
Molecular characterization and expression of the SiUCP2 gene in sea urchin Strongylocentrotus intermedius*
2021-07-29 09:12LingshuHANZijiaoQUANBingHANBeichenDINGXiaofangHUANGHengWANGYaqingCHANGJunDING
Lingshu HAN, Zijiao QUAN, Bing HAN, Beichen DING, Xiaofang HUANG,Heng WANG, Yaqing CHANG, Jun DING
Key Laboratory of Mariculture& Stock Enhancement in North China's Sea, Ministry of Agriculture and Rural Aff airs, Dalian Ocean University, Dalian 116023, China
Abstract Uncoupling protein 2 (UCP2) is a proton transporter located in the inner mitochondrial membrane, and inhibits the formation of adenosine triphosphate and reactive oxygen species by uncoupling oxidative phosphorylation. To provide a theoretical basis for the role of SiUCP2 in lipid metabolism, a 2 341-bp full-length cDNA of SiUCP2 from sea urchin Strongylocentrotus intermedius, which encodes 323 amino acids (predicted MW 36.11 kDa) was obtained, and the structure and function of the SiUCP2 gene and its expression at the mRNA and protein level were studied. SiUCP2 had high homology with UCP2 of other species. Expression of SiUCP2 was detected in the order of tube feet > gonads > coelomocytes > intestines.The expression level was the highest in prismatic larvae and lowest in the two-cell stage. Moreover, using in-situ hybridization, we found that SiUCP2 protein was expressed in the gonads and intestine. This study provided a theoretical basis for subsequent studies on the role of SiUCP2 and its regulatory mechanism in lipid metabolism, and for the improvement of gonad quality to obtain a higher economic value from sea urchins.
Keyword: Strongylocentrotus intermedius; SiUCP2; gene cloning; lipid metabolism; in-situ hybridization;western blot
1 INTRODUCTION
Adenosine triphosphate (ATP) represents one of the final compounds produced by animal organisms to store chemical energy converted from food. The generation of ATP is related to the energy requirements of the organism and is closely related to the metabolic rate. The metabolic rate quantifies the energy requirements of the organism, which is related to the size of the animal; therefore, the larger the body size is, the higher the metabolic rate is (McCGraham,1967). The metabolic rate is always in a state of fluctuation, which varies with the amount of energy used by the animal. When the animal is in a stage of reproduction or stress, the metabolic rate would change accordingly. Most organisms maintain a basal metabolic rate (BMR) before they consume additional energy for life activities. Therefore, organisms have evolved a feedback system called “energy balance”that responds to energy requirements, so that the metabolic rate can change with energy requirements(Campbell and Reece, 2005).
When starvation occurs, there lacks suffi cient energy substances to meet the needs of the juvenile perch (Percafluviatilis), thus the organism first uses glycogen stored in the body, and then uses fat as energy substance, which is oxidized and decomposed to maintain energy metabolism, and finally the organism uses protein (Mehner and Wieser, 1994).However, some aquatic animals use energy substances in a diff erent order during starvation. FishSciaenopsocellatus(Jiang et al., 2002) and juvenileEriocheirsinensis(Wen et al., 2002) prioritize the use of fats and carbohydrates, while a few species of fish, such asPagrusmajor(Zhang et al., 2000) mainly use protein as an energy source during starvation. Fatty acids (FAs) are an important source of energy for cells and are the primary raw material for triglyceride synthesis. Polyunsaturated fatty acids (PUFAs) play important functional roles in a variety of tissues, such as promoting nervous system development, mediating immunity, and promoting coagulation processes.Fatty acid derivatives and metabolites are involved in mediating multiple gene expression and enzymatic reactions, ion channel and membrane receptor activities, etc. FA metabolism in animals is a complex process that is regulated by several factors such as genetics, hormones, nutrition, and immunity. Factors such as inheritance influences fat deposition (Geri et al., 1990). Fat deposition depends on the eff ects of several enzymes during fat synthesis and catabolism,as well as the regulation of hormones involved in fat metabolism (Malavazos et al., 2007; Jiang et al.,2008; Kang et al., 2017). Researchers have undertaken several studies on fat deposition and its regulatory mechanism in livestock for many years, and found that candidate genes such asUCP1,UCP2,UCP3,LPL,PPARs,Leptin, andSREBPwere closely associated with fat metabolism and meat quality(Douaire et al., 1992; Taouis et al., 1998; Huang and Xie, 2004; Li et al., 2010).
Uncoupling proteins 2 (UCP2) belongs to UCPs family which is a class of proton transporters located in the inner mitochondrial membrane and are members of the mitochondrial anion carrier superfamily(MACF) (Fleury and Sanchis, 1999; Luévano-Martínez, 2012). Studies have shown thatUCP2is involved in multiple metabolic processes such as food intake (Coppola et al., 2007; Andrews et al., 2008),FA metabolism (Klingenberg and Echtay, 2001;Brookes et al., 2008), insulin secretion (Wang et al.,2010), and the immune response (Tagen et al., 2009;Emre and Nübel, 2010). Currently, theUCP2gene has been cloned from rat (Matsuda et al., 1997), pig(Werner et al., 1999), carp and zebrafish (Stuart et al.,1999), and silver carp (Liao et al., 2006), but the cloning ofUCP2from Echinoderms has not yet been reported.
Strongylocentrotusintermediuswas introduced to China from Japan by Dalian Ocean University in 1989 and is currently distributed in Liaoning and Shandong Peninsula. It is one of the main species of sea urchins cultured in China (Chang et al., 2004).The gonads of sea urchins are the only edible part of the organism. These organs contain essential nutrients such as lipids and PUFAs, which not only determine the nutritional value of sea urchins, but also ensure the normal growth and reproduction of the organisms during cultivation processes. Uneven feeding or excessive feeding often happens in the aquaculture of sea urchins. Moderate deposits of fat help the sea urchins maintain normal metabolic activities, but excessive deposits will not only aff ect the health of the sea urchins but also reduce the quality of the gonads (Zhou et al., 2008). Artificially cultured sea urchins are often exposed to starvation stress due to environmental and seasonal changes. During starvation, sea urchins will consume stored glycogen and fat to maintain their basal metabolism (Lares and Pomory, 1998).UCP2regulates lipid metabolism directly or indirectly, but the structure and expression ofUCP2and its role in lipid metabolism inS.intermediushave not been reported.
