
棘胸蛙的两性异形及生殖力
2021-08-03 06:09龙嘉航侯金亮胡亚洲向建国潘望城
野生动物学报 2021年3期
龙嘉航 侯金亮 胡亚洲 向建国* 潘望城
(1.湖南农业大学动物科学技术学院,长沙,410128;2.常德大北农饲料有限公司,常德,415400)
Darwin[1]最早对生物的两性异形进行了研究并认为性别差异性状与相关的求爱行为之间存在密切联系,从而为作为两性异形的主要进化机制——性选择理论奠定了基础。雌雄外部形态性状差异的形成归因于生育力选择(雌性较大)[2]或性选择(雄性较大)[3]。也有其他关注于副性征的两性异形的研究,如蛙的体色支持性选择或性生态位划分理论[4]。但两性异形产生的原因争议不断,还未统一定论。两栖类形态上的两性异形研究及其与雌性生殖力的关系受到广泛关注,如东北林蛙(Ranadybowskii)[5]的雌性成体大于雄性成体;棘腹蛙(Quasipaaboulengeri)[6]和饰纹姬蛙(Microhylafissipes)[7]雌性的生殖力均与个体大小有关。
棘胸蛙(Quasipaaspinosa)属两栖纲(Amphibia),无尾目(Anura),新蛙亚目(Neobatrachia),叉舌蛙科(Dicroglossidae),棘蛙属,其中大多数种确认分布在中国境内,属于易危物种[8]。近年内棘胸蛙自然资源日益减少[9-10]。目前国内外对棘胸蛙的两性异形的研究集中于体型与繁育之间的关系[11-12];Yu等[13]揭示了地理环境对棘胸蛙两性异形的影响。食物组成、能量配置等因素会在很大程度上影响个体形态大小[14]。研究两性异形特征与能量配置以及生殖力之间是否存在关联,对于棘胸蛙的保育工作具有一定的指导意义。
本研究通过对性成熟棘胸蛙数量性状的测量,比较了雌雄外表形态和肝脏、肺脏等主要器官组织的差异。旨在通过较全面地比较棘胸蛙雌雄间形态差异,探索差异形态性状与器官组织能量分配的关系,并寻找雌雄的体大小与生殖力的相关性,为解析导致棘胸蛙雌雄异形现象的原因提供理论参考。
1 材料与方法
样本于2020年2月在湖南石门采集(29°16′—30°8′ N,110°29′—111°33′ E),性成熟棘胸蛙共103只(雄性50只,雌性53只)。通过胸部是否存在刺疣以及解剖来判断性别。用数显游标卡尺统一测量动物体右侧头体长、头长等16个形态数量性状(精确到0.01 mm):各个形态数量性状测量标准参照文献[15]及图1。用电子天平称量体重(body mass,BM)以及皮重(skin mass,SKM)、心脏重(heart mass,HM)、肺脏重(lung mass,LUM)、脾脏重(spleen mass,SPM)、肾脏重(kidney mass,KM)、肝脏重(liver mass,LIM)、性腺重(gonad mass,GM)、脂肪重(fat mass,FM)、消化道重(DTM)10个重量数量性状(精确到0.01 g)。
