
波密岗乡华山松林优势植物空间格局及其关联性
2021-08-16 01:46姚慧芳屈兴乐
西北林学院学报 2021年4期
姚慧芳,卢 杰*,王 超,陈 康,屈兴乐,张 铎
(1.西藏农牧学院 高原生态研究所,西藏 林芝 860000;2.西藏高原森林生态教育部重点实验室,西藏 林芝 860000;3.西藏林芝高山森林生态系统国家野外科学观测研究站,西藏 林芝 860000)
种群的空间分布格局是指种群中个体在水平空间中的分布状况[1-2],是种群生物学特性、种内、种间联结以及与外部环境综合作用的结果[3],这不仅能够反映出种群在适应某个生境后在空间内的分布格局,还能反映出该种群与其他种群之间的关系属性[4]。研究植物的空间分布格局有助于了解种群特性及发展趋势,进一步揭示出植物群落的特征以及分布状况,对于制定相关的保护措施提供一定的科学依据[5-6]。关联性分为种内关联性和种间关联性,种内关联性是指在一个种群中,不同大小个体之间的关联性,种间关联性是指不同物种的空间分布关系,是植物群落的重要特征之一。研究种内和种间联系,有助于深入了解植物种群和群落的内部特征以及不同物种之间的关系。
华山松属松科 (Pinaceae) 松属 (Pinus) 的常绿乔木,对于我国西部一些比较干燥降雨量少的地区而言是十分重要的树种,并且在全国分布也较为广泛,在山西、陕西、云南、西藏等13个省份海拔1 000~3 300 m均有分布,生长在平均气温18℃以下、年降水量600~2 000 mm的地区,在西藏主要分布在雅鲁藏布江下游,其生长迅速、耐寒力强,能适应多种土壤,且其木材性质优良,种子可供食用,经济效益很高,具有较强的水源涵养及水土保持生态功能[7]。目前关于华山松的研究主要集中在群落特征[8-9]、种群动态[10]、幼苗更新特征[11]等方面,研究区域也主要以秦岭为主,对于西藏华山松的研究鲜有报道。
因此,以藏东南波密县岗乡的华山松林作为研究对象,通过点格局研究方法探究华山松林优势植物的空间分布格局,并且对其关联性进行分析,以便了解本地区华山松林优势植物的分布形态及其与环境之间的关系,对华山松的后续研究提供借鉴和参考。
1 研究区概况
研究地位于西藏东南部的林芝市波密县岗乡自然保护区内,该区位于波密县以西22 km,帕隆藏布江以南,雅鲁藏布江大拐弯的东北部(29°53′8.379″N,95°37′22.626″E),区内大部分山区海拔约2 600~5 000 m,根据中国气候带的划分,保护区属于高原高山寒温带气候,但由于可以受到从印度洋而来西南季风的影响,因此气候较为湿润,年平均温度约8.3℃,年平均降水量可达1 000 mm、相对湿度也较高约70%~80%。该区占地面积近4 600 hm2,其中森林面积占1/2以上约2 800 hm2,森林覆盖率超过61%,森林内总蓄积量约2.52×106m3,林区植被茂密,并且以华山松和林芝云杉等针叶林为主,区内森林的生产力较高达2400 m3/hm2。