
人釉原蛋白全长及其功能片段的体外自组装和矿化行为的研究
2021-08-14 07:02钟秀赖婷婷陈亮田鲲
华西口腔医学杂志 2021年4期
钟秀 赖婷婷 陈亮 田鲲
1.西南医科大学口腔医学院,泸州646000;2.四川省医学科学院·四川省人民医院口腔科,成都610072;3.遵义医学院,遵义563000
釉质覆盖在牙冠表面,作为人体最坚硬的组织具有强大的机械性能,但一经损伤不能再生。因此,釉质的仿生修复具有重要的科学价值和治疗需求。釉质发育的矿化过程由成釉细胞分泌的细胞外基质蛋白持续介导,其中釉原蛋白(amelogenin,AM)及其裂解产物占90%以上[1]。AM主要由含45个氨基酸的N端酪氨酸富集段(tyrosinerich amelogenin peptide,TRAP)、X-Y-脯氨酸重复序列组成的疏水性中央区域、相对较短含11个氨基酸的亲水性带电荷C末端3部分组成[2],其中亲水性C末端以及疏水性N末端高度保守,具有重要的生理学相关性,在釉质发生中发挥作用。
N末端主要影响AM的自组装并调节蛋白间的相互作用[3]。蛋白水解的N末端裂解形成较小的TRAP功能肽,参与了釉基质蛋白的相互作用过程,为组装中的各种釉基质蛋白提供分子连接位点[4]。亮氨酸富集段(leucine-rich amelogenin peptide,LRAP)由AM的26个C-末端氨基酸和33个N-末端氨基酸构成,包含了AM完整的带电C末端。LRAP主要影响对无机晶体的吸附,能稳定无定形磷酸钙(amorphous calcium phosphate,ACP),调节其晶体平行排列,引导ACP转化为有序的束状磷灰石晶体[5]。LRAP还能指导成釉细胞、骨细胞的分化[6],促进牙周组织的再生[7]等。
AM在釉质矿化过程中具有无法替代的地位,体外“仿生矿化模板”对AM及其功能片段的模仿程度越高,引导生成的羟磷灰石(hydroxyapatite,HA)晶体排列就越接近天然牙釉质,使得AM及其功能片段在体外诱导釉质再生领域中具有广阔的应用前景。但目前关于AM功能片段LRAP及TRAP的单独研究较少,且大多使用动物蛋白,如鼠、猪等。因此本实验重组表达了人AM及功能片段LRAP和TRAP,观察比较其自组装过程,探寻其从单分子克隆到矿化骨架的组合变迁。并在不同时间点添加AM、LRAP及TRAP以模拟体内蛋白作用顺序,观察不同时间段HA晶体的形成特点,旨在探讨AM和功能片段LRAP和TRAP在釉质发生及HA晶体各向生长过程中的可能机制,为釉质仿生修复治疗提供更多的参考和依据。