雌性个体取出的卵巢测总重(F)后,随机选取其中一部分卵巢称重(FN1),对这部分的卵进行计数,记为N,最终计算雌性个体的怀卵量即生殖力(FN),其公式为:FN=N/FN1×F。
雄性个体睾丸由于无法计数则直接采用精巢重(g)作为代表生殖力的指标。重复测量3次取平均值。
计算两性异形程度公式[16]为:
两性异形程度=1-(体型较小性别的平均体长/体型较大性别的平均体长)。
2 数据处理
数据计算分析与制图采用软件Excel 2010和SPSS 23.0,结果采用“平均值±标准误”表示。用 Kolmogorov-Smirnov和Bartlett 分别检测所有数据的正态性和方差同质性,经检验,数据符合参数统计条件。采用t-检验检测体长和体重的两性差异;皮尔森相关性检验分析其他形态数量性状与头体长、重量数量性状与体重是否显著相关,显著相关数量性状的两性差异分别采用头体长和体重作为协变量进行协方差分析获得。采用皮尔森相关性检验分析雌雄群体形态数量性状与重量数量性状的相关性。线性回归分析雌性生殖力(怀卵量)与体大小、雄性生殖力(精巢重)与体大小是否具有相关性。显著性水平设置为α=0.05。
3 结果与分析
3.1 形态数量性状的两性异形
t-检验结果表明,湖南石门地区棘胸蛙雌雄个体的头体长间不存在显著差异。棘胸蛙雄性头体长平均值略大于雌性,两性异形指数为0.002。棘胸蛙头体长与其他15个形态数量性状均显著正相关(RHL=0.65,P<0.01;RHW=0.75,P<0.01;RSL=0.64,P<0.01;RINS=0.45,P<0.01;RIOS=0.32,P<0.01;RED=0.597,P<0.01;RUEW=0.42,P<0.01;RTD=0.46,P<0.01;RLAHL=0.59,P<0.01;RHAL=0.58,P<0.01;RLAW=0.36,P<0.01;RHLL=0.71,P<0.01;RTL=0.69,P<0.01;RTW=0.57,P<0.01;RFTL=0.63,P<0.01)。
协方差分析结果(表1)显示,在控制头体长的情况下,雌雄个体的头长、头宽、前臂及手长、手长、前臂宽、腿长、胫长和胫宽间差异达到显著水平(P<0.05);吻长、鼻间距、眼间距、眼径、上眼睑宽、鼓膜径、跗足长差异不具有统计学意义(P>0.05),即雄性个体的头长、头宽、前臂及手长、手长、前臂宽、腿长、胫长和胫宽显著高于雌性。

表1 成体棘胸蛙雌雄两性的形态数量性状差异
3.2 重量数量性状的两性差异
t-检验结果表明,湖南石门地区棘胸蛙雌雄个体间体重不存在显著差异。棘胸蛙体重与其他9个重量数量性状均显著正相关(RSKM=0.90,P<0.01;RHM=0.69,P<0.01;RLUM=0.53,P<0.01;RSPM=0.65,P<0.01;RGM=0.34,P<0.01;RKM=0.76,P<0.01;RLIM=0.71,P<0.01;RFM=0.58,P<0.01;RDTM=0.62,P<0.01)。