研究地乔木层植被除华山松外,还有川滇高山栎(Quercusaquifolioides)、槭属(Acer)等,灌木层主要有忍冬(Lonicerajaponica)、三桠乌药(Linderaobtusiloba)、卫矛(Euonymusalatus)、锦鸡儿(Caraganasinica)、牛奶子(Elaeagnusumbellata)、鲜黄小檗(Berberisdiaphana)、白簕(Acanthopanaxtrifoliatus)、常春藤(Hederanepalensisvar.sinensis)、黑茶藨子(Ribesnigrum)等,草本主要有天南星(Arisaemaheterophyllum)、唐松草(Thalictrumaquilegifoliumvar.sibiricum)、狭叶兔耳风(Ainsliaeaangustifolia)、草玉梅(Anemonerivularis)、蕨(Pteridiumaquilinumvar.latiusculum)等。
2 材料与方法
2.1 样地设置与调查
2020年6月下旬在西藏林芝市波密县岗乡进行实地调查,设置1块具有典型性的100 m×100 m华山松林样地,按相邻格子法将样地划分成100个10 m×10 m 的单元格,对样地的基本情况进行调查并记录:1)样地海拔为2 716 m,坡度为15°,郁闭度达0.86,盖度为90%;2)对样地内所有乔木进行每木检尺,并以样地的西南角作为原点,测定并记录每株乔木的空间位置,样地内是以华山松为优势树种的混交林,其他树种还有高山松、林芝云杉和川滇高山栎等,树种组成比例依次是华山松占62.64%,高山松占21.39%,林芝云杉占8.09%,川滇高山栎占2.97%,其他占4.91%。3)样地内对角线上设置9个5 m×5 m的小样方,调查样方内华山松的幼树和灌木的位置及种类;4)样地内对角线上设置17块2 m×2 m的小样方,调查样方内草本的种类。
2.2 林木分级方法
根据研究目的,并结合本研究的具体情况,记录完数据后筛选出树高(H)≥1 m的乔木,参考石蒙蒙等[12]和许爱云等[13]对龄级的划分标准将筛选出来的华山松根据胸径(D)划分为3个龄级:Ⅰ级(D<20 cm且H≥100 cm)、Ⅱ级(20 cm≤D<40 cm)、Ⅲ级(D≥40 cm),其中,Ⅰ级为小树,Ⅱ级为中树,Ⅲ级为大树。
2.3 研究方法
2.3.1 点格局分析 点格局分析的相关理论最早是Ripley(1977)提出来的,Diggle等进一步发展完善。这种方法可以清晰直观地分析在各种尺度下的种群的分布类型,因此被生态学家广泛应用[14]。本次研究所采用的是点格局分析方法中的RipleyL函数来分析探索植物的空间分布格局及其关联性。此函数以植物的空间位置信息作为基础来分析空间分布格局,L函数为K函数的改进,首先列出K函数公式如下:
(1)