In this study, theSiUCP2gene was cloned fromS.intermediususing the rapid-amplification of cDNA ends (RACE) technique, and the expressions of this gene in diff erent tissues and at diff erent development stages and in diff erent starvation periods ofS.intermediuswere studied using quantitative realtime PCR (qRT-PCR). Gene expression in intestinal and gonadal tissues was also examined by in-situ hybridization. Finally, the expression ofSiUCP2recombinant protein in the intestine and gonads were examined using western blotting. This study aimed to investigate the structure and function of theSiUCP2gene fromS.intermediusand its expression at the mRNA and protein level to provide a theoretical basis for subsequent studies on the role ofSiUCP2and its regulatory mechanism the fatty acids synthesis ofS.intermedius. In the future, we will continue to pay attention toSiUCP2, study which fatty acids are produced bySiUCP2, clarify the mechanism ofSiUCP2in the synthesis of fatty acids, improve the content of fatty acids in sea urchins, and further improve the gonadal quality of sea urchins to obtain sea urchins with a higher economic value.
2 MATERIAL AND METHOD
Sea urchins and rabbits used in this study were farmed, and all experiments were conducted in accordance with the ethics committee of Dalian Ocean University and national guidelines. No endangered or protected species were involved in this test. The location of the training experiment does not require special permission.
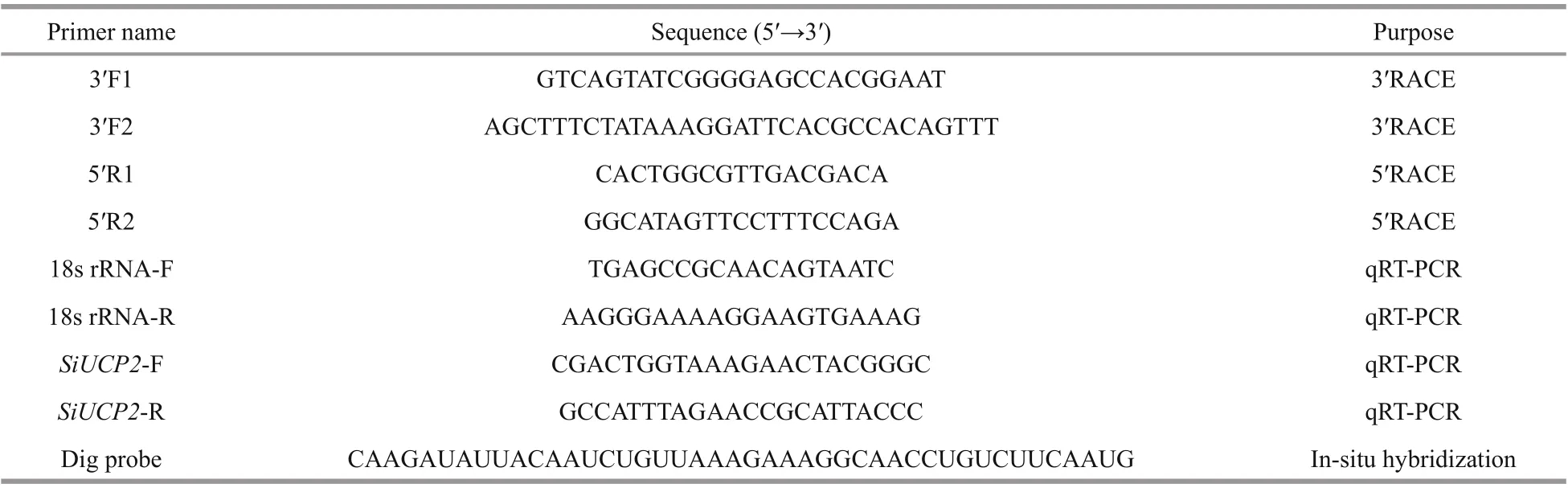
Table 1 The sequences of PCR primers used in this study
2.1 Animals, rearing conditions, and sample collection
One-year-oldS.intermediusadults used in this study were bred at Dalian Ocean University. Healthy sea urchins were homogeneous in size with test heights of 21.80±1.20 mm, shell lengths of 37.62±1.53 mm, and weights of 24.27±1.46 g.
The tube feet, coelomocytes, Aristotle’s lantern,stomachs, and gonads, intestines, were dissected from three sea urchins. To collect samples from diff erent developmental stages, we selected three females and three males and injected with 1-mL KCL (40 μL/g of body mass) to induce spawning. Unfertilized eggs were transferred to a beaker containing 1.5-L fresh sea water (salinity: 31) in density of 20-30 ind./mL in temperature of 19±0.5 °C. They were then dripped into semen (sperm-egg ratio was 1 000:1). Water was stirred to ensure sperm and eggs in good contact, and stayed for 15 min. After artificial fertilization, eggs were washed three times every 30 min to remove extra sperm and impurities. Morphological changes in larvae were observed under a microscope (Kelly et al., 2000). The fertilized eggs develop in three stages,early embryonic development, planktonic larva, and competent larvae, and four major developmental stages, the blastula, prismatic larval, long-arm larval,and juvenile sea urchin. All samples were frozen in liquid nitrogen immediately and stored at -80 °C.
A starvation experiment commenced in May 2019,in which 25 sea urchins were cultured in a 500-L tank in water temperature of 19 °C. Feces were observed every day since feeding was stopped. Once fecal excretion was significantly reduced (Li et al., 2004),five samples were taken as the control group, and the hunger experiment offi cially commenced. The intestines and gonads of five sea urchins were taken on Days 0, 7, 14, and 21 of starvation and rapidly frozen in liquid nitrogen, and stored at -80 °C.
