
一株鸡源性沙门氏菌的基因组分析
2021-08-11 02:58:34张海韵郭焜鹏王芳斌
生命科学研究 2021年3期
宋 晟,张海韵,郭焜鹏,王芳斌
(食品安全监测与预警湖南省重点实验室湖南省食品质量监督检验研究院,中国湖南长沙410117)
沙门氏菌是一种嗜温性、兼性厌氧、无芽孢、无荚膜的革兰氏阴性杆菌,在自然界中广泛分布,为食源性细菌性肠胃炎的首要病原菌。典型沙门氏菌菌株多具有周生鞭毛,能运动、能分解葡萄糖并产气[1~2]。当前,全球每年因食品中沙门氏菌污染而引起的感染病例数以亿计,生熟不分是家庭内流行最常见的原因。在国家卫生系统的全国食物中毒事件情况通报中,沙门氏菌引起的食物中毒占总数的50%以上。我国食品安全国家标准中规定,各类预包装食品中不得检出沙门氏菌,但对于生鲜肉制品并无沙门氏菌的限量要求。目前,各地各类生鲜肉制品中沙门氏菌的分布情况和检出情况均有所不同[3~4]。本研究从湖南省长沙市生鲜市场处采购生鲜鸡肉,依据《食品安全国家标准食品微生物学检验沙门氏菌检验》(GB 4789.4—2016),分离鉴定出一株沙门氏菌(FC10727),并对其进行全基因测序与生物信息学研究,可为沙门氏菌基因层面的深入研究提供一定的实验基础,为科学防范、食品安全监管、政策制定提供科学依据。
1 材料与方法
1.1 材料
生鲜鸡肉(2 kg)购于湖南省长沙市雨花区一生鲜市场。
缓冲蛋白胨水(buffered peptone water,BPW)、四硫酸钠煌绿增菌液(tatrathionate broth base,TTB)、亚硒酸盐胱氨酸增菌液(selenite cystine broth,SC)和HE琼脂(Hektoen enteric agar)均购自青岛海博生物技术有限公司;亚硫酸铋琼脂(bismuth sulfite agar,BS)、木糖赖氨酸脱氧胆盐琼脂(xylose lysine desoxycholate agar,XLD)、营养琼脂(nutrient agar,NA)以及三糖铁琼脂(triple sugar iron agar,TSI)购自北京陆桥技术股份有限公司;沙门氏菌显色培养基和革兰氏阴性细菌鉴定卡(Gram-negative identification card,GN)购自法国Chromagar公司。
1.2 增菌
生鲜鸡肉按照500 mL/kg的比例加入BPW,对生鲜鸡肉进行充分的润洗,反复揉搓2~3 min后(36±1)℃培养18 h。吸取1 mL BPW培养物转种到10 mL的TTB中,(42±1)℃培养24 h;另吸取1 mL BPW培养物至10 mL SC中,(36±1)℃培养24 h。
1.3 分离
用无菌接种环从TTB选择性增菌液与SC选择性增菌液中分别取增菌液1环(10 μL),划线XLD、BS和HE平板。BS平板和XLD平板置于(36±1)℃培养48 h,HE平板置于(36±1)℃培养24 h。
1.4 生化试验
从XLD平板上挑取中心为黑色、周边为无色透明环的菌落;从BS平板上挑取棕褐色、灰色或黑色带金属光泽的菌落;从HE平板上挑取中心为黑色、周边为无色或蓝绿色的菌落,将挑取的菌落接种三糖铁琼脂,先在斜面划线,再于底层穿刺,最后(36±1)℃培养 24 h。
1.5 鉴定
从三糖铁琼脂结果为斜面产碱、底层产酸、产气、产H2S的斜面上挑取菌落,接种至营养琼脂并于(36±1)℃培养24 h。对得到的纯化单菌落采用全自动微生物生化鉴定系统(VITEK 2,法国生物梅里埃公司)进行菌种鉴定。
1.6 全基因组测序
委托广州研科生物科技有限公司对全自动微生物生化鉴定结果为沙门氏菌属的菌落进行基因组DNA的提取、测序、功能基因预测及注释等工作。
