
磷缓解油茶铝胁迫的叶片生理响应
2021-07-30 02:15渠心静张宸辉陈隆升
中南林业科技大学学报 2021年7期
渠心静,陈 铭,廖 皎,张宸辉,陈隆升,袁 军
(1.中南林业科技大学,经济林培育与保护省部共建教育部重点实验室,湖南 长沙 410004;2.湖南省林业科学院,湖南 长沙 410004)
铝是地壳中含量最丰富的金属元素,在土壤中主要以植物不可利用的铝氧化物和铝硅酸盐化合物存在[1],然而,在pH 值低于5.5 的酸性土壤中,难溶态铝大量溶出为Al3+,Al(OH)2+和Al(OH)2+等生物活性形态,对植物的生长产生毒害作用[1]。研究发现,铝胁迫首先抑制根系伸长,引起根系损伤,过多的Al3+还会抑制植株对钾、钙、镁等养分元素的吸收和转运,从而影响植物的正常代谢和细胞结构的稳定性[2]。长期的铝胁迫还会破坏叶绿体被膜,降低叶绿体光转化效率,抑制植物的光合作用和碳水化合物的合成[3]。此外,铝胁迫还会引起抗氧化应答异常,Mukhopadyay 等[3]就发现茶树在铝胁迫下其叶片超氧化物歧化酶(SOD)、抗坏血酸过氧化物酶(APX)、过氧化物酶(POD)和过氧化氢酶(CAT)活性被抑制,导致活性氧(ROS)累积。磷是植物生长所必需的大量元素,参与核酸、磷脂、ATP 等大分子物质的合成[4]。然而,酸性土壤中磷易被铁、铝等固定,导致土壤有效磷含量极低,对植株生长、碳水化合物合成、有机酸、氨基酸等代谢产生重要影响[5]。酸性土壤中同时存在的低磷和高铝胁迫严重影响作物的生产和增产[6],研究表明低磷和铝胁迫对植物生长的抑制呈正协同效应。因此,磷添加被认为是缓解铝胁迫的一个重要手段,Sun等[7]研究表明磷的添加能够降低植株的铝累积,提高磷的吸收并向地上部转运;于姣妲等[2]对杉木的研究表明,磷能够缓解铝胁迫对抗氧化酶活性的抑制,增强细胞自由基清除能力。
油茶Camellia oleifera是我国重要的木本油料树种,已有2 300年栽培历史。茶油因其不饱和脂肪酸含量高,被认为是健康优质的食用油,逐渐扩大油茶种植面积成为缓解粮油供应、改善居民膳食结构的有效方式[8-10]。目前,油茶在我国的种植面积超过400 万hm2,主要分布在长江以南的酸性红壤区[8],南方酸性红壤区高铝和低磷成为限制油茶生长和产量的主要因素之一,研究表明,磷的添加可以缓解铝胁迫对油茶植株生长的抑制,提高了植株对铝的忍耐能力。叶片是光合作用的主要器官,叶片的功能健康是植株生长和高产的关键,本研究设置不同的磷(+P 和–P)、铝(–Al 和+Al)组合处理,测定油茶‘华金’的生长、光合及抗氧化生理相关指标,以期为揭示磷缓解油茶铝胁迫机制提供参考资料。
1 材料与方法
1.1 供试材料及培养条件