1 材料和方法
1.1 实验材料
选取正畸患者预防性拔除的处在釉质发育阶段的下颌第三磨牙牙胚(四川省医学科学院·四川省人民医院齿槽外科),全部取样均取得患者及其监护人同意。Trizol RNA分离试剂、质粒提取试剂盒(上海赛默飞世尔科技有限公司),pMD19-T载体(大连宝生物工程有限公司),pET-28a(+)载体、BL21(DE3)Chaprone感受态细胞、大肠杆菌Top10感受态细胞(四川省医学科学院·四川省人民医院人类疾病基因研究四川省重点实验室),大肠杆菌BL21plys(北京全式金生物技术有限公司),pCold-SUMO载体、Ni-NTA纯化树脂(杭州海基生物科技有限公司),4-羟乙基哌嗪乙磺酸(N-2-hydroxyethylpiperazine-N-ethane-sulphonicacid,HEPES)缓冲液、MgCl2、CaCl2·H2O、KH2PO4、KCl、NH4Cl、NaF(杭州众化科技有限公司)。
1.2 方法
1.2.1 AM全长的合成 从处于釉质发育阶段的儿童下颌第三磨牙的牙胚中提取牙胚细胞总RNA,设计AM全长上游引物序列为5’-CGCGGATCCATGGGGACCTGGATTTT-3’,下游引物序列为5’-CGAGCTCTTAATCCACTTCCTCCCGC-3’。通过逆转录聚合酶链反应(reverse transcription-polymerase chain reaction,RT-PCR)反转录、扩增得到AM全长的cDNA,与pMD19-T载体进行质粒构建后转入大肠杆菌Top10中扩增,筛选与鉴定后得到AM全长基因。AM全长基因与原核表达载体pET-28a(+)进行质粒构建,将重组质粒进行筛选和鉴定后用热激法转化到大肠杆菌BL21plys中,异丙基-β-D-硫代半乳糖苷(isopropyl-beta-D-thiogalactopyranoside,IPTG)诱导表达,十二烷基磺酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate polyacrylamide gelelectrophoresis,SDS-PAGE)检测,纯化,得到AM全长。