2 结果
2.1 菌种分离及鉴定结果
BPW润洗液经过18 h培养变浑浊,转接TTB和SC培养24 h后,培养液也均变浑浊。将TTB与SC增菌液分别接种BS、HE和XLD鉴定培养基,培养结果呈现不同特征。其中,BS琼脂中的菌落为黑色有金属光泽,菌落周围培养基呈黑色;HE琼脂中的菌落呈蓝绿色,带黑色中心;XLD琼脂中的菌落呈粉红色,带黑色中心。将3种培养基中鉴定出的菌落进一步接种三糖铁琼脂,结果显示:斜面产碱,底层产酸,产H2S,产气(图1)。FC10727经全自动微生物生化鉴定系统鉴定为沙门氏菌属。

图1 FC10727在BS、HE、XLD与TSI培养基上的形态特征Fig.1 Morphological characteristics of FC10727 on BS,HE,XLD and TSI media
2.2 FC10727基因组序列的基本信息
通过Illumina HiSeq 4000平台测序,组装结果表明,FC10727基因组大小为4 638 420 bp,预测有4 016个基因,GC含量为54.15%,总长度为3 849 972 bp,平均长度为788 448 bp,占基因组全长的83%。另外,tRNA预测数有74个,5S rRNA有4个,16S rRNA有1个,sRNA有131个。绘制FC10727的全基因组图谱,结果如图2所示。
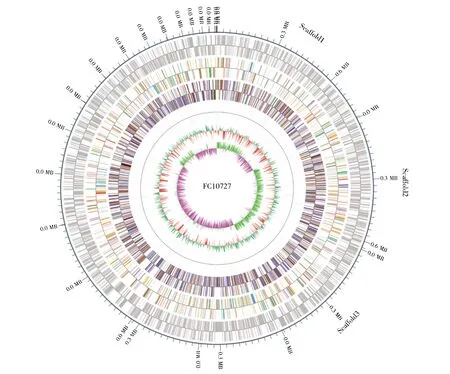
图2 FC10727全基因组图谱最外圈是基因组序列位置坐标,由外到里,分别是编码基因,COG(Cluster of Orthologous Groups of proteins)、KOG(clusters of orthologous groups for eukaryotic complete genomes)、eggNOG(evolutionary genealogy of genes:Non-supervised Orthologous Groups)、KEGG(Kyoto Encyclopedia of Genes and Genomes)、GO(Gene Ontology)数据库基因功能注释信息,非编码 RNA(noncoding RNA,ncRNA),基因组GC含量,基因组GC skew值分布。Fig.2 Genomic map of the Salmonella strain FC10727The outermost circle is the position coordinates of the genome sequence.From outside to inside,they are coding genes,Cluster of Orthologous Groups of proteins(COG),clusters of orthologous groups for eukaryotic complete genomes(KOG),evolutionary genealogy of genes:Non-supervised Orthologous Groups(eggNOG),Kyoto Encyclopedia of Genes and Genomes(KEGG),Gene Ontology(GO)database gene function annotation information,non-coding RNA(ncRNA),genome GC content,and genome GC skew value distribution.