式中,A表示样地的面积,n为样方中物种的数量,uij表示i、j2棵植株之间的距离,r表示空间尺度,当uij≤r时,Ir(uij)=1;当uij>r时,Ir(uij)=0。上式中Wij表示权重,用于边缘校正。L函数公式如下:
(2)
式中,若L(r)=0则该种群为随机分布;L(r)>0,则种群为集群分布;L(r)<0则种群为均匀分布。
为了更加准确地分析空间分布格局的类型,本研究将采用Monte-Carlo随机模拟,会得到2条包迹线围成的置信区间,位于置信区间上方的属于集群分布;位于置信区间内的属于随机分布;位于置信区间以下的属于均匀分布。
2.3.2 关联性分析 有关种内和种间空间关联性的点格局分析,也可以叫作多元点格局分析[15],将K函数和L函数应用于双变量中进行计算。具体公式如下:
(3)
式中,n1、n2分别代表2个种群或同一种群不同等级的个体数,其他指标与式(1)中的含义相同。L函数的双变量公式如下:
(4)
若L12(r)<0,表明在r尺度上2种群之间为负关联;若L12(r)=0则表明在r尺度上2个种群无关联;若L12(r)>0则表明在r尺度上2个种群的种间关系为正关联[16-17]。继续采用Monte-Carlo随机模拟方法拟合出置信区间,位于置信区间以上的是正相关;在置信区间内的是无相关;位于置信区间以下的属于负相关。
2.4 数据处理
采用Programmita(2008版)软件完成数据计算和分析,选取的空间尺度是0~50 m,通过100次Monte-Carlo随机模拟得到99%的置信区间[18]。利用Excel 2010进行绘图。
3 结果与分析
3.1 不同龄级华山松种群的空间分布格局
1 hm2样地内树高≥1 m的华山松共计有503株:其中,小树(D<20 cm且H≥100 cm)有364棵,中树(20 cm≤D<40 cm)有117棵,大树(D≥40 cm)仅有22棵(图1)。华山松的株数随着龄级的增大逐渐减少,小树和中树的个体数占总数的96%,大树仅占4%,这说明华山松种群属于增长型,结构良好,更新能力较强。
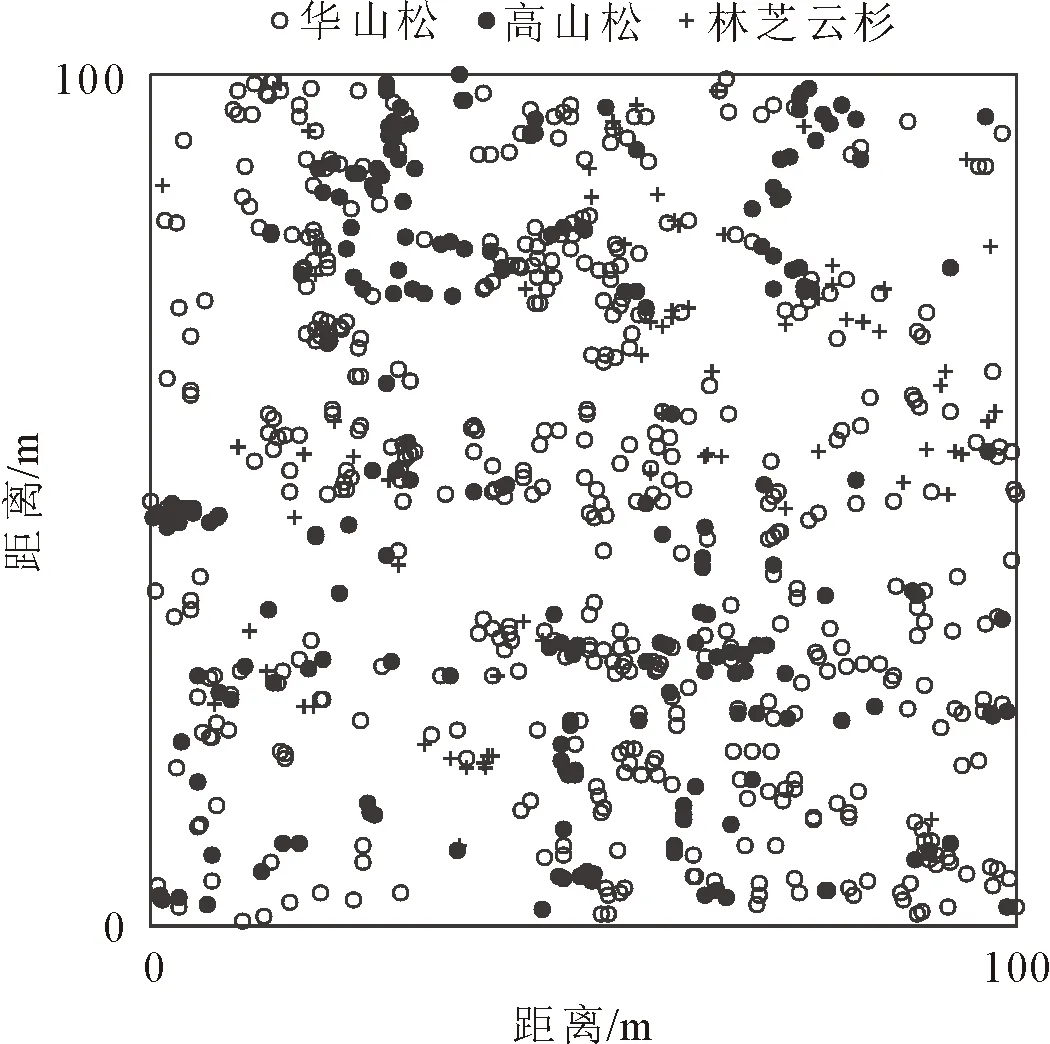
图1 3种优势植物在1 hm2样地内的分布点
从图2可以看出,小树的L(r)函数值在0~50 m的空间尺度内都位于置信区间上方,表现为集群分布,其中在空间尺度37 m时集聚度最高,L(r)值为1.759 4;中树的L(r)函数值在空间尺度0~3 m、11 m和16~50 m处位于置信区间内,表现为随机分布,在3~11 m和11~16 m的空间尺度内位于置信区间上方,呈集群分布,在空间尺度6 m时集聚度最高,L(r)值为1.642 1;大树的L(r)函数值在0~50 m的空间尺度内都位于置信区间内,表现为随机分布。

注:TS:实线代表确定空间分布格局的函数L(r)的值,上下2条虚线代表用Monte-Carlo随机模拟方法拟合后最大值、最小值形成的包迹线。
3.2 华山松林优势植物的空间分布格局