2.2 Total RNA extraction and reverse transcription
Total RNA was extracted from diff erent tissues, at diff erent development stages and diff erent starvation periods ofS.intermedius, using an RNAprep Pure Kit for tissue (Tiangen Biotech, Beijing, China) according to the manufacturer’s protocol. Standard RNAs were used for cDNA preparation. The cDNA for qRT-PCR was prepared using a Prime Script™ RT Reagent Kit(TaKaRa, Japan). The reaction was performed in a total volume of 10 μL containing 2-μL 5×Primer script Buff er, 0.5-μL Primer script Enzyme MixI, 0.5-μL Oligo dT Primer, 0.5-μL Random Primer, 500-ng Total RNA and ddH2O. The PCR reaction conditions were 37 °C for 15 min and 85 °C for 5 s. All cDNA samples were diluted five times and stored at -20 °C before subsequent experiments.
2.3 Cloning the full-length cDNA of SiUCP2 using rapid amplification of cDNA ends (RACE) PCR
The partial cDNA sequence ofSiUCP2was obtained from our transcriptome assembly data(http://gsa.big.ac.cn/, accession number:PRJCA000276), and the gene-specific primers were designed by Primer 5.0 and obtained from Sangon Biotech, Shanghai as shown in Table 1. The cDNA for RACE was prepared using the M-MuLV First Strand cDNA Synthesis Kit (Sangon, Shanghai, China). The PCR reactions for 5′-RACE and 3′-RACE were conducted following the manufacturer’s instructions.Products from RACE were extracted and purified using a Quick Gel Extraction kit (Transgen Biotech,China). For specific experimental content, refer to kits instructions and Han et al. (2019). After the fulllength cDNA ofSiUCP2was amplified, it was ligated into the PEASY-1 vector and sequenced.
2.4 Bioinformatics analysis
The open reading frame (ORF) ofSiUCP2was analyzed using ORF Finder (https://www.ncbi.nlm.nih.gov/orffi nder/) and the deduced amino acid (AA)sequence was determined using the Expert Protein Analysis System (http://www.expasy.org/). The physical and chemical parameters of the deduced protein were computed by the ProtParam tool (http://web.expasy.org/protparam/). The functional domains of the SiUCP2 protein were predicted by InterPro(http://www.ebi.ac.uk/interpro/). Multiple sequence alignment of the deducedSiUCP2AA sequence was performed using DNAMAN 6.0. The phylogenetic tree was constructed using the neighbor-joining method in MEGA 7.0.
2.5 Real-time PCR analyses of SiUCP2 expression
The relative expression levels ofSiUCP2mRNA transcripts in diff erent tissues (coelomocytes, tube feet, Aristotle’s lantern, gonads, intestines and stomachs), were assessed at developmental stages(eggs (1-2 min after fertilization), two cells (40 min after fertilization), eight cells (2 h after fertilization),blastula (5-6 h after fertilization), gastrula (17-20 h after fertilization), prism larvae (24 h after fertilization), two-arm larvae (30-40 h after fertilization), eight-arm larvae (7-13 d after fertilization), juvenile sea urchins (30 d after fertilization)) and intestines and gonads of sea urchins that were fasted for 0, 7, 14, and 21 d were analyzed with quantitative real-time PCR (qRT-PCR) using the Applied Biosystems 7500 Real-time System (Applied Biosystems, Foster City, CA, USA) and SYBR Premix Ex Taq (SYBR PrimeScript™ RT-PCR kit II,TaKaRa, Japan) following the manufacturer’s instructions. The 18s rRNA gene was used as an internal reference gene. The amplification was performed in a total volume of 20 μL containing 2 μL of 1:5 diluted original cDNA, 10 μL of 2×SYBR Green Master mix (TaKaRa, Japan), 0.4 μL of ROX Reference Dye, 6 μL of PCR grade water and 0.8 μL(10 mmol/L) of each primer. The reaction conditions were followed by 40 cycles of 94 °C for 5 min, 94 °C for 30 s, 60 °C for 30 s, and 72 °C for 30 s. The final extension step was at 72 °C for 5 min. Three independent biological replicates and three technical repetitions of each group were carried out. The relative expression levels of the target gene were calculated by the 2-ΔΔCtmethod (Livak and Schmittgen,2001).
2.6 Statistical analysis
All data were expressed as the mean±SD (n=5).Pvalue was adjusted for multiple tests using a false discovery rate (Benjamini-Hochberg). Significant diff erences (P<0.05) for each variable were firstly detected using the one-way ANOVA test, followed by Tukey’s test and Tamhane’s T2 method. Statistical analysis was performed using SPSS software version 19.0 (IBM, Armonk, NY, USA).