将基因组测序结果提交NCBI数据库进行比对,比对结果显示第1位是肠道沙门氏菌。沙门氏菌属包括肠道沙门氏菌和邦戈沙门氏菌两类,其中肠道沙门氏菌广泛存在于猪、牛、羊、家禽、鸟类、鼠类等多种动物的肠道和内脏中,是世界各国分离率最高的菌型之一。该菌能引起各种家禽和哺乳动物的传染病,也可引起人类感染,具有重要的公共卫生意义。许多家禽(鸡、鸭、鸽等)、家畜(猪、牛、羊、马、狗、猫等)、鼠类和飞鸟的肠道中储存有这种细菌,所以家禽、家畜、鼠类、病人和带菌者是主要的传染源。
2.3 FC10727基因组的GO预测分析
将FC10727基因组序列提交至GO数据库进行比对,结果显示与其对应的基因共2 901个。将这些基因信息依据GO分类标准进行分类,可分为分子功能、细胞组分、生物过程3大类,42小类。
分子功能类基因包括:抗氧化活性、结合、催化活性、通道调节活性、酶活调节活性、分子变换活性、核酸结合翻译活性、蛋白质结合翻译活性、分子结构活性、转运活性等相关基因,共10小类,其中结合与催化活性相关基因最多,均超过1 000个;细胞组分类基因包括:细胞、细胞组分、细胞外区域、大分子复合物、膜封闭腔、细胞器、病毒体等相关基因,共10小类,其中细胞与细胞组分基因最多,有1 064个;生物过程类基因包括:生物黏附、生物调节、细胞成分组织与生物发生、细胞过程、死亡、发育过程、细胞定位与建立、免疫系统过程、定位、运动、代谢过程、多组织过程、多细胞生物过程、生物过程负调控、氮利用、积极调控生物过程、生物过程调节、复制、复制过程、应激反应、信号系统、病毒繁殖等相关基因,共22小类,其中细胞过程与代谢过程基因最多,均超过1 500个(图 3)。
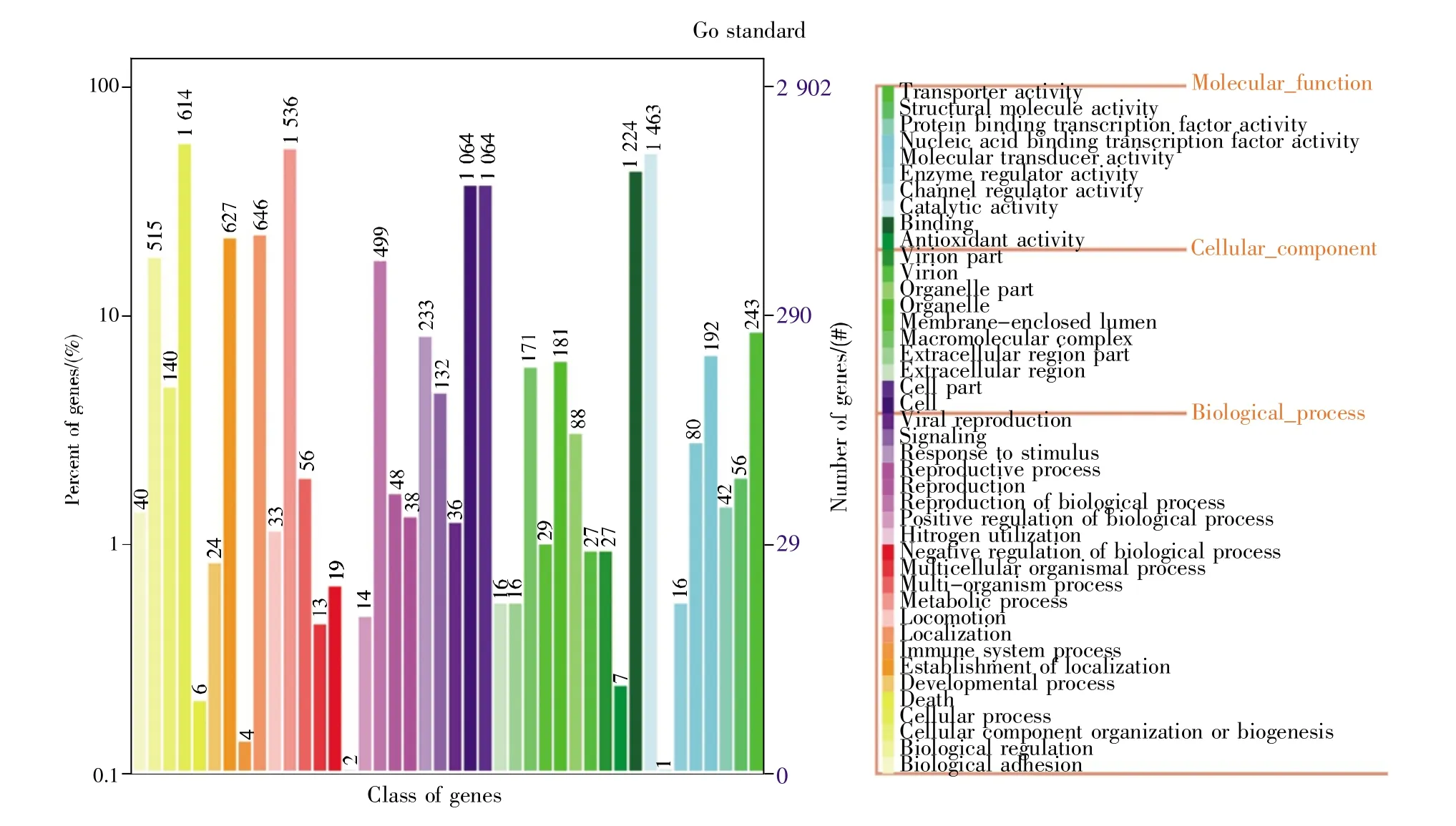
图3 FC10727基因在GO数据库的分类左图为基因数量;右图为基因功能,可分为分子功能、细胞组分、生物过程3大类。Fig.3 GO classification for the Salmonella strain FC10727 genesThe left picture shows the number of functional genes,and the right picture shows the prediction of gene functions,which is divided into three categories:molecular function,cellular component,and biological process.