在所调查的华山松林样地中,优势植物有华山松、高山松和林芝云杉,华山松有612株,占所有乔木数量的62.64%;高山松有209株,占21.39%;林芝云杉有79株,占8.09%。通过图2可以发现,它们在样地内的分布情况不同,华山松在整个样地内都有分布,集聚程度也较高;高山松在样地的西北和东南方向分布较为集中,主要分布在坐标为0≤x≤50,50≤y≤100和50≤x≤100,0≤y≤50的范围内;林芝云杉由于个体数相对较少,在样地内的分布呈小区域集中。3种优势植物在样地内的分布状况不同,这可能是由于不同树种对于环境的响应不同而造成的。对这3个树种分别进行独立的空间分布格局分析(图3),华山松的L(r)函数值在0~50 m的空间尺度上均位于置信区间上方,呈集群分布,其中在空间尺度4 m和39 m处的集聚度最高,L(r)值分别为1.359 6和1.347 2;高山松的L(r)函数值在0~30 m的空间尺度内位于置信区间上方,表现为集群分布,在30~37 m及46~50 m的尺度上位于置信区间内,呈随机分布,在37~46 m的空间尺度内位于置信区间下方,表现成均匀分布;林芝云杉的L(r)函数值在0~49 m的空间尺度内位于置信区间上方,呈集群分布,其中在空间尺度27 m时集聚度最高,L(r)值为4.886,L(r)函数值在49~50 m空间尺度上位于置信区间内,表现为随机分布。

图3 华山松林优势植物空间分布格局
3.3 不同龄级华山松的种内关联性
华山松的种内关联性表现为(图4):小树和中树在空间尺度1~2、3~9 m和14~50 m内的L12(r)函数值位于置信区间内,呈现不相关,L12(r)函数值在2~3 m和9~14 m的空间尺度内位于置信区间下方,呈显著负相关;小树和大树的L12(r)函数值在0~21 m和23~50 m的空间尺度上位于置信区间内,呈现不相关,在21~23 m的空间尺度内位于置信区间下方,呈显著负相关;中树和大树的L12(r)函数值在整个空间尺度(0~50 m)都位于置信区间内,呈现不相关,L12(r)函数值在空间尺度13 m处达到最大,为1.550 1。
3.4 华山松林优势植物的种间关联性
在所调查的样地中,华山松林优势植物有华山松、高山松和林芝云杉,它们的种间关联性表现为(图5):华山松和高山松的L12(r)函数值在0~17 m的空间尺度上位于置信区间下方,呈显著负相关,在18~50 m的空间尺度上位于置信区间内,呈现不相关,在空间尺度30 m处L12(r)函数值达到最大,为1.170 4;华山松和林芝云杉的L12(r)函数值在0~3 m及40~50 m的空间尺度上位于置信区间内,呈现不相关,但在3~40 m的空间尺度内位于置信区间下方,呈显著负相关;高山松和林芝云杉的L12(r)函数值在0~28 m的空间尺度内位于置信区间下方,呈显著负相关,在28~38 m及49~50 m的空间尺度上位于置信区间内,呈现不相关,在38~49 m的空间尺度内位于置信区间上方,呈显著正相关。结果说明不同的种群在相同的生长环境中表现出显著的差异,种群之间对于资源的竞争较为激烈。