本试验以油茶 ‘华金’Camellia oleifera‘Huajin’扦插苗为试验材料。选取健康粗壮的穗条,剪成8 cm 长插穗,每个穗条保留一芽一叶,扦插于干净的珍珠岩苗床上,进行常规管理。4 个月后,选取根系良好,长势一致的健壮扦插苗(高15.0±1.0 cm)移植于18 cm×22 cm 的塑料花盆中。生长介质为干净的珍珠岩和河沙混匀(体积比5∶2)。移植后的油茶 ‘华金’放置于小型温室中,白天平均温度为(28±3)℃,夜间平均温度为(22±3)℃,光源为日光,平均光强为847 μmol/(m2·s),平均湿度为47%。放入温室的油茶‘华金’幼苗在试验开始前进行3 周的预培养,5 天浇1 次营养液。营养液配方:0.713 mmol/L(NH4)2SO4、0.73 mmol/L NH4NO3、0.46 mmol/L K2SO4、0.50 mmol/L CaCl2、0.41 mmol/L MgSO4、0.032 mmol/L Fe-EDTA、0.046 mmol/L H3BO3、0.002 mmol/L CuSO4、0.09 mmol/L MnSO4、0.002 6 mmol/L Na2MoO4、0.009 1 mmol/L ZnSO4,溶液pH 值为4.2[11]。预培养3 周后,在原营养液基础上加入KH2PO4作为P 源,加入AlCl3作为Al源,进行不同的P、Al 组合处理。根据前期试验结果,本试验设置4 个组合处理:0.50 mmol/L P和0 mmol/L Al(+P–Al,对照);0.025 mmol/L P 和0 mmol/L Al (–P–Al);0.50 mmol/L P 和4.00 mmol/L Al (+P+Al);0.025 mmol/L P 和4.00 mmol/L Al (–P+Al)。–P 处理组的营养液加入相应量的KCl溶液以补齐因KH2PO4浓度降低引起的K+浓度降低。本试验用于浇灌油茶‘华金’幼苗的营养液澄清透明无任何沉淀。幼苗随机区分为4 个处理,每个处理20 株。P、Al 组合处理8 周后进行各项指标测定。
1.2 油茶‘华金’幼苗生长指标测定
处理结束后,记录每株幼苗叶片数,分别用卷尺和游标卡尺测定株高和地径。叶片和根系用根系扫描仪(Epson,Japan)扫描成图像后用WinRhizo Pro 2013 图像处理软件(Regent Instruments,Quebec,Canada)计算叶面积、根长、根表面积和根尖数。幼苗收获后,用纯水冲洗干净,置于60℃烘箱中烘干至质量不再变化,记录植株质量,记为植株干质量。
1.3 叶片磷和铝含量测定
油茶‘华金’叶片烘干后粉碎,用浓硫酸-双氧水法消解制备待测液,用全自动间断式化学分析仪(Westco Scientific Instruments,Rome,Italy)测定磷含量;适当稀释后用电感耦合等离子体质谱仪(iCAP Q,Thermo Fisher Scientific,Waltham,MA,USA)测定铝含量。
1.4 叶绿素含量的测定