2.4 FC10727基因组的KEGG预测分析
通过比对KEGG数据库发现,FC10727基因组中与其对应的基因共2 981个,按功能可分为细胞过程、环境信息处理、遗传信息处理、人类疾病、代谢、生物系统6大类,包含24小类代谢通路,分别对应不同基因序列表达产物。
细胞过程类基因涉及运输和分解代谢系统、细胞运动系统、细胞生长与死亡3小类,其中细胞运动系统基因最多,共34个;环境信息处理类基因涉及信号转导和膜转运两小类,基因数均超过100个;遗传信息处理类基因涉及翻译、复制和修复以及折叠、分类和降解3小类,其中翻译类基因最多,共80个;人类疾病类基因涉及传染性疾病、免疫疾病、耐药性3小类,其中传染性疾病基因最多,共31个;代谢类基因涉及异生物素的生物降解和代谢、核苷酸代谢、萜类和聚酮化合物的代谢、其他氨基酸代谢、辅助因子和维生素的代谢、脂质代谢、聚糖的生物合成与代谢、能量代谢、碳水化合物代谢、其他次级代谢产物的生物合成、氨基酸代谢,共11小类,其中碳水化合物代谢基因最多,共295个;生物系统类基因涉及内分泌系统和消化系统两小类,各1个基因(图4)。
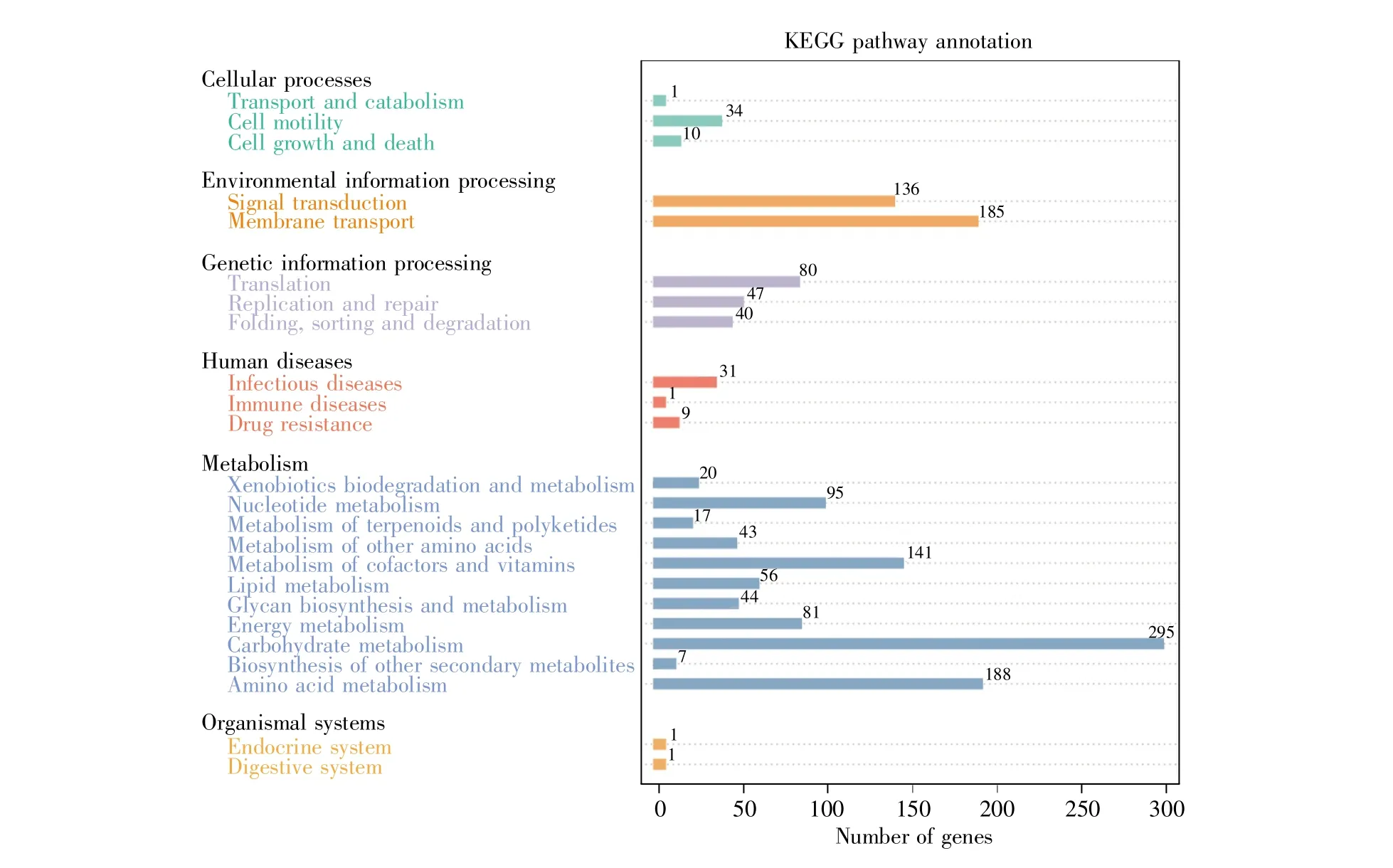
图4 FC10727基因的KEGG代谢通路分类左图为基因功能,可分为细胞过程、环境信息处理、遗传信息处理、人类疾病、代谢、生物系统6大类;右图为基因数量。Fig.4 KEGG pathway classification for the Salmonella strain FC10727 genesThe left picture shows the prediction of functional genes,which can be divided into six categories:cellular processes,environmental information processing,genetic information processing,human diseases,metabolism,and organismal systems.The right picture shows the number of corresponding functional genes.