1.2.2 LRAP和TRAP片段的合成 用以上AM全长基因做模板,设计LRAP上游引物序列为5’-CGCGGATCCATGCCTCTACCACCTCATCCT-3’,下游引物序列为5’-CGAGCTCTTAATCCACTTCCTCCCGCTTG-3’;TRAP上游引物序列为5’-CGCGGATCCATGCCTCTACCACCTCATCCT-3’,下游引物序列为5’-CGAGCTCTTACCATCCACCCATGGGC-3’。聚合酶链反应(polymerase chain reaction,PCR)扩增构建LRAP、TRAP基因,克隆后测序鉴定。将LRAP、TRAP基因分别和pCold-SUMO原核表达载体进行质粒构建,鉴定后转化到BL21(DE3)Chaprone宿主菌中,IPTG诱导表达,表达产物进行SDS-PAGE验证,rTEV酶切后Ni-NTA纯化树脂纯化,得到LRAP、TRAP蛋白。
1.2.3 自组装实验 在50×三氯基甲烷(Tris-HCl,pH=8)中分别加入纯化的AM全长、LRAP、TRAP蛋白储备液(2 mg·mL-1),调节pH至8,最后再添加适量50×Tris-HCl(pH=8),在冰上建立最终的蛋白质溶液,浓度为100μg·mL-1。用毛细吸管吸取各蛋白溶液滴至铜网多孔碳侧,在室温下孵育1~15 min,分别在1、15 min时夹取铜网,滤纸吸去多余溶液,1%过滤磷酸-钨酸负染30 s,滤纸吸去染液,室温干燥后透射电镜(transmission electron microscope,TEM)观察其自组装过程。
1.2.4 体外矿化实验 配制人工唾液,在20 mmol·L-1HEPES缓冲液中加入一定量的CaCl2·H2O(1 mmol·L-1)、MgCl2(0.2 mmol·L-1)、KH2PO4(4 mmol·L-1)、KCl(16 mmol·L-1)和NH4Cl(4.5 mmol·L-1),调节pH至8,使用前加入NaF(3×10-4)。在玻片上滴加2 mL人工唾液,滴入AM蛋白储备液至AM蛋白浓度为100μg·mL-1,孵育3 d后扫描电镜(scanning electron microscope,SEM)观察新生晶体形貌;再滴入等量TRAP、LRAP蛋白储备液至TRAP、LRAP蛋白浓度为100μg·mL-1,继续孵育3 d,SEM再次观察新生晶体形貌。