采用王学奎等[12]的方法,叶片切成细条状,称取0.1 g 样品,加入10 mL 95%乙醇,避光提取24 h,提取液用分光光度计测定663 和645 nm 波长处的吸光值,并计算叶绿素a、叶绿素b、总叶绿素含量及叶绿素a/b。
1.5 光合及叶绿素荧光参数的测定
光合和叶绿素荧光参数选取完全展开的成熟叶片测定。每个处理随机选取5 株苗,每株苗选取3 片叶,在9:00—11:00 间用Li-6400XT 光合仪(LICOR,Lincoln,NE)测定净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)等光合参数。叶绿素荧光参数用LI-6400XT 光合仪荧光叶室测定,于夜间测定初始叶绿素荧光(Fo),然后施加饱和脉冲为7 200 μmol/(m2·s)的光强下0.8 s,测得暗适应下最大叶绿素荧光(Fm),根据Fv/Fm=(Fm-Fo)/Fm计算出Fv/Fm[13]。第2 天上午,测定PS II 有效光化学量子产量Fv′/Fm′、实际光化学量子效率(ΦPS II)、非光化学淬灭系数(NPQ)以及相对电子传递速率(ETR)等参数。
1.6 叶片可溶性糖、可溶性蛋白、游离氨基酸、脯氨酸含量测定
参照王学奎和黄见良的方法[12],可溶性糖含量采用蒽酮比色法测定;可溶性蛋白含量采用考马斯亮蓝G-250 法测定;游离氨基酸含量采用茚三酮显色法测定;游离脯氨酸含量采用磺基水杨酸提取,茚三酮显色法测定。
1.7 叶片PAL、PPO 活性及酚类物质含量测定
苯丙氨酸解氨酶(PAL)活性参照王学奎和黄见良的方法以苯丙氨酸为底物进行测定[12];多酚氧化酶(PPO)活性参照Hajiboland 等的方法以邻苯二酚为底物进行测定[14];可溶态酚类物质和细胞壁结合态酚类物质测定参照Heidarabadi 等的方法用福林酚试剂以没食子酸为标准测定[15]。
1.8 叶片H2O2、MDA 含量以及抗氧化酶活性测定
参照王学奎和黄见良的方法[12],SOD 采用氮蓝四唑法测定;CAT 活性以H2O2为底物分光光度法测定;POD 活性以愈创木酚为底物测定;APX活性采用Nakano 和Asada 的方法以抗坏血酸为底物用分光光度计测定[16];丙二醛(MDA)含量采用硫代巴比妥酸法测定。参照张志良等的方法[17],H2O2含量采用二甲酚橙法测定。
1.9 数据处理
采用Excel 2013 软件记录数据,采用SPSS 22.0 软件进行均值、标准差计算及方差分析,其中方差分析采用Duncan 法,采用Origin 8.0 软件作图。
2 结果与分析
2.1 磷对铝胁迫下油茶‘华金’幼苗生长的影响