图4 不同等级华山松的关联性
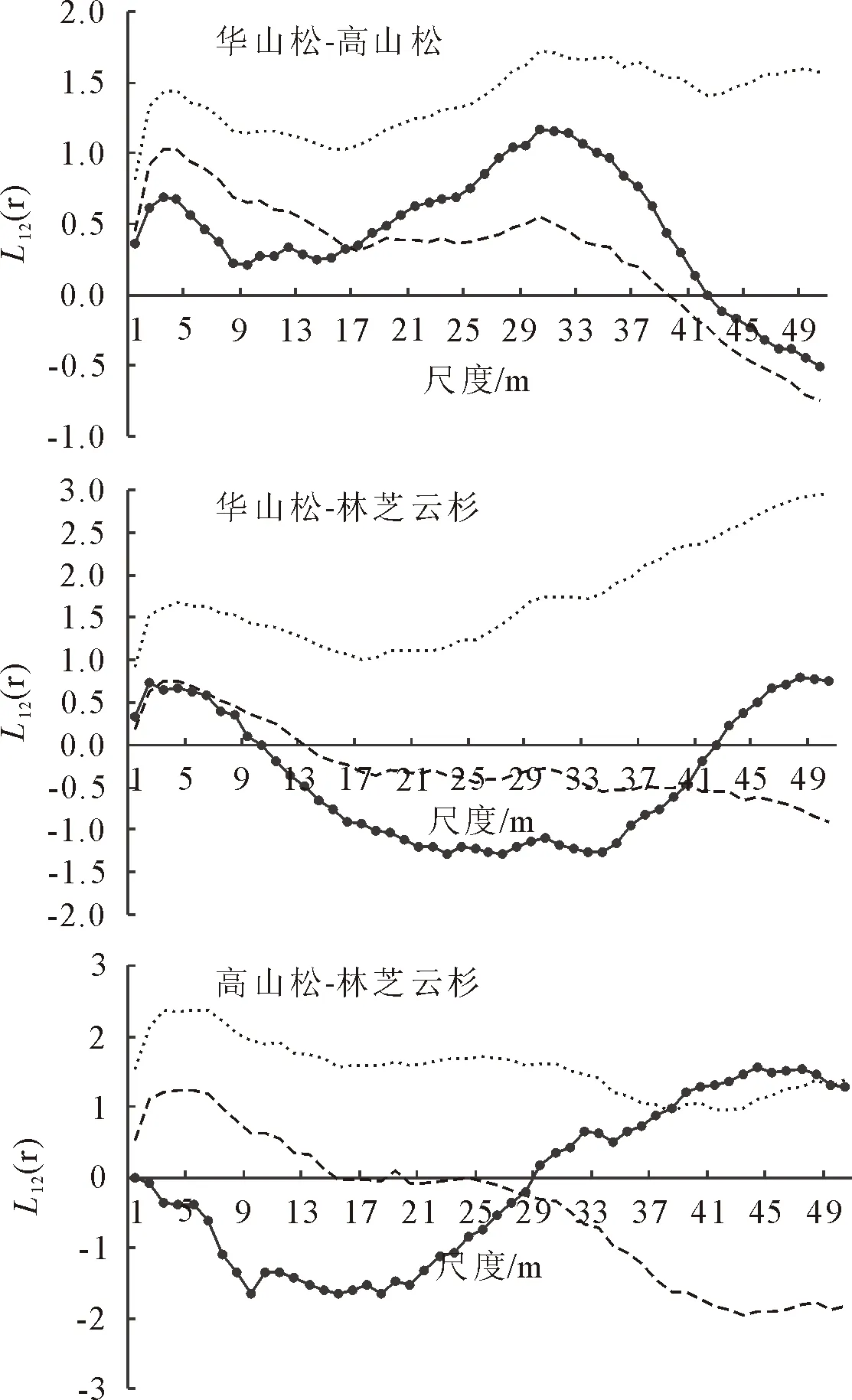
图5 华山松、高山松和林芝云杉的关联性
4 讨论
4.1 华山松林优势物种的空间分布格局
植物种群的空间分布类型决定于其自身的特性以及与环境之间的相互关系,不同的分布类型体现出种群对环境不同的响应机制,这样也有利于种群更好地发展[19-20]。集群分布体现出种群内部正向关系,集聚在一起共同抵御外界可能产生的不利因素,以此来提高存活率;随机分布则表现出个体之间关联较小,相互独立;均匀分布在分布类型中产生的几率较小,也体现出个体之间的负向关系[21-22]。本研究发现,华山松的小树在整体尺度上均表现为集群分布,这种现象与“聚集分布是植物种群最常见的分布形式”的观点一致,原因是小树的生命力较脆弱,在生长的过程中需要多个集聚起来应对外界不利因素,以此来提高存活率;中树在小尺度内呈集群分布,在大尺度空间内又表现为随机分布,可能因为华山松的球果较大,种子掩藏在球果中,无法进行风力传播,所以会集聚在母株周围,另外华山松的种子可以通过松鼠等啮齿类动物进行二次传播,但距离有限,因此在大尺度空间内会形成随机分布;大树在整个空间尺度上都表现为随机分布,原因是华山松的种子随机分布比集群分布的竞争力小,更容易获得生存所需要的资源,利于形成比较高大的华山松,所以大树在整个尺度上都是随机分布。在不同优势植物方面:华山松在整体尺度上都呈集群分布,可能因为华山松作为研究区的优势种,空间资源占有率较大,能够快速繁殖,因此易形成集群分布;高山松在不同空间尺度上分布格局波动较大,在小尺度内表现为集群分布,但集聚的程度随尺度增加而减小,接着是随机分布,在大尺度上呈均匀分布,可能是由于高山松在面对不同的环境因素所表现出不同的应对机制,以此来提高种群的生存力;林芝云杉几乎在整个尺度上都呈集群分布,可能因为在整个样地中相对于华山松和高山松的数量较少,需要聚集起来相互庇护,抵抗外界的干扰更好地生存下去。
4.2 华山松林优势植物的空间关联性
树种的空间分布格局及关联性可以有效揭示森林群落的生态过程,深入了解研究区的群落演替及对植被恢复提供借鉴。种内关联性是表示同一种群内不同生长发育阶段的个体在空间分布上的关联性,是对个体之间相互关系的描述[23-24]。种间关联性是不同物种之间在空间内的关联性,这也是群落演替和进化的基础[25-26]。研究表明,华山松的种内关联性几乎都呈不相关,这说明华山松的小树、中树和大树的关联性较弱,相互之间不会产生影响,原因可能是大树的数量较少,其分布密度也较小,因此对小树和中树的生长不会产生实质性的影响[27]。