2.5 FC10727基因组的eggNOG预测分析
将FC10727基因组与eggNOG数据库进行比对,得到与其相关的基因共944个。将这些基因信息依据GO分类标准进行分类,共有20类。
代谢类基因共252个,涉及氨基酸运输与代谢、核苷酸运输与代谢、碳水化合物运输与代谢、脂质运输与代谢、辅酶运输与代谢以及转运、分解代谢与次生代谢产物的生物合成;细胞功能类基因共95个,涉及细胞周期控制、细胞分裂、染色体分区、细胞壁/膜/包膜生物发生与细胞运动;分子功能基因共243个,涉及翻译、核糖体结构和生物发生、转录、复制、重组和修复、翻译后修饰、蛋白质转换和分子伴侣;运输与信号转导功能基因共123个,涉及无机离子的运输与代谢、信号转导机制、细胞内运输、分泌和囊泡运输。此外,其他类型的基因有能量生成与转化基因91个、防御机制基因12个、未知功能基因174个等(图5)。

图5 FC10727基因在eggNOG数据库的分类左图为基因数量;右图为基因功能,可分为20类,主要与氨基酸运输与代谢、能量生成与转化、无机离子的运输与代谢、碳水化合物运输与代谢等生物功能有关。Fig.5 eggNOG classification for the Salmonella strain FC10727 genesThe left picture shows the number of corresponding functional genes,and the right picture shows the prediction of functional genes,which can be divided into 20 categories.They are mainly related to amino acid transport and metabolism,energy production and conversion,inorganic ion transport and metabolism,and carbohydrate transport and metabolism,etc.