磷铝交互处理对油茶‘华金’生长指标的测定结果见表1。与+P–Al 处理相比,低磷和铝胁迫(–P+Al)处理植株的生长明显受到抑制,植株干质量、株高、地径、叶片数、叶面积、根长、根表面积和根尖数分别降低了39.81%、38.33%、18.97%、51.61%、39.79%、42.75%、40.63% 和33.97%。高磷处理明显缓解了铝胁迫对植株生长的抑制,与–P+Al 处理相比,+P+Al 处理植株干重、株高、地径、叶片数和叶面积分别提高了36.94%、10.25%、13.58%、50.00%、47.79%、35.06%、31.02%和42.46%。

表1 磷对铝胁迫下油茶‘华金’幼苗生长指标的影响†Table 1 Effect of P on growth indices of C.oleifera ‘Huajin’ seedlings under Al stress
2.2 磷对铝胁迫下油茶‘华金’幼苗叶片磷和铝含量
铝胁迫显著提高了铝在油茶‘华金’叶片的累积(图1A),+P+Al 处理叶片铝含量显著低于–P+Al 处理,铝含量降低了20.12%;而+P–Al 和–P–Al 处理铝含量没有明显变化。高磷处理显著提高了油茶‘华金’叶片磷含量(图1B),其中+P+Al 处理磷含量显著高于+P–Al 处理,磷含量提高了16.19%。
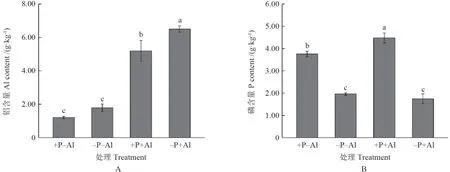
图1 磷和铝处理对油茶‘华金’叶片磷铝含量的影响Fig.1 Effect of P and Al on P and Al content in leaves of C.oleifera ‘Huajin’
2.4 磷对铝胁迫下油茶‘华金’叶片叶绿素含量的影响
磷铝交互处理对油茶‘华金’叶片叶绿素含量的影响见表2。低磷和铝胁迫均显著抑制了叶绿素含量,其中–P+Al 处理抑制作用最为明显。与+P–Al 处理相比,–P+Al 处理的叶绿素a、叶绿素b 和叶绿素含量降低了43.36%、50%和44.19%。高磷处理显著缓解了铝胁迫诱导的叶片叶绿素含量的降低。与–P+Al 处理相比,+P+Al 处理的叶绿素a、叶绿素b 和叶绿素含量提高了27.68%、26.67%和36.46%。与+P–Al 处理相比,低磷和铝胁迫处理增加了叶绿素a/b 的比值。
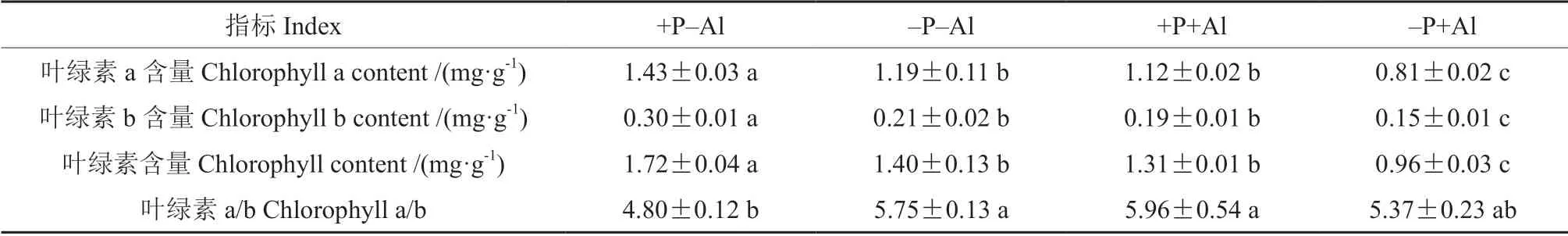
表2 磷对铝胁迫下油茶‘华金’叶片叶绿素含量的影响Table 2 Effect of P on chlorophyll content of C.oleifera ‘Huajin’ leaves under Al stress
2.3 磷对铝胁迫下油茶‘华金’幼苗叶片光合指标的影响
低磷和铝胁迫均显著抑制净光合速率Pn、气孔导度Gs和蒸腾速率Tr,与+P–Al 处理相比,–P+Al 处理的抑制作用最为明显;而高磷明显缓解了铝胁迫对Pn、Gs和Tr的抑制(图2A—C)。与+P–Al 处理相比,铝胁迫下油茶‘华金’叶片胞间CO2浓度Ci明显提高,其中–P+Al 处理Ci高于+P+Al 处理,但差异不显著(图2D)。