1.2.5 X射线衍射分析 采用DMAX-3A型X射线多晶衍射仪对矿化6 d的新生晶体进行分析,工作条件为:CuKα射线,电压70 kV,电流50 mA,衍射起始角10°,衍射终止角80°,步长0.02°。
2 结果
2.1 AM全长及LRAP、TRAP的合成和鉴定
PCR扩增构建的AM全长、LRAP、TRAP基因经1%琼脂糖凝胶电泳后,可分别见570、230、130 bp的特异性条带,与AM全长、LRAP、TRAP基因理论大小基本相符(图1)。对获得的重组蛋白进行SDS-PAGE后,可分别见大小23 000、5 000、6 000的条带,与AM全长、LRAP及TRAP的理论相对分子质量基本符合,表明成功重组并表达了AM全长、LRAP及TRAP蛋白(图2)。
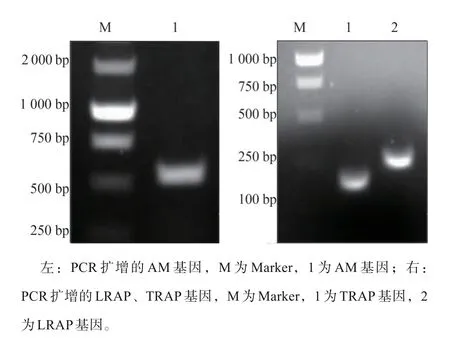
图1 AM全长及LRAP、TRAP基因的重组Fig 1 Recombination of AM,LRAPand TRAPgenes
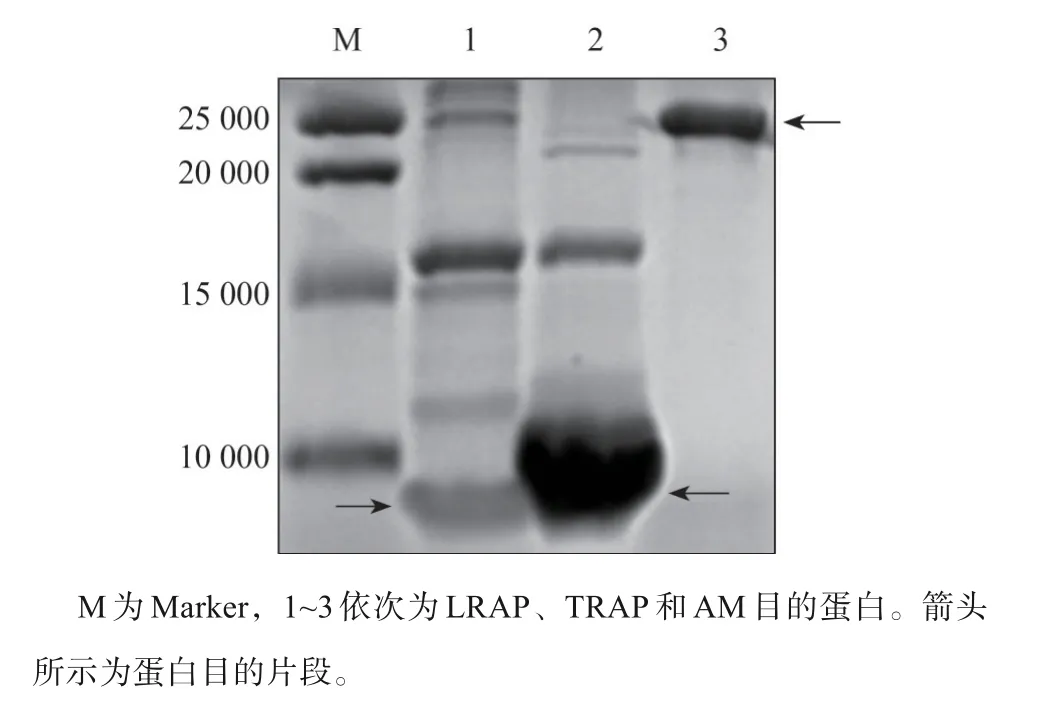
图2 AM全长及LRAP、TRAP的SDS-PAGE图谱Fig 2 SDS-PAGEof AM,LRAPand TRAP
2.2 AM全长的自组装结果
蛋白液孵育1 min时,溶液中的AM全长大部分以单体形式存在,单体直径为1~2 nm。继续孵育15 min时,蛋白单体自组装形成了直径20~30 nm的“纳米球”。这些纳米球进一步聚集,可见3个纳米球成角串起形成的“三角形”结构,数个纳米球排列形成的“串珠样”结构以及多个纳米球聚集形成的有进一步排列成链状、网状趋势的不规则结构,呈现AM全长的分级自组装(图3)。
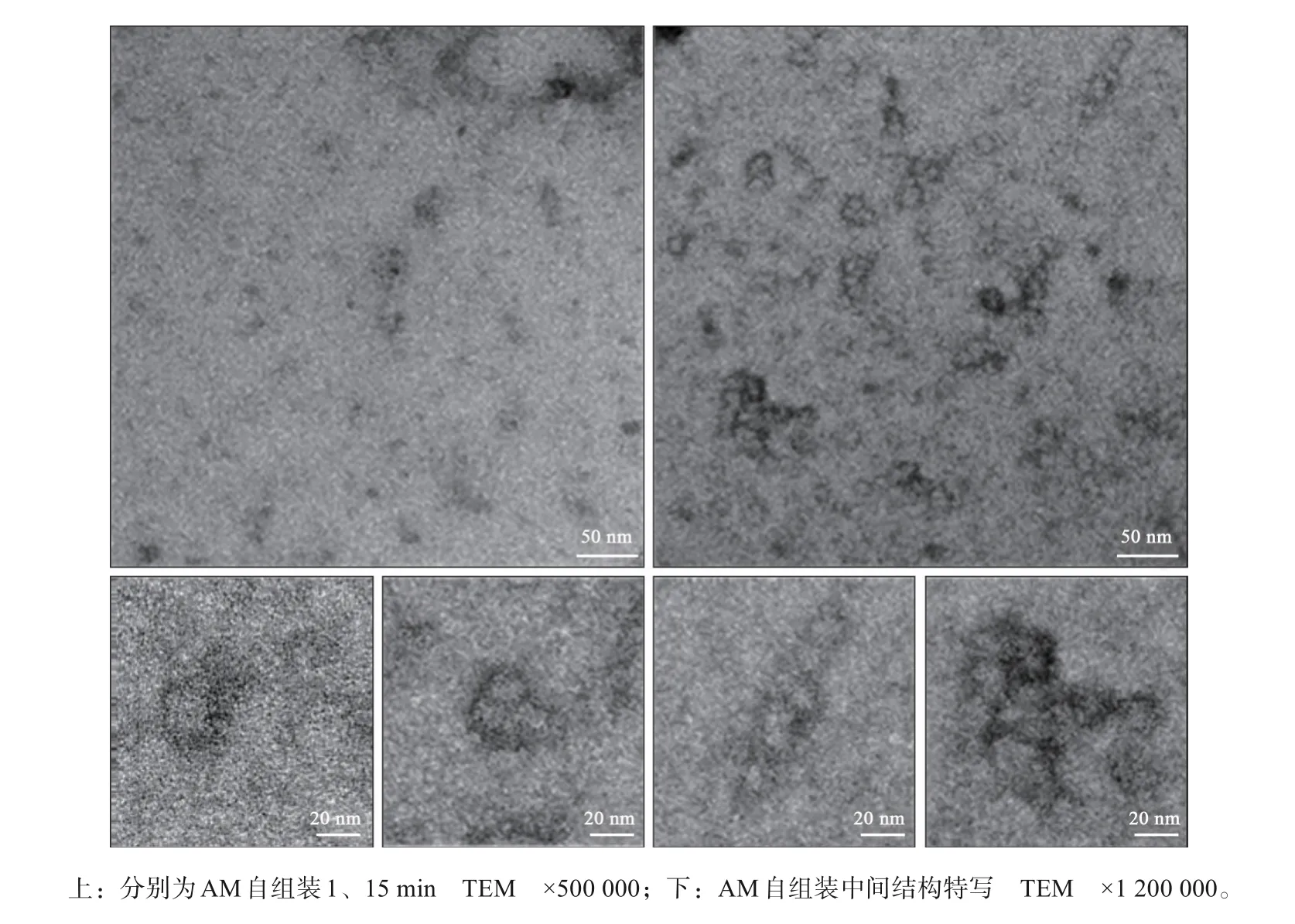
图3 AM全长自组装的观察结果Fig 3 Micrographs of full-length AM self-assembly
2.3 LRAP的自组装结果
LRAP蛋白溶液在孵育1 min时,溶液中的LRAP蛋白多以单体形式存在,孵育15 min后TEM下仍不能明显观察到LRAP蛋白自组装形成的纳米球及中间结构,溶液中的蛋白优势形态为无结构的单体形态,也可见少量低聚物(图4)。