2.7 In-situ hybridization
In-situ hybridization was performed using a DIG High Prime DNA Labeling and Detection Starter Kit I(Sangon). Probe primers were based on the full-length cDNA sequence ofSiUCP2. The intestines and gonads ofS.intermediuswere collected at the three stage of reproductive cycle the gonads and soaked in a 4%paraformaldehyde solution overnight. After the tissues were fixed, dehydrated with gradient alcohol, waxed,and embedded, they were cut into slices and baked in oven at 62 °C for 2 h. Paraffi n sections were dewaxed by placing in water and sequentially in xylene I for 15 min, xylene II for 15 min, anhydrous ethanol I for 5 min, anhydrous ethanol II for 5 min, 85% alcohol for 5 min, 75% alcohol for 5 min, and washed in diethylpyrocarbonate (DEPC). Slices were placed in the repair solution, boiled for 10-15 min, and cooled naturally. Proteinase K (20 μg/mL) was added over 30 min at 37 °C and washed with pure water, the prehybrid solution was added and incubated at 37 °C for 1 h and then decanted. The hybridization solution containing the probe was added at a concentration of 5 ng/μL, and the mixture was mixed overnight at 37 °C in an incubator. To wash away the hybridization solution, the sections were washed by 2×saline sodium citrate (SSC) at 37 °C for 10 min, washed by 1×SSC at 37 °C for 5 min, and finally washed by 0.5×SSC at room temperature for 10 min. Serum bovine serum albumin (BSA) was added and incubated at room temperature for 30 min. After pouring off the sealing solution, anti-digoxin-labeled alkaline phosphatase was added and incubated at 37 °C for 40 min. Then washed for 5 min with phosphate buff er saline (PBS)for four times. After the sections were slightly dried,the freshly prepared nitro-blue tetrazolium (NBT)color rendering solution was added, and the color rendering time was controlled under the microscope.The sections were washed with pure water to terminate the color rendering, and then nuclear solid red dye was added to stain the nucleus, which was washed with pure water after an appropriate degree of staining.Finally, neutral gum was used to seal the film and an optical microscope was used to observe the results.
2.8 Construction of prokaryotic expression plasmid and preparation of antibody
2.8.1 Construction of prokaryotic expression plasmid
The encoding region ofSiUCP2was amplified by a gene-specific primer with the enzyme cleavage site Nde I/Xho I. After sequencing, it was inserted into the expression vector pET22b (Sangon) to construct the recombinant plasmid pET22b-SiUCP2, which was expressed inEscherichiacoli.
2.8.2 Animal immunity
Four-month-old healthy female New Zealand white rabbits weighing 2.1 kg were used. For the first immunization, the protein antigen was emulsified with an equal volume of Freund’s complete adjuvant and injected (100 μL) into the lymph nodes of the hind legs. A second immunization was performed after 21 d. The protein antigen was emulsified with an equal volume of Freund’s incomplete adjuvant and injected (100 μL) into the lymph nodes of the hind legs. The third immunization was performed after 35 d. After 42 d, 1 mL of blood was collected from the ear vein and the antiserum titer was detected by enzyme-linked immunosorbent assay (ELISA).
2.8.3 Indirect ELISA test
The antigen was coated with 0.05-mol/L carbonate(pH=9.6) at 100 μL/well, incubated at 4 °C overnight,removed and washed for 3 min with 0.05% Tween-20(PBST) for three times. Blocking solution (150 μL of 5% skimmed milk) was added to each well and blocked at 37 °C for 60 min, and then washed for 3 min three times with 0.05% Tween-20 (PBST). The rabbit serum was diluted 1:1 000 and then incubated at 37 °C for 1 h, and washed three times with 0.05%Tween-20 (PBST) for 3 min per time. Horseradishlabeled enzyme goat anti-rabbit IgG (H+L) (Sangon)was diluted 1:8 000 and incubated at 37 °C for 45 min.The plate was washed five times with 0.05% Tween-20(PBST) and 100 μL/well of substrate solution (TMB)was added, the reaction was carried out for 15 min.Finally, 100 μL of 2-mol/L sulfuric acid was added to terminate the reaction. Optical density (OD) value was measured at a wavelength of 450 nm using a microplate reader (KeHua ST-360).
2.9 Extraction and expression of the SiUCP2 protein
About 20-mg gonads were taken and 200-L RIPA lysate was added (with 1-mmol/L PMSF, broadspectrum protease inhibitor, and phosphatase inhibitor). Low temperature and high speed homogenate for 3 times, 10 s each time, 30 s pause,then ice lysis for 2 h, and take out ice bath ultrasound once in the middle. At 4 °C, 12 000×ghigh speed centrifugation was performed, the supernatant was absorbed and centrifuged again, and the intermediate clarified sample was taken again until the obtained protein solution was clarified and transparent, which was temporarily stored at 4 °C. The protein concentration was determined. In this study, western blotting was used to detect SiUCP2 in the gonads ofS.intermedius, with an antibody that was diluted 1:2 000 and β-actin acted as a house-keeping gene.The resultant protein was isolated on a reduced sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE, 10% separating gel and 5% stacking gel, 40-μg protein extract was loaded into lanes for electrophoresis) and visualized after staining with Coomassie Brilliant Blue R-250.
3 RESULT
3.1 Analysis of SiUCP2 sequence
TheSiUCP2gene was obtained fromS.intermediusby RACE technology and the sequence was deposited in the GenBank database, under the accession number MN065154.SiUCP2was 2 341 bp in length and contained a 969-bp ORF encoding 323 AAs, a 232-bp 5′UTR, and a 1 140-bp 3′UTR containing a polyadenylation signal (AATAAAA).The molecular weight ofSiUCP2was 36.11 kDa,and the theoretical pI was 9.68. Analysis of the AA sequence predicted thatSiUCP2had no signal peptide and three characteristic features of carrier proteins in the inner mitochondrial membrane.Analysis of the AA hydrophilicity/hudrophobicity indicated thatSiUCP2had an aliphatic index of 87.24 and a grand average of hydropathicity(GRAVY) of 0.014, thusSiUCP2was a hydrophobic protein (Fig.1).
3.2 Multiple alignment and evolution analysis of SiUCP2 amino acid sequence
The AA sequence of SiUCP2 was aligned with UCP2 AA sequences from other species using DNAMAN 6.0 software. The results showed thatSiUCP2shared a high homology with allUCP2from vertebrates and invertebrates as shown in Table 2.
To reveal the phylogeny of theSiUCP2cloned in this study, a phylogenetic tree was constructed using the neighbor-joining method. The result in Fig.2 shows thatSiUCP2fromS.intermediusandUCP2fromS.purpuratuswere clustered into one branch,indicating that these two species were closely related.