2.6 FC10727基因组的CAZy预测分析
FC10727基因组与CAZy数据库的比对结果显示,与其相关的基因共96个。依据CAZy分类标准进行分类,FC10727基因组可被划分为5类,其中,碳水化合物结合结构域(carbohydrate binding domain,CBM)类共18个基因;碳水化合物酯酶(carbohydrate esterase,CE)类共4个基因;糖苷水解酶(glycoside hydrolase,GH)类共41个基因;糖基转移酶(glycosyltransferase,GT)类共34个基因;氧化还原酶(oxidoreductase,AA)类共1个基因;不存在多糖裂解酶(polysaccharide lyase,PL)相关基因。
2.7 FC10727耐药性基因的预测分析
为了深入了解FC10727的耐药机制,进一步分析耐药基因功能,发现14个功能基因与抗生素耐药性相关。其中8个基因与β-内酰胺类抗生素耐药性相关,包括 sei:SPC_0458、stt:t2384、set:SEN3224、sei:SPC_3260、kpe:KPK_1516、see:SNSL254_A2451、sei:SPC_1000 和 spe:Spro_3259,涵盖外膜孔蛋白基因3个、外膜通道蛋白基因1个、膜融合蛋白基因2个、β-内酰胺酶基因1个。sei:SPC_0458具有β-内酰胺酶感应信号传感器AmpG的功能,属于调节基因;stt:t2384与set:SEN-3224编码的蛋白质属于膜融合蛋白acrA;sei:SPC_3260编码的蛋白质属于外膜通道蛋白TolC;kpe:KPK_1516与see:SNSL254_A2451编码的蛋白质属于外膜孔蛋白ompC;sei:SPC_1000编码的蛋白质属于外膜孔蛋白ompF;spe:Spro_3259编码的蛋白质属于β-内酰胺酶。另外,1个基因(sew:SeSA_A1717)与万古霉素耐药性有关,其编码的蛋白质属于D-丙氨酰-D-丙氨酸二肽酶;1个基因可表达位于沙门氏菌细胞外膜的酰基转移酶PagP;4个基因的编码产物分别对应厌氧型一氧化氮还原酶norV、一氧化氮还原酶FoRd-AND(+)norW、厌氧型一氧化氮还原酶转录调节因子norR和Rrf2家族转录调节因子nsrR,主要起到还原一氧化氮的作用。
2.8 FC10727致病基因的预测分析
为了深入了解FC10727的致病机制,进一步分析致病基因功能,发现27个功能基因与沙门氏菌致病性相关,其中1个基因(seh:SeHA_C3828)与免疫系统疾病有关,可引起系统性红斑狼疮;26个基因与传染性疾病有关。在26个传染性疾病相关基因中,8个基因与沙门氏菌侵袭上皮细胞相关:invasin A、invasin B、invasin C 和invasin D 4个基因起到侵袭素的作用;sopD和sptP的编码产物分泌效应蛋白,在入侵真核细胞中起协同作用;yeeJ起到黏附素的作用,介导沙门氏菌对宿主细胞的黏附;sopB编码的磷脂酰肌醇-4,5-双磷酸4-磷酸酶是一种肌醇磷酸酶,在发病机理中起到重要作用。相似性分析显示,invasin A、invasin B、invasin C、invasin D 及 sopB 与志贺氏菌致病基因高度相似。另外,本研究发现4种Ⅰ型菌毛蛋白基因fimA在细菌和宿主细胞的相互作用中发挥重要毒力作用;6种OmpD基因可表达外膜引入蛋白,参与巨噬细胞和肠上皮细胞对细菌的识别;1个ipaH9.8基因存在于侵袭性大质粒,其基因产物为效应分子,与志贺氏菌致病基因高度相似。