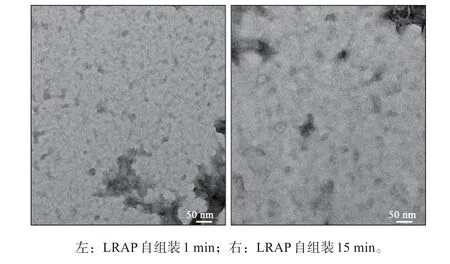
图4 LRAP自组装的观察结果 TEM ×500 000Fig 4 Micrographs of LRAP self-assembly TEM ×500 000
2.4 TRAP的自组装结果
TRAP蛋白溶液在孵育1 min时,溶液中仅见蛋白单体。孵育15 min后,溶液中可见明显的直径20~50 nm的纳米小球。TRAP蛋白形成的纳米球形状十分规则,呈均匀的圆球形,没有明显的内部结构,外周有月晕状的黑色光圈,且纳米小球的直径越大,其外周的光圈越明显。TRAP蛋白由单体到纳米球的形成中无分级组装过程,且组装形成的纳米小球之间无进一步聚集的趋势,相互之间孤立存在(图5)。
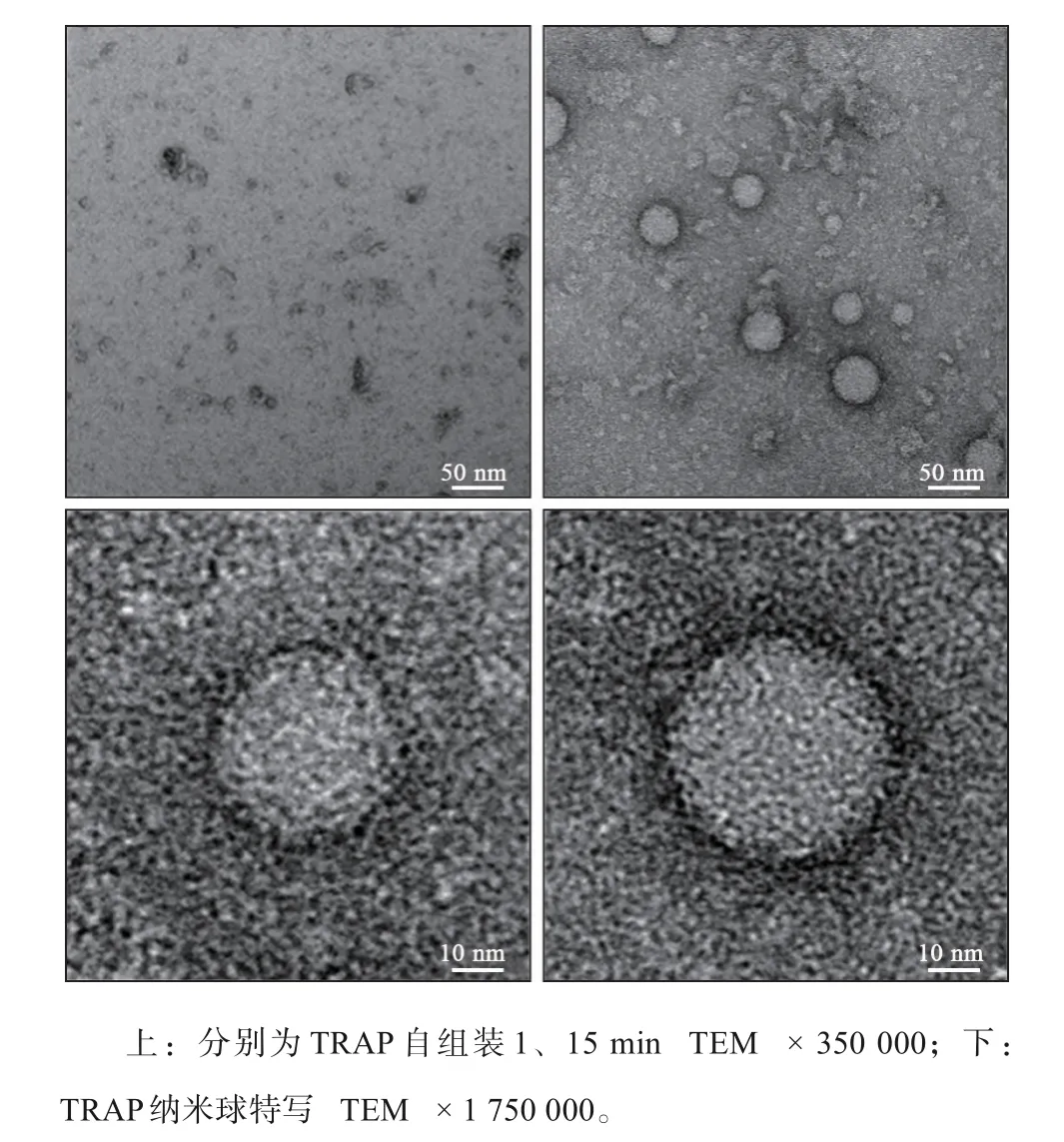
图5 TRAP自组装的观察结果
2.5 体外矿化实验结果
AM在人工唾液中孵育3 d后,可见其引导的新生晶体呈细长棒状,沿c轴取向,直径55~60 nm,平均长度800 nm,数个晶体相互呈一定角度捆绑形成直径220 nm的晶体束。加入TRAP、LRAP孵育3 d后,新生晶体在a、b面生长欠佳,直径无明显变化(60~65 nm),而在c轴上显著伸长(1 200 nm);晶体较AM单独诱导数量更多、取向更佳,形成直径为500 nm的晶体束,绝大部分细长棒状晶体有序近似平行排列,仅有晶体束最外层的少量晶体与晶体束长轴成较大角度排列(图6)。

图6 不同时间点加入相应蛋白诱导生成的晶体Fig 6 The crystals induced by the corresponding proteins were added at different time
2.6 X射线衍射分析结果
X射线衍射图谱在2θ衍射角为25.8°、32°、32.2°和32.9°处出现强度不同的衍射峰,分别与HA晶体(002)、(211)、(112)、(300)面的特征峰一致,表明AM及TRAP、LRAP诱导新生的晶体为HA晶体(图7)。
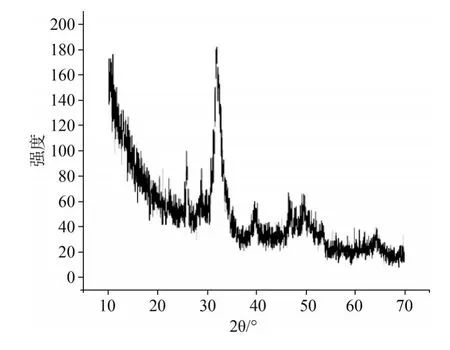
图7 新生晶体的X射线衍射图Fig 7 XRD pattern of thenewborn crystal
3 讨论