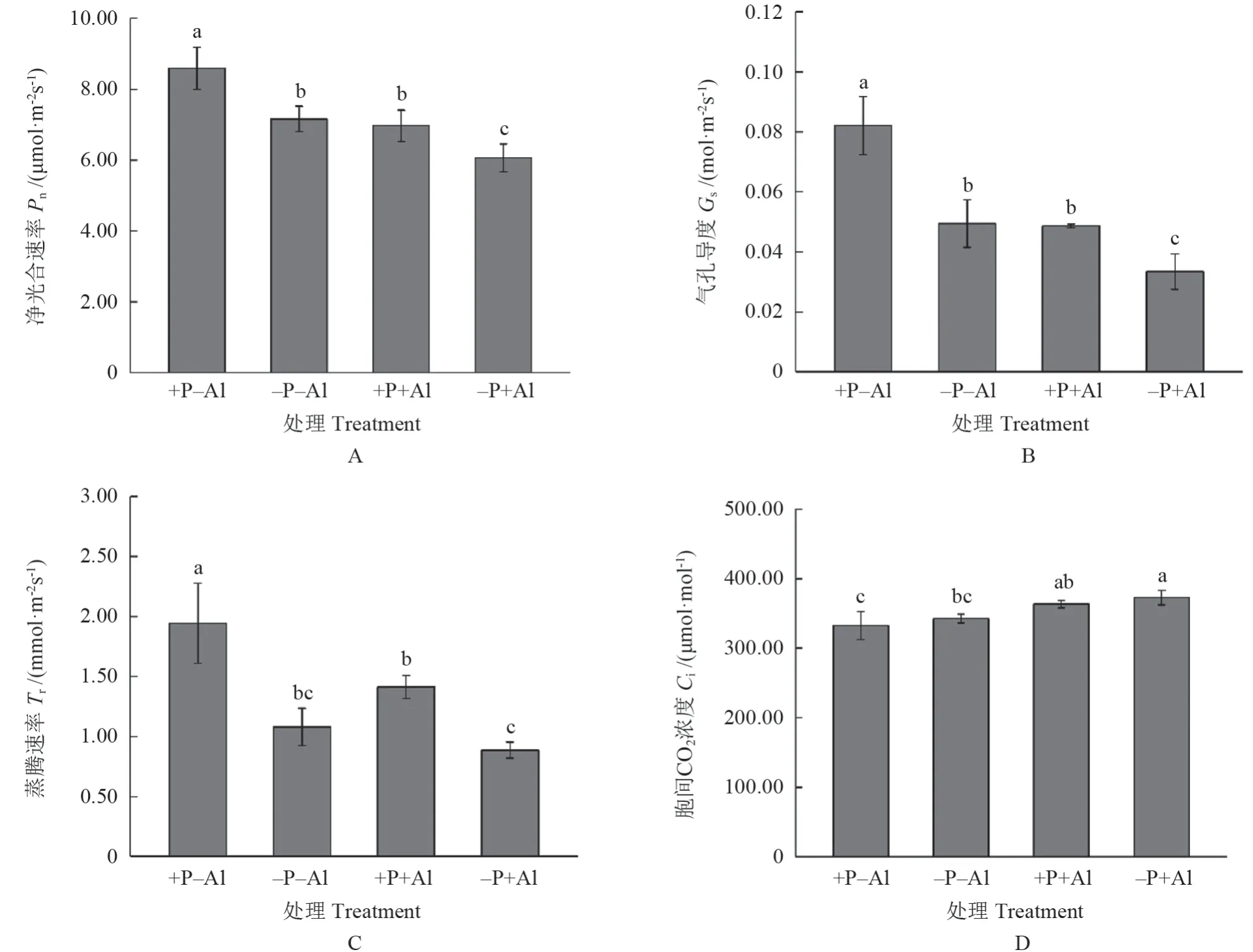
图2 磷对铝胁迫下油茶‘华金’叶片光合参数的影响Fig.2 Effect of P on photosynthesis parameters of C.oleifera ‘Huajin’ leaves under Al stress
2.5 磷对铝胁迫下对油茶‘华金’叶片叶绿素荧光参数的影响
Fv/Fm反映了PS Ⅱ最大光化学效率,当植物受到胁迫时,该指标会明显下降。–P+Al 处理Fv/Fm显著低于+P–Al 处理,而+P+Al 处理Fv/Fm值显著高于–P+Al 处理(表3);表明高磷处理提高了铝胁迫下叶片PS Ⅱ的光能转换效率。与+P–Al 处理相比,低磷和铝胁迫显著降低了PSII 有效光化学量子产量Fv′/Fm′值,磷的添加提高了铝胁迫下叶片Fv′/Fm′值,但差异不显著。–P+Al 处理的相对电子传递速率ETR、实际光化学量子效率ΦPSII 和非光化学淬灭系数NPQ 显著低于+P–Al处理,而+P+Al 处理的ETR 和ΦPSII 值显著高于–P+Al 处理,表明磷添加缓解了铝胁迫对ETR 和ΦPSII 的抑制。

表3 磷对铝胁迫下油茶‘华金’叶片叶绿素荧光参数的影响Table 3 Effect of P on chlorophyll fluorescence parameters of C.oleifera ‘Huajin’ leaves under Al stress
2.6 磷对铝胁迫下油茶‘华金’叶片可溶性糖、可溶性蛋白、游离氨基酸和游离脯氨酸含量的影响
低磷和铝胁迫均明显降低了叶片可溶性糖和可溶性蛋白含量(表4)。与+P–Al 处理相比,–P+Al 处理的可溶性糖和可溶性蛋白含量分别降低了19.29%和21.52%;而磷添加缓解了铝胁迫诱导的可溶性糖和可溶性蛋白含量的降低,与–P+Al处理相比,+P+Al 处理的可溶性糖和可溶性蛋白分别提高了18.46%和19.89%。相反,低磷和铝胁迫均显著提高了游离氨基酸含量和游离脯氨酸含量,其中以–P+Al 处理最高。
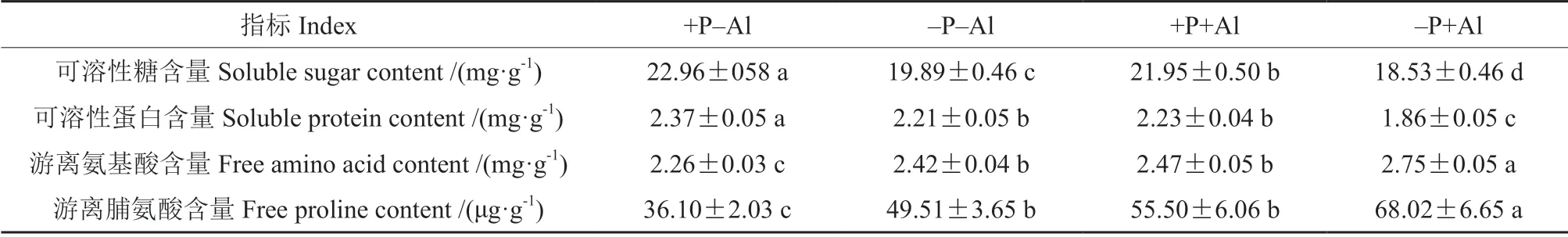
表4 磷对铝胁迫下油茶‘华金’叶片可溶性糖、可溶性蛋白、游离氨基酸和游离脯氨酸含量的影响Table 4 Effect of P on soluble sugar,soluble protein,free amino acid,and free proline content of C.oleifera ‘Huajin’leaves under Al stress
2.7 磷对铝胁迫下叶片PAL、PPO 活性及酚类物质含量的影响