协方差分析结果(表2)表明,在控制体重的情况下,皮重、心脏重、肺脏重、性腺重、肾脏重、肝脏重、脂肪重差异具有统计学意义(P<0.05);脾脏重和消化道重差异未达到显著水平(P>0.05)。其中,雄性个体的皮重、心脏重、肺脏重和肾脏重显著高于雌性,雌性个体的性腺重、肝脏重和脂肪重明显高于雄性。雌雄间存在差异的重量数量性状与一些形态数量性状显著相关(图2),其中肝脏重在雌雄个体中均与16个形态性状相关性显著(P<0.05)。

表2 成体棘胸蛙雌雄两性的重量数量性状差异
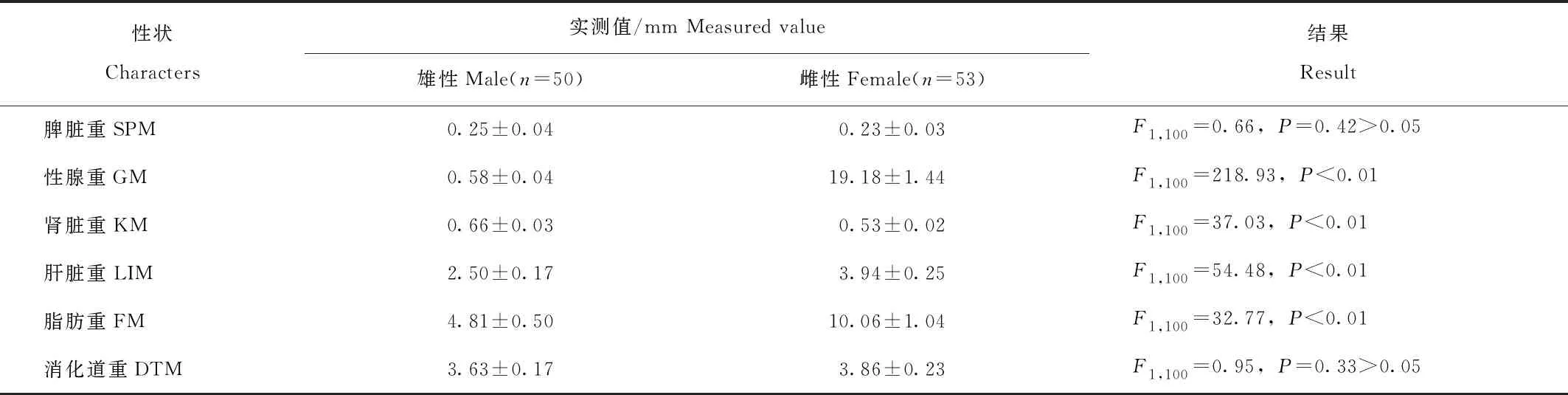
续表2
3.3 生殖力与体大小的线性回归关系
棘胸蛙雌性的生殖力随体重(F=19.54,P<0.01)和头体长(F=26.85,P<0.01)的增加而增加,(图3A-B)。一元线性回归分析表明:棘胸蛙雄性生殖力(精巢重,g)随体重(F=12.36,P<0.01)和头体长(F=6.79,P<0.05)的增加而增加(图3C-D)。
4 结论与讨论
4.1 形态数量性状的两性异形
两栖类中的两性异形可根据身体大小分为3种类型:雌雄同型[17]、雄性体型较大[18]、雌性体型较大[19]。大部分两栖类(无尾目中90%;有尾目Caudata中61%)都表现出雌性体型大于雄性[20]。本研究结果表明:棘胸蛙雄性与雌性个体的头体长和体重间无显著差异,两性异形表现在前臂及手长、手长、前臂宽、腿长、胫宽、胫长、头长和头宽这些局部形态特征上,且雄性大于雌性,所以湖南石门棘胸蛙是雄性体型较大类型。两性异形指数为0.002,与安徽棘胸蛙的0.06[21],闽西山区棘胸蛙0.07[18],湖南平江棘胸蛙的0.09[11]以及同属的双团棘胸蛙(Quasipaayunnanensis)的0.01[22]相比较小。棘胸蛙两性异形程度与当地最高温度和降水量呈正相关[13]。本研究的棘胸蛙两性异形程度与Yu等[13]研究中5个采集地的棘胸蛙相比最低,湖南石门的降水量和最高温度与5个采集地相比最低。也就是说,本研究的棘胸蛙与其他地区的棘胸蛙两性异形程度有异,可能是因为物种的两性异形程度与栖息地生境密切相关。物种两性异形程度的地理变异也存在于中国林蛙(Ranachensinensis):越往北方,降水量越大,环境温度越低,两性异形程度越明显,如内蒙古扎兰屯种群的两性异形指数高于河南郑州种群[23]。黑龙江林蛙(R.amurensis)两性异形差异程度与海拔呈正相关[24]。
本研究中,棘胸蛙雄性的前臂及手长、手长、前臂宽均大于雌性,说明前肢粗壮且长,便于与雌性在繁殖季节抱对,提高繁殖率;雄性的腿长、胫宽、胫长也大于雌性,便于跳跃活动,提高觅食效率。这与棘腹蛙以及浙江金华的棘胸蛙结果[6,13]一致。雄性的头长和头宽大于雌性,这与平江棘胸蛙未检测到头宽差异的结果[11]不同,头宽增大的同时,可利用食物的体积增大,从而获取更高的能量来增大个体;棘胸蛙在性选择时,雄性会强化有利于捕食和繁殖的形态特征。