成熟釉质主要由HA晶体组成,这些晶体平行排列,形成釉柱,而釉柱又定向排列成高度有序的釉质结构。釉质中的HA晶体非常狭长、呈六方棱柱形,在c轴方向延长,表观长度一般为250~1 000 nm,宽 度 为60~100 nm,厚 度 为26.3~35.0 nm。与天然HA晶体一样,釉质中的HA晶体由六面晶体的晶胞组成,晶胞六边形边长为0.943 2 nm,长度为0.688 nm[8]。
成釉细胞在分泌期合成的有机基质可分为AM和非釉原蛋白两类,其中AM占90%以上。发育中天然牙的釉质基质里,AM及其酶解产物融合成较大尺寸的纳米结构而发挥作用,在釉质成熟后则基本消失。AM分泌到釉基质中很快被裂解,在分泌早期,基质金属蛋白酶-20(matrix metalloproteinase-20,MMP-20)在有限且相同的位点裂解或加工全长AM,产生富含酪氨酸的TRAP、相对分子质量为23 000的中间肽段和不同的C末端片段;而在成熟期,激肽释放酶4(kallikrein4,KLK4)会特异性地将MMP-20处理后的釉基质蛋白降解为更小的多肽,被成釉细胞彻底吸收,无机物含量能达到95%,控制着釉质的赋形和矿化率[9]。
在体外,AM在pH=8和37℃环境中,N端疏水的脯氨酸富集区域相互聚拢,C端亲水的带电荷氨基酸尾巴TLAGGVA朝向外,形成直径20 nm的球状纳米自组装体,然后纳米小球多向延伸,组装成纳米链和纳米网状结构[10],与本实验结果相符。而本实验结果表明:TRAP片段形成没有亲水氨基酸尾巴的纳米微球,且纳米球之间没有相互进一步聚集成链状、网状的趋势;LRAP片段则一直处于无结构的单体形态;AM单独诱导3 d后形成棒状HA晶体,再加入TRAP、LRAP蛋白诱导3 d后,HA晶体在c轴方向快速择优生长,而a、b平面生长被抑制,且晶体取向更优。在磷酸八钙晶体(octalcium phosphate,OCP)的生长实验中,AM的疏水部分在控制晶体形状方面起作用,最有可能是通过在a、b晶体平面上的选择性吸附来抑制a、b平面的生长,这一抑制过程与C端区域无关,而带负电荷亲水的C端区域能通过Glu和Asp残基侧链羧基之间的金属离子螯合作用调节OCP晶体沿c轴方向相互平行排列[11],与本实验结果相符。