–P+Al 处理PAL 活性较+P–Al处理提高了20.59%,而+P+Al 处理PAL 活性较–P+Al 处理降低了14.92%,但3 个处理间差异不显著(表5)。低磷和铝胁迫均显著提高了叶片PPO 活性,其中–P+Al 处理PPO 活性最高,是+P–Al 处理的3.22倍;而磷的添加显著降低了铝胁迫诱导的PPO 活性的增加。铝胁迫显著提高了油茶‘华金’叶片可溶性酚含量,而高磷处理并未显著影响可溶性酚含量。与+P–Al 处理相比,–P+Al 处理结合态酚含量显著提高,而磷的添加显著降低了铝胁迫诱导的结合态酚含量的增加。
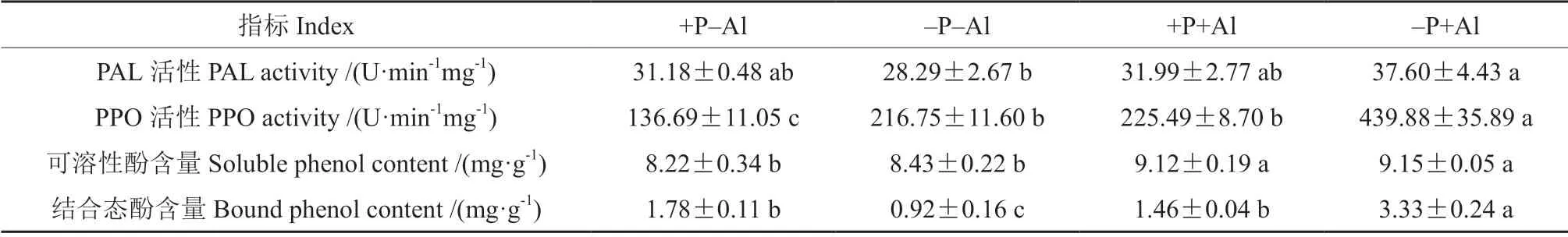
表5 磷对铝胁迫下油茶‘华金’叶片PAL、PPO 活性及酚类物质含量的影响Table 5 Effect of P on PAL and PPO activity and phenol content of C.oleifera ‘Huajin’ leaves under Al stress
2.8 磷对铝胁迫下油茶‘华金’叶片MDA、H2O2 含量及抗氧化酶活性的影响
与高磷处理相比,低磷处理显著抑制了SOD活性(图3A)。低磷和铝胁迫均显著抑制了POD和APX 活性(图3B—C),其中–P+Al 处理POD和APX 活性最低,磷的添加缓解了铝胁迫对POD和APX 活性的抑制。与–P+Al 处理相比,+P+Al处理POD 和APX 活性提高了40.01%和41.05%。CAT 活性以+P–Al 处理最高,其余处理均显著低于+P–Al 处理,且处理间无显著差异(图3D)。铝胁迫引起了油茶‘华金’叶片H2O2的累积,其中–P+Al 处理H2O2含量显著高于+P+Al 处理,H2O2含量提高了10.82%(图3E)。MDA 含量以+P–Al处理最低,低磷和铝胁迫均提高了MDA 含量,其中–P+Al 处理MDA 含量最高(图3F)。

图3 磷对铝胁迫下油茶‘华金’叶片MDA、H2O2 含量及抗氧化酶活性的影响Fig.3 Effect of P on MDA and H2O2 content and antioxidant enzyme activities of C.oleifera ‘Huajin’ leaves under Al stress
3 讨 论
强烈的淋溶作用导致酸性红壤区土壤活性铝大量溶出,而过量的活性铝会抑制植物的根系活力,降低养分吸收,阻碍叶片光合作用,破坏细胞结构,从而影响植株生长[18]。研究表明磷的添加能够提高植物的耐铝能力[2]。本研究结果显示磷添加缓解了铝胁迫对油茶 ‘华金’幼苗植株生长的抑制,降低了铝在叶片的累积(表1和图1A)。叶片是光合作用制造有机物的重要器官,铝在叶片的过量累积不仅会导致细胞壁僵化,进入原生质体的活性铝还会对细胞器产生毒害作用[19]。因此,磷添加降低了铝在叶片的累积,缓解了铝胁迫对油茶‘华金’叶片的毒害,降低了细胞壁的僵化度,而+P+Al 处理磷含量高于+P–Al 处理,可能是磷铝协同吸收并转运至叶片的结果[20]。
