13