结合本实验的结果及HA晶体的生长过程来看,可能的机制是,在釉质矿化的生长基元形成期,AM分级自组装形成球形超分子结构,COOH-端亲水尾部基团与钙离子发生交联反应,启动矿化过程,纳米球开始HA成核然后继续和釉原蛋白COOH-端的负电基团紧密结合。由于HA晶体表面电荷分布的空间匹配,纳米球聚集在其周围沿着晶体的c轴包裹晶体,组装成纳米长链,这种长链结构为晶体的成核和c轴方向生长提供有序模板,并阻止a、b晶面融合生长趋势。随后在生长基元叠合期,MMP-20选择性降解AM,形成一些功能小分子蛋白片段,其中疏水性的TRAP及中间肽段组装成纳米微球,包覆a、b轴面,因其负电性及亲水性均减弱,屏蔽了HA晶体a、b轴向的晶面结合位点。与此同时,c轴上成核位点暴露,钙磷盐不断沉积,晶体迅速沿着c轴择优生长,在亲水带负电LRAP的调节下相互平行排列。最后,在KLK4的作用下,大部分蛋白降解后被成釉细胞彻底消化,蛋白与HA晶体的结合降低,对a、b轴面的包覆作用减弱,成核位点暴露,晶体得以沿着a、b轴方向生长、变粗,最终成熟。
许多研究已经实现了AM促进釉质形成的过程,并肯定AM的重要作用,但AM全长及其被降解的功能片段完成一系列引导HA晶体不同时机不同轴向生长的具体机制还未完全清楚。本实验对比了AM全长、TRAP和LRAP的自组装行为,并根据以上蛋白多肽在釉质发育过程中的大致作用时间顺序设计体外矿化实验,结果表明:AM全长分级自组装形成直径20~30 nm的“纳米球”后有进一步多向延伸组装成链状、网状趋势;疏水的TRAP组装成直径20~50 nm的孤立纳米球,无分级组装过程;而LRAP自组装行为不明显,多以无定形单体形式存在;AM在体外单独诱导可形成细长的棒状HA晶体,继续加入TRAP、LRAP蛋白后HA晶体在c轴明显延长,而a、b平面生长欠佳,且晶体取向更优。本实验结果与体内HA晶体定向生长的机制契合,为它们在HA晶体形成过程中的作用提供了理论依据。
利益冲突声明:作者声明本文无利益冲突。
猜你喜欢
生殖医学杂志(2022年10期)2022-10-19
农业机械学报(2022年7期)2022-08-08
口腔医学(2021年10期)2021-12-02
中风与神经疾病杂志(2021年9期)2021-11-08
能源工程(2021年3期)2021-08-05
中小学德育(2020年11期)2020-03-18
口腔疾病防治(2020年2期)2020-02-26
标记免疫分析与临床(2016年9期)2016-11-21
医学研究杂志(2015年9期)2015-07-01