叶绿素在光合作用中起到捕获和转化能量的作用,将光能转化为化学能,是影响光合效率的关键因素之一。镁是叶绿素合成的关键元素,前人研究表明Al3+与Mg2+离子会产生竞争作用,铝胁迫会抑制根系对Mg2+的吸收,使叶片镁含量降低,抑制叶绿素的合成[21]。–P+Al 处理显著降低了叶绿素含量,而磷的添加提高了铝胁迫下叶片的叶绿素含量,可能是磷的添加提高了植株对镁等养分元素的吸收,促进了叶绿素的合成,降低了过量铝对叶绿素被膜的破坏。
逆境胁迫会导致植物光合能力下降,影响碳水化合物的累积[3]。本研究发现,–P+Al 处理油茶‘华金’叶片的净光合速率、气孔导度、蒸腾速率均明显受到抑制,表明铝胁迫引起根系水分横向运输受阻,导致植物体内水分缺乏,气孔导度和蒸腾速率下降[22-23]。影响净光合速率的因素分为气孔因素和非气孔因素,当气孔导度下降伴随胞间CO2浓度升高时,植物光合速率的抑制则是光合系统活性降低的非气孔因素引起的[24]。本研究中–P+Al 处理油茶‘华金’叶片胞间CO2浓度明显升高,表明长期的铝胁迫对光合的影响更多的是非气孔因素导致的,铝胁迫降低了光合系统活性,使得进入叶肉细胞的CO2不能被及时同化。磷的添加缓解了铝胁迫引起的净光合速率、气孔导度和蒸腾速率的下降,降低了胞间CO2浓度,表明磷可能是降低了铝胁迫引起来根系疏水物质的累积,促进了水分和养分向地上部的运输,减轻了铝胁迫对光合系统的破坏。此外,研究认为叶绿素含量与净光合速率呈正相关关系[13]。当叶绿素含量降低时,天线色素和反应中心色素捕获光能转化为电能的能力降低,从而导致光合效率下降[13]。
Fv/Fm值用来反映 PSⅡ复合物的光抑制程度,正常情况下,Fv/Fm一般在0.80~0.83 之间,该值随着胁迫程度增加而降低[24]。–P+Al 处理Fv/Fm值降低至0.727,表明该处理油茶‘华金’叶片严重光抑制使光合机构活性降低,PS Ⅱ的电子传递效率、光化学效率均显著降低,导致光能用于光化学电子传递的份额降低[18]。而+P+Al 处理Fv/Fm值为0.78,与–P+Al 处理相比光抑制程度降低,PS Ⅱ的电子传递效率、光化学效率均明显提高,提高了CO2同化能力,从而降低铝胁迫对光合的抑制。
蛋白质合成是一个高耗能的过程,降低蛋白质的合成是植物抵御逆境胁迫的重要方式,而铝胁迫能够诱导氨基酸的累积[25-26]。本研究发现–P+Al 处理提高了游离氨基酸和游离脯氨酸的累积,而+P+Al 处理降低了游离氨基酸和游离脯氨酸含量,提高了可溶性蛋白含量,增强了氮代谢。此外,+P+Al 处理叶片可溶性糖含量明显高于–P+Al 处理,表明光合效率的提高增强了碳代谢。