Fig.1 Nucleic acid sequence of SiUCP2 in S. intermedius and its deduced amino acid sequence
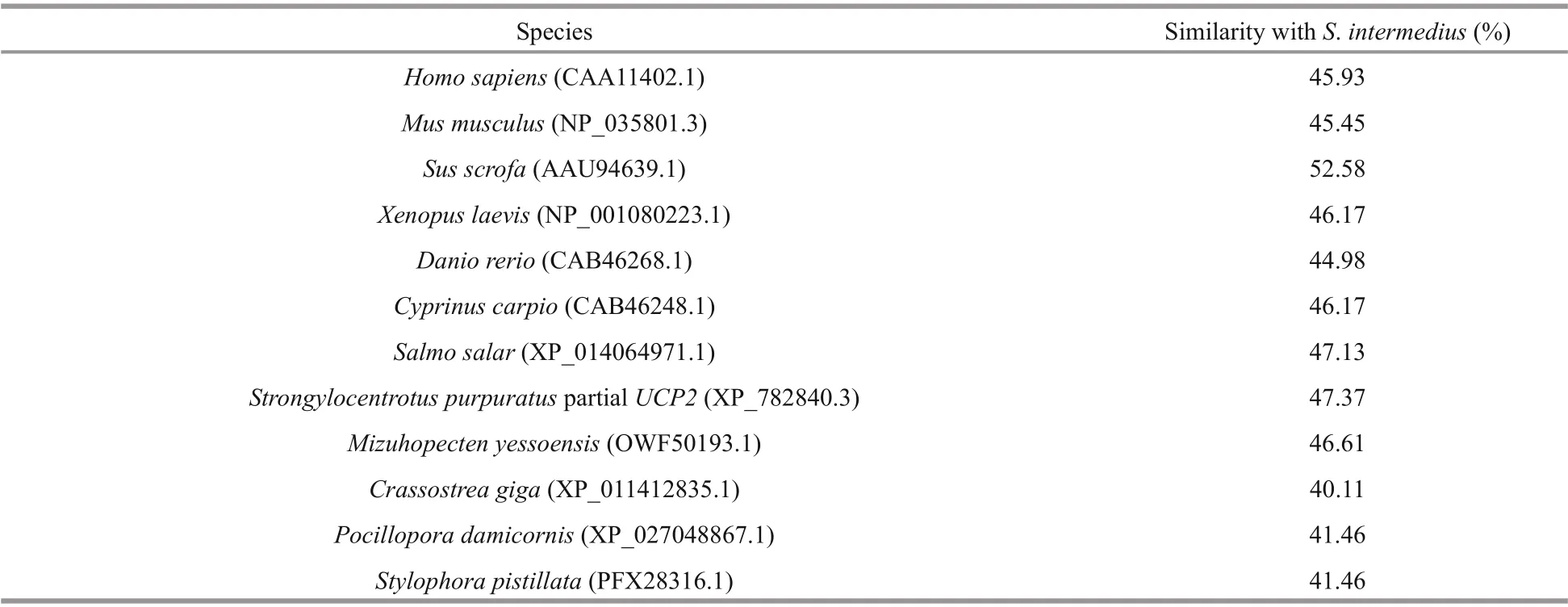
Table 2 Homologous comparisons of SiUCP2 with UCP2 in vertebrates and invertebrates
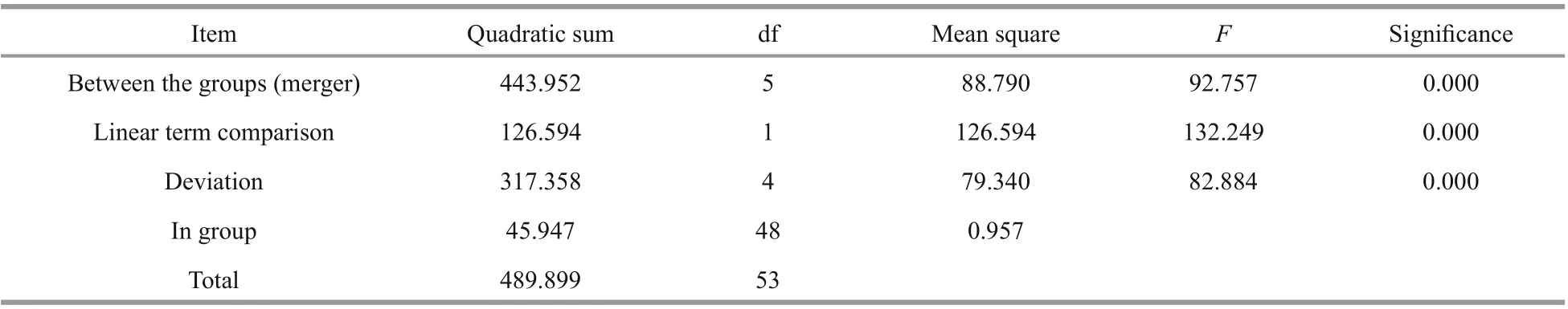
Table 3 One-way ANOVA of diff erent tissues
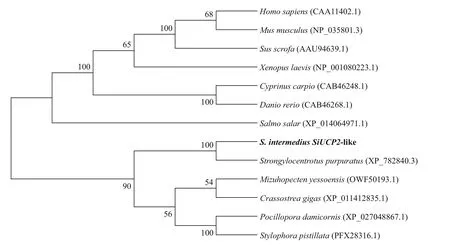
Fig.2 Consensus neighbor-joining tree based on the amino acid sequences of SiUCP2 genes from other species
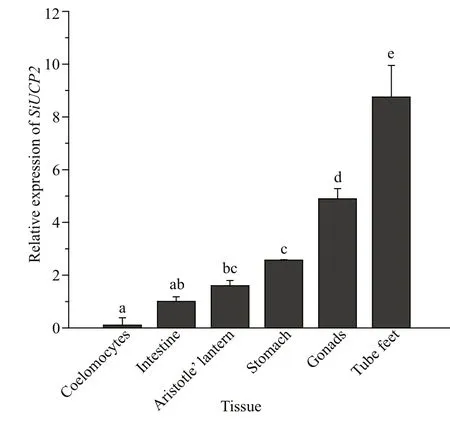
Fig.3 Relative expression levels of SiUCP2 in diff erent tissues
3.3 Relative expression of SiUCP2 in diff erent tissues
The tissue distribution ofSiUCP2mRNA inS.intermediuswas investigated by qRT-PCR. The results in Fig.3 show that theSiUCP2gene was expressed in all tissues examined. The data analysis results are shown in Table 3. Descriptive statistical analysis is shown in Supplementary Table S1. The relative expression level was the highest in the tube feet, followed by the gonads, and the relative expression level was the lowest in the coelomocytes.