性选择学说认为雄性中体型较大者同时携带了优质基因,雌性与其交配会增加繁殖成功率[25];抱对时,雄蛙增加个体大小能增大抱对的概率,但体型过大的雄蛙也会造成雌蛙的负担[26]。本研究雌雄个体间体大小不具有显著差异,但出现明显差异的各形态指标可以增强雄性竞争力支持性选择学说。雄性争夺资源能力与雌性对雄性的选择息息相关,进而影响繁殖成功率,雌雄形态特征差异也在此过程中强化。
4.2 重量数量性状的两性差异
重量数量性状不仅能反映个体发育和健康的状态,还与个体的生长性能及性状息息相关。影响脊椎动物个体大小的主要因素之一是摄食及消化能力,消化效率越强,其生长速度越快,个体越大[14]。此外,雌雄个体能量的分配差异也是导致雌雄个体大小出现差异的重要原因之一[27]。本研究中棘胸蛙雌性个体的肝脏重、性腺重和脂肪重明显高于雄性,其能量倾向于繁殖储存;肝脏重与形态性状相关性显著。肝脏是两栖类重要的消化器官,其发育状态影响各种营养物质的代谢,从而影响个体大小。雄性个体皮重、心脏重、肺脏重和肾脏重显著高于雌性,雄性的呼吸及循环排泄上能量占比更多。棘胸蛙雌雄间存在显著差异的重量数量性状与繁殖、呼吸及消化代谢等功能密切相关,这也可能是造成雌雄外形差异的原因之一。
4.3 生殖力
繁殖是动物生活史中重要的一个环节。棘胸蛙是1年多次产卵型,雌性生殖力主要依据怀卵量来判断大小[28]。成年雌体越大,繁殖输出越强,腹腔容纳量越大,从而提高繁殖适合度[6]。棘胸蛙雌性的生殖力(怀卵量)与头体长和体重呈正相关,此结果支持生殖力假说。棘腹蛙[6]、海陆蛙(Fejervaryacancrivora)[29]、中国林蛙[23]等相关研究也证实了雌蛙可以通过增加体大小实现繁殖率的提高。本研究棘胸蛙雄性个体生殖力(精巢重)与体大小呈正相关,雄性也有可能通过增加体大小来相应增加自身的精巢重来提高繁殖率。这也许是雌蛙倾向于选择体型更大的雄蛙与其交配[20]的一个重要原因。
对个体形态数量性状值综合分析可知,湖南石门棘胸蛙是雄性体型较大的两性异形类型。协方差分析两性差异结果表示,雄性会强化有利于捕食和繁殖的形态特征,故在性选择中雄性更占优势;棘胸蛙雌雄间存在显著差异的重量数量性状与繁殖、呼吸及消化代谢等功能密切相关,其中肝脏重在雌雄个体中均与16个形态性状相关性显著。雌性生殖力与体大小呈正相关的结果支持生殖力选择压力假说。雄性个体可通过增加体大小来提高生殖力,从而提高自身被雌性选择的竞争力。也就是说,在性选择、生殖力选择等压力作用下,棘胸蛙形态特征与功能性器官组织随性别分化而产生差异且相互之间相关性显著,促进其两性异形的进化。
猜你喜欢
儿童故事画报·自然探秘(2019年10期)2019-01-14
现代农业研究(2017年10期)2018-01-02
汽车与安全(2017年7期)2017-08-03
汽车与安全(2017年6期)2017-07-25
汽车与安全(2017年5期)2017-07-20
汽车与安全(2017年3期)2017-04-26
Coco薇(2015年12期)2015-12-10
现代畜牧科技(2015年1期)2015-10-21
中学生理科应试(2014年2期)2014-04-23
大学(2008年4期)2008-07-10