PAL 是催化苯丙氨酸转化为反式肉桂酸,进而合成酚类物质的关键酶[14]。多种逆境胁迫均能诱导PAL 活性增加,提高酚类物质合成[26-28]。研究表明铝胁迫可以诱导叶片可溶性酚含量提高,且通过Al 核磁共振检测出细胞液中的铝与儿茶酚形成化合物降低了活性铝对原生质体内部的毒害[23]。本研究表明,铝胁迫显著提高了油茶‘华金’叶片PAL 活性以及可溶性酚含量(表5),增强了活性铝螯合能力,抵御逆境胁迫。细胞壁上的结合态酚被认为是铝累积的潜在位点,细胞壁上铝的累积会诱导结合态酚含量的提高[22]。–P+Al 处理显著提高了细胞壁结合态酚含量,可能是过多的铝累积于细胞壁的结果。酚类物质是木质素合成的前体[22],低磷和铝胁迫诱导了细胞壁结合态酚含量的增加,促进了木质素的合成,而磷的添加降低了细胞壁结合态酚含量,降低了木质素合成速率,增加了细胞壁的灵动性。然而,PPO 活性的提高则会促进酚类物质氧化为醌类物质,进而诱导ROS的产生,对细胞产生毒害作用[29]。磷的添加降低了铝胁迫引起的油茶‘华金’叶片PPO 活性的提高,降低了有害物质的产生。
有研究表明,低磷和铝胁迫均会破坏抗氧化酶与ROS 间的平衡,引起细胞氧化伤害[2]。其作用机制是SOD 将超氧化物阴离子转化为H2O2和O2,POD、APX、CAT 将H2O2分解为H2O 和[30]。本研究表明,低磷和铝胁迫抑制了SOD、POD、CAT 和APX 活性,提高了H2O2的累积,引起了细胞氧化伤害,而磷的添加缓解了铝对SOD、POD 和APX 活性的抑制,降低了H2O2的累积,维持了细胞活性氧平衡,提高了植株耐铝能力[2]。本研究中+P+Al 处理MDA 含量显著低于–P+Al 处理,进一步表明磷添加诱导了抗氧化酶活性提高有效清除了活性氧,降低了膜脂过氧化伤害[31]。
本研究通过磷铝交互试验探究了铝毒胁迫及磷缓解铝毒胁迫的叶片光合和抗氧化防御作用机制,而根系也是铝胁迫的主要部位,磷在根系铝的累积吸附及抗氧化胁迫过程中的作用机制是下一步研究的内容。
4 结 论
磷和高铝处理显著抑制油茶‘华金’幼苗的生长,降低叶绿素合成,减弱光合效率,并引起氧化胁迫。而磷的添加不仅降低铝胁迫下油茶‘华金’叶片铝的含量,还提高了叶绿素含量,增加了叶肉细胞光捕获能力和CO2同化效率,提高了净光合速率,增强了碳氮代谢。同时,磷的供应还降低了PPO 活性,提高了抗氧化酶活性,降低了活性氧的累积。因此,磷添加可以增强油茶‘华金’对铝胁迫的忍耐能力,促进油茶‘华金’的生长发育。
猜你喜欢
南方农业(2022年12期)2022-11-23
农业工程学报(2022年1期)2022-03-25
河南科技(2022年1期)2022-03-11
农村百事通(2020年14期)2020-11-23
阅读(科学探秘)(2020年8期)2020-11-06
故事作文·低年级(2020年7期)2020-07-28
绿色科技(2019年2期)2019-05-21
女性天地(2016年10期)2017-04-25
源流(2017年1期)2017-03-07
农家顾问(2016年7期)2016-05-14