3.4 Relative expression of SiUCP2 at diff erent development stages
The expression ofSiUCP2inS.intermediusat diff erent development stages, including fertilized eggs,two cells, eight cells, blastulae, gastrula, prism larvae,two-arm larvae, eight-arm larvae, and juvenile sea urchins, was determined to understand the role of theSiUCP2gene in diff erent development stages. As shown in Fig.4, The data analysis results are shown in Table 4. Descriptive statistical analysis is shown inSupplementary Table S2.SiUCP2was expressed at all nine development stages and the relative expression at the prism larval stage was the highest, while the expression at the 2-cell stage was significantly lower than at other development stages. During the whole development process, the overall expression ofSiUCP2showed a pattern of first increasing and then decreasing.
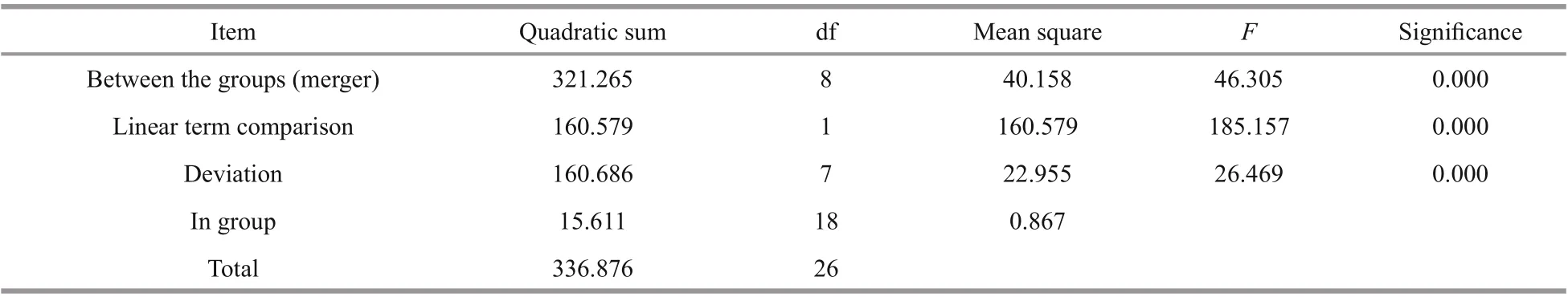
Table 4 One-way ANOVA of diff erent development stages
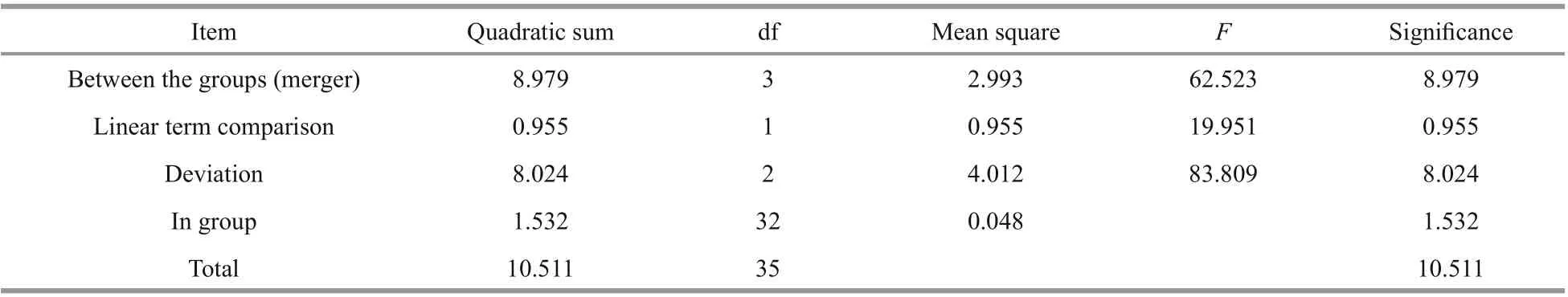
Table 5 One-way ANOVA of gonads at the diff erent starvation periods
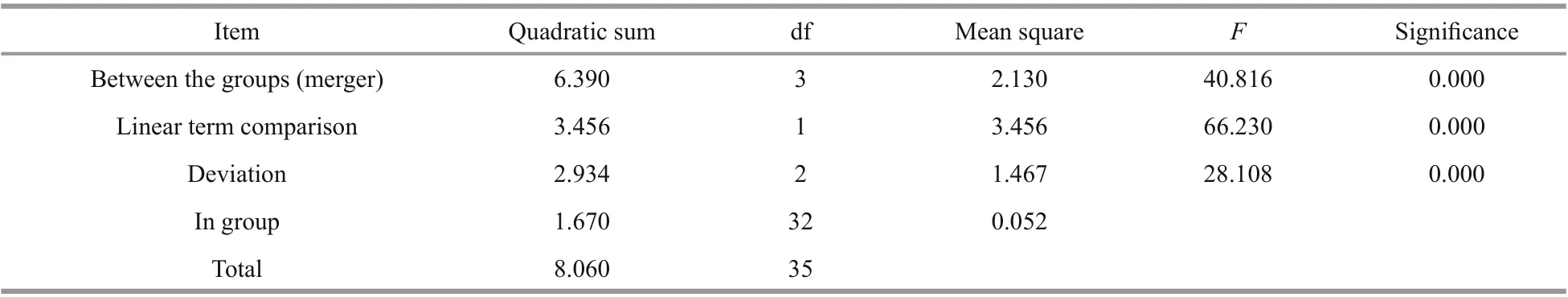
Table 6 One-way ANOVA ofintestines at the diff erent starvation periods
3.5 Expression of SiUCP2 during starvation
The relative expression ofSiUCP2in the intestines and gonads ofS.intermediusat diff erent starvation periods was determined using qRT-PCR to understand the expression and role ofSiUCP2during starvation.The data analysis results are shown in Tables 5-6.Descriptive statistical analysis is shown in Supplementary Tables S3-S4. As shown in Fig.5, the trends of diff erent periods of starvation onSiUCP2expression in the intestines and gonads ofS.intermediuswere similar. The relative expression levels ofSiUCP2in the intestines and gonads during starvation showed a pattern of first decreasing, then increasing and then decreasing again.