相反,华山松林优势植物的种间关联性不尽相同:华山松和高山松、华山松和林芝云杉在中小尺度上呈显著负相关,可能因为华山松处于优势地位,但会受到众多物种的排挤,因此与高山松和林芝云杉存在一种竞争关系,这也恰好体现出当物种间利用同一资源时,其相互间的竞争排斥作用就越强烈的物种竞争原理;在较大空间尺度上华山松和高山松、华山松和林芝云杉呈不相关,原因可能是为了更好地生存发展,三者最初较为激烈的资源争夺缓和为彼此之间无关联,尽可能做到共同生存;高山松和林芝云杉的空间关联性随着空间尺度的增加表现为负相关-不相关-正相关,原因可能是在生存发展的过程中,它们意识到了合作的重要性,起初相互竞争,后来共同利用资源,相互依赖相互促进,以此对抗优势种华山松,提高存活率。
4.3 华山松保护与恢复措施
根据岗乡华山松林自然分布区的生态环境以及干扰情况,加强对华山松的保护和抚育是其可持续发展的关键。首先建议采取就地保护措施,就地保护可以使植物避免很多外界干扰;其次要加强巡护、监管,以免人为破坏改变其原有的生存环境进而影响华山松的发展;最后加强基础研究,了解华山松的主要竞争者及其在发展过程中的影响因素。
5 结论
通过对藏东南波密岗乡华山松林优势植物空间格局及关联性研究,得出以下结论:1)华山松小树在整个空间尺度上呈集群分布;中树在小尺度内呈集群分布,大尺度内呈随机分布;华山松大树在整体空间尺度上都呈随机分布。2)研究区优势种华山松在整体尺度内表现为集群分布;高山松随空间尺度的增加表现为集群分布-随机分布-均匀分布-随机分布;林芝云杉在0~49 m的空间尺度内均呈现集群分布,但集聚程度随尺度的增加波动较大。3)华山松的种内关联性几乎在整体尺度上都表现为不相关,只有小树和中树在2~3 m和9~14 m的空间尺度内呈显著负相关。4)华山松的种间关联性类型较多,华山松和高山松、华山松和林芝云杉、高山松和林芝云杉在中小尺度上呈空间负相关,随空间尺度的增加逐渐表现为不相关,高山松和林芝云杉在较大空间尺度上呈正相关。
综上所述,波密岗乡华山松林优势植物-华山松、林芝云杉和高山松的空间分布格局及其关联性对于空间尺度的大小具有很强的依赖性,在不同的空间尺度上,种群的分布格局及关联性也存在较大的差异[28-30]。本研究有助于认识藏东南波密县华山松种内的共生过程以及种间复杂的竞争过程,了解到不同种群对于资源环境的响应机制,对波密县植被的恢复发展、物种的更新和可持续经营具有重要价值。
猜你喜欢
科海故事博览(2022年28期)2022-11-25
乡村科技(2021年19期)2021-12-16
现代农村科技(2020年7期)2020-12-19
中国森林病虫(2020年1期)2020-01-18
成都信息工程大学学报(2019年6期)2019-08-13
乡村科技(2019年14期)2019-02-21
新闻传播(2018年15期)2018-09-18
音乐天地(音乐创作版)(2018年4期)2018-07-18
启迪与智慧·教育版(2018年12期)2018-02-26
甘肃林业(2016年2期)2016-11-07