3 讨论
随着全基因组测序技术在国内外的迅速发展,生物信息学成为研究致病菌基因组整体功能特别是致病性、耐药性的重要手段。本研究对从新鲜鸡肉中分离出的一株沙门氏菌FC10727进行了全基因组测序,并将基因组信息提交至eggNOG数据库、KEGG数据库、GO数据库以及CAZy数据库进行匹配,对参与合成和代谢过程(如细胞膜合成、能量转化产生、转录、碳水化合物及氨基酸运输等)的基因进行了预测与分析,重点对致病基因及耐药基因进行了分析。
研究发现,FC10727可能有多种耐药机制。首先,基于β-内酰胺类抗生素耐药基因的分析,可知FC10727通过多种基因或蛋白质协同作用获得β-内酰胺类抗生素耐药性,可能的机制有以下几种:一是β-内酰胺酶使易感抗生素水解而灭活;二是β-内酰胺酶与抗生素迅速、牢固结合,使其停留于胞膜外间隙中,不能进入靶位,从而产生耐药性;三是细菌接触抗菌药物后,发生外膜孔蛋白ompC和ompF的表达基因失活,造成孔蛋白丢失或严重减少,最终导致β-内酰胺类药物进入菌体内减少,切断β-内酰胺类药物输入细菌体内的途径,以产生耐药性;四是主动外排系统,膜融合蛋白acrA捕获抗菌药物后,通过外膜通道蛋白TolC,将抗菌药物运转至外界[5]。其次,基于万古霉素耐药基因的分析,预测FC10727可能通过D-丙氨酰-D-丙氨酸二肽酶,将细胞壁中合成的D-丙氨酰-D-丙氨酸水解,导致细胞壁前质末端的二肽发生结构性变化,从而降低万古霉素药物分子跟细菌细胞壁前质的结合力,以获得对万古霉素的耐药性[6]。再者,基于酰基转移酶基因PagP的分析,预测FC10727可表达位于沙门氏菌细胞外膜的酰基转移酶PagP,并能将磷脂的C16碳脂肪酸链转移到内毒素分子的脂肪酸链上,形成次级脂肪酸链,PagP产生的内毒素可以干扰Toll样受体4(Toll-like receptor 4,TLR4)的识别,使沙门氏菌对阳离子抗菌肽产生抗性[7]。最后,基于一氧化氮还原酶基因norV和norW以及一氧化氮还原酶转录调节因子编码基因norR和nsrR的分析,预测沙门氏菌可通过还原一氧化氮,实现对一氧化氮的脱毒,导致吞噬细胞无法杀死沙门氏菌[8]。
沙门氏菌是肠道致病菌,其发病机理是菌体摄入机体后,通过胃进入肠道,抵抗肠道蠕动,黏附并定殖到肠道内,呈分子内寄生,释放或不释放肠毒素,引起组织病理损伤。通过对沙门氏菌致病基因的预测可知,FC10727含有多种毒力因子,包括侵袭因子、效应蛋白、黏附素、肌醇磷酸酶、菌毛蛋白、外膜引入蛋白和效应分子,它们通过协同作用,产生综合毒力。其中,侵袭因子invasin A位于致病岛SPI-1,决定细菌进入宿主上皮细胞的能力,是沙门氏菌致病的关键性毒力因子;效应蛋白sopD和sptP在细菌入侵真核细胞中起协同作用;yeeJ起到黏附素的作用,对沙门氏菌的定植起着重要的作用,在化学性质方面,黏附素可能为沙门氏菌表面某种特定的蛋白质结构或糖脂成分[9~12];肌醇磷酸酶sopB在发病机理中起到重要作用;菌毛在细菌和宿主细胞的相互作用中发挥重要毒力作用,文中预测的4种Ⅰ型菌毛蛋白fimA可特异性介导细菌对骨髓源树突状细胞的黏附和侵袭作用;6种外膜引入蛋白OmpD参与巨噬细胞和肠上皮细胞对细菌的识别,有研究表明OmpD突变菌株的毒力与野生菌株相比明显降低[13];ipaH9.8的编码产物作为效应分子,可以阻止血小板吸附到创面内皮下,还可调节宿主对沙门氏菌的反应,如抑制正常凝集作用的形成,并且促使水肿的形成[14~15]。同时,FC10727菌株中还发现了seh:SeHA_C3828基因,该基因能引起人类免疫系统疾病,如系统性红斑狼疮,这从基因层面证实个别沙门氏菌可引起系统性红斑狼疮。另外,我们发现,5个细菌侵袭上皮细胞基因(sew:SeSA_A1155、spq:SPAB_03584、sea:SeAg_B3007、spq:SPAB_03587、seg:SG2785)与志贺氏菌致病基因相似度高;3个上皮细胞信号转导基因(seh:SeHA_C1782、spq:SPAB_04085、sei:SPC_1568)与幽门螺杆菌感染基因相似度高,这从基因层面证实致病基因可在各菌种之间传递,从而使其获得致病性。
总的来讲,本研究对FC10727进行全基因组分析,预测致病基因和耐药基因等重要遗传信息,丰富了沙门氏菌的基因组数据库。在基因层面,本研究阐明了沙门氏菌可能存在的致病基因与致病机制,提示了沙门氏菌潜在的致病风险,为相关疾病的致病机理研究提供了方向,为相关疾病的疫苗开发和药物研发提供了分子基础。另外,该菌株多个耐药基因、多种耐药机制的发现,说明养殖户在家禽家畜的养殖期间可能存在滥用抗生素的情况,这客观上造成对沙门氏菌的多轮药物筛选,从而加快了高耐药性毒株的产生,因此,农业生产监管部门应加大抗生素使用危害的宣传力度,加强对养殖户抗生素使用的监管力度,从源头减少抗生素的使用,降低对抗生素的依赖。从食品安全角度分析,本研究在长沙市生鲜市场随机采集生鲜鸡肉即可检出沙门氏菌,说明生鲜鸡肉在养殖、屠宰、运输或销售等环节存在沙门氏菌污染的风险。虽然我国传统饮食习惯是不直接食用生鲜鸡肉,但普通消费者对于致病菌污染并无防范意识,这使致病菌存在较大程度的暴露风险,如:处理生鲜果蔬与生鲜畜禽肉的厨具混用;处理生食与熟食的厨具混用,这都会造成不可避免的交叉污染,导致普通消费者摄入沙门氏菌,引起腹泻、呕吐等疾病。本研究提示,消费者要提高生鲜畜禽肉中包括沙门氏菌及其他致病菌如志贺氏菌、致泻性大肠杆菌等的防范意识,不食用未充分加热的畜禽肉制品。同时,对于生鲜畜禽肉以及生鲜鸡蛋等沙门氏菌污染可能性较高的食品,目前我国并未制定致病菌限量标准,也未有关于生鲜食品中致病菌食品安全监督检验的相关报告,因此一定程度上存在食品安全监管盲区。本研究提示,食品安全监管部门应根据致病菌污染风险等级制定生鲜畜禽肉或其他生鲜食品的相关国家标准,并开展各地各类生鲜食品的致病菌监督抽检与风险评估工作,加强监督,倒逼生鲜畜禽肉和其他生鲜食品生产与销售企业,使其优化工艺、提高质量、防范风险,最终保障食品安全。
猜你喜欢
今日农业(2021年11期)2021-08-13 08:53:24
食品与机械(2019年1期)2019-03-30 01:14:36
食品研究与开发(2017年9期)2017-06-01 12:20:03
食品研究与开发(2017年5期)2017-04-11 08:20:36
公民与法治(2016年4期)2016-05-17 04:09:29
兽医导刊(2016年12期)2016-05-17 03:51:45
特产研究(2014年4期)2014-04-10 12:54:12
云南畜牧兽医(2014年4期)2014-02-28 21:25:30
遗传(2014年3期)2014-02-28 20:58:49
世界科学(2014年8期)2014-02-28 14:58:31