3.6 In-situ hybridization of SiUCP2
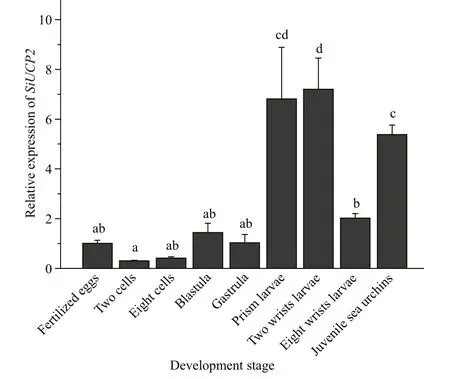
Fig.4 Relative expressions of SiUCP2 during the development of S. intermedius
In-situ hybridization ofSiUCP2in the intestines and gonads ofS.intermediuswas carried out. The results are shown in Fig.6. Scattered blue-violet positive signals in the gonads were observed under a microscope. The signals were present in nutritivephagocytes, while no positive signals were observed in the intestines, indicating thatSiUCP2was expressed in small amounts in the gonads and intestines.
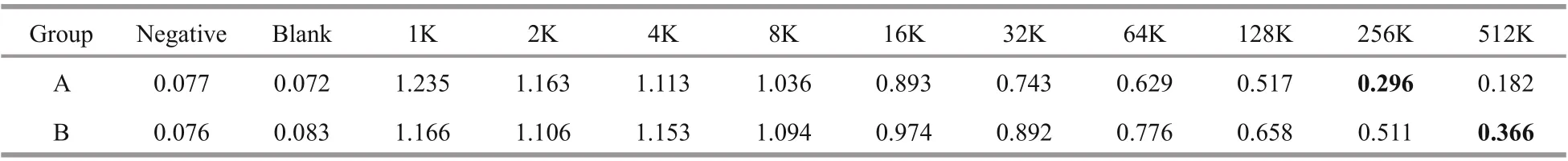
Table 7 ELISA assay data
3.7 Indirect ELISA results
The prepared antibody was subjected to indirect ELISA. The OD values were measured using a microplate reader at 450-nm wavelength. The results are presented in Table 7.
3.8 Expression of the SiUCP2 protein in the gonads of S. intermedius
To investigate the function of the SiUCP2 protein,total protein was extracted from the gonads of three sea urchins. The SiUCP2 protein was detected by western blotting using β-actin as an internal reference.Through the pre-experiment, we determined the destination band in the gonads, as shown in Fig.7. The results of SDS-PAGE in Fig.8 show that a clear band of about 36 kDa appeared in the gonads.
4 DISCUSSION
Sea urchins are considered delicacies in many countries. The gonads of sea urchins contain essential nutrients such as lipids and PUFAs, which not only determine the nutritional value of the sea urchins but also ensure the proton transporters, are located in the inner mitochondrial membrane. Free FAs provide necessary free carboxyl groups for UCPs, making proton transport possible or facilitating proton transport. Currently, the mechanisms of proton leaks caused by UCPs are still controversial (Garlid et al.,2000). UCPs that are present in the inner mitochondrial membrane are capable of disrupting the coupling between oxidation in the electron transport chain and adenosine triphosphate (ATP) synthesis, thus dissipating energy in the form of heat and possibly aff ecting metabolic effi ciency (Thompson and Kim,2004). The full-length cDNA of a gene is the basis for studying gene structure, function, and protein expression. TheSiUCP2gene ofS.intermediuswas characterized in this study. DNAMAN sequence alignment showed thatSiUCP2shared high homology withUCP2from other species and had similar domains; thereforeSiUCP2may also have similar biological functions to other homologs from diff erent species. Phylogenetic analysis showed thatSiUCP2fromS.intermediusandUCP2fromS.purpuratuswere clustered into one branch, indicating that these two species were closely related and distantly related to other vertebrates. Sequence similarity, conserved domains, and phylogenetic analysis provided evidence that the cloned gene wasSiUCP2cDNA.
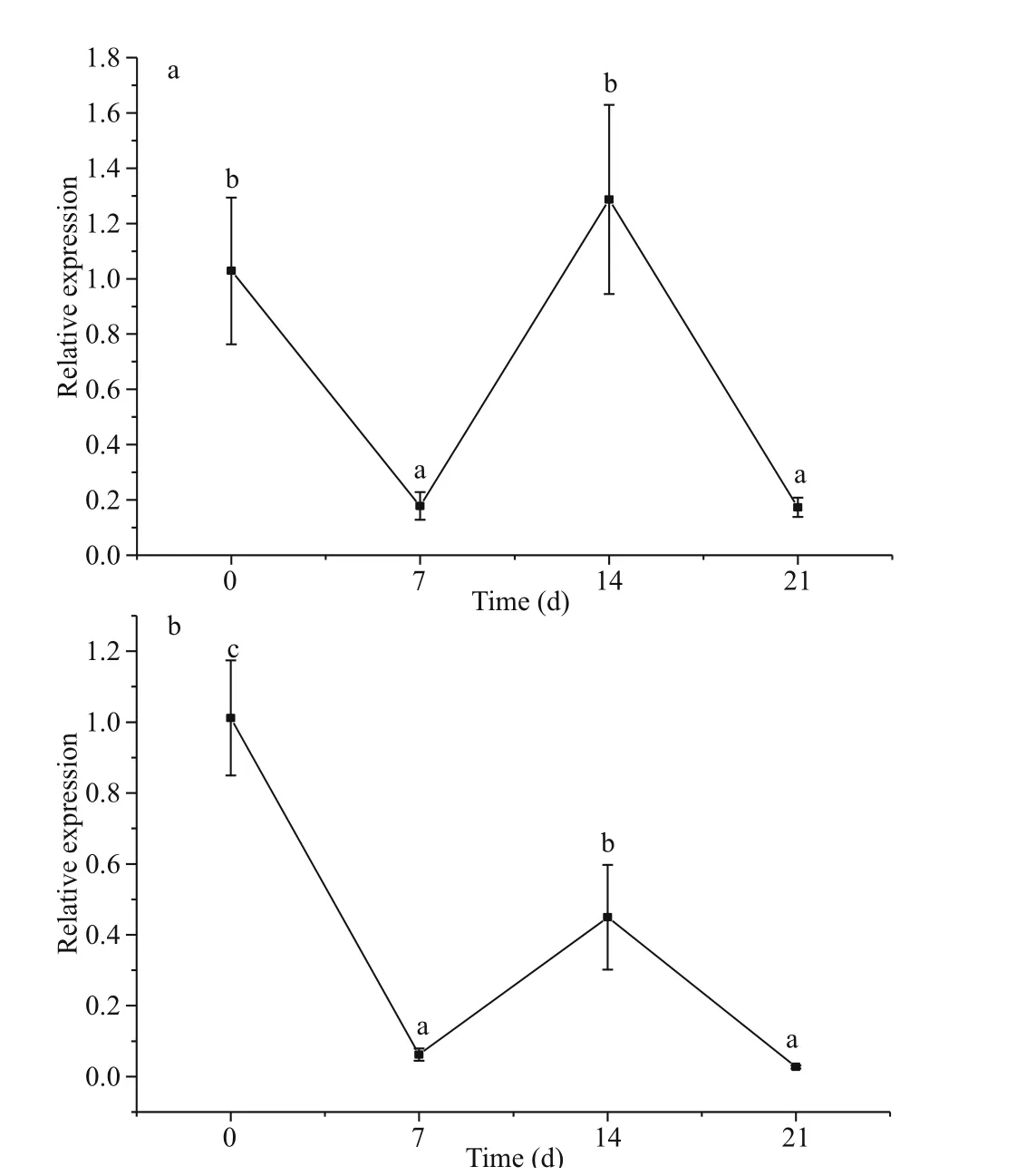
Fig.5 Eff ects of starvation on the relative expression of SiUCP2 gene in gonads (a) and intestines (b)
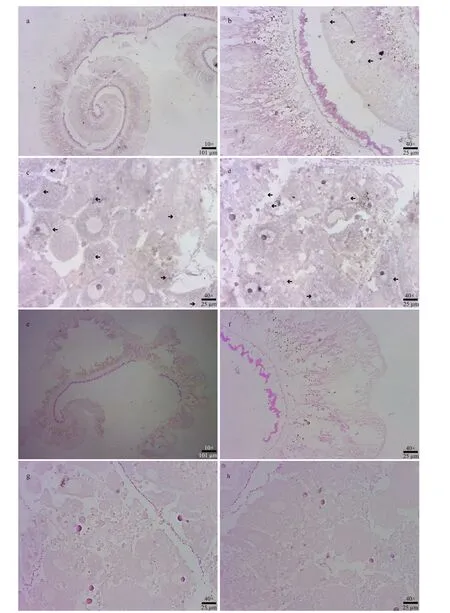
Fig.6 In-situ hybridization of the SiUCP2 gene in intestines and gonads
UCP2was highly expressed in the liver ofPagrusmajor, but almost undetectable in visceral mesenteric adipose tissue (Liang et al., 2003). The relative expression levels ofUCP2were high in the tissues ofliver, stomach, and eye ofSinipercachuatsi, but low in the spleen, intestine, and brain (Wen et al., 2002).The qRT-PCR analysis showed that theSiUCP2gene was expressed in all examined tissues ofS.intermedius, which was consistent with the study results in other species (Wen et al., 2002; Coulibaly et al., 2006; Liao et al., 2006). Sea urchin tube feet,which facilitate movement and adhesion, contain a large amount ofinorganic residues (45.5%), protein(6.4%), lipid (2.5%), and neutral sugar (1.2%) (Santos et al., 2009). The highest relative expression level ofSiUCP2in tube feet could be related to the energy consumption during movement and adhesion, which will be investigated in a follow-up study.
The high relative expression level ofSiUCP2in gonads may be due to fat metabolism in the gonads.The gonads of sea urchins store fat and are the main sites for fat metabolism. The decomposition of triglycerides produces a large amount of free FAs,resulting in a high expression level ofSiUCP2in the gonads, which was consistent with previous studies(Medvedev et al., 2002; Li et al., 2010). Sea urchin gonads contain nutritive phagocytes (NPs), which are versatile somatic cells that provide structural and nutritional microenvironments for germinal cells throughout sea urchin gametogenesis. In addition to mobilizing stored nutrients, NPs also continue to accumulate additional nutrients. NPs may also devour remaining eggs or sperm, to simply recover nutrients(Walker et al., 2013).SiUCP2may be expressed in NPs and participate in nutrient metabolism. Sea urchins feed mainly on large algae such as kelps,Wakame, and sea lettuces. Some algae contain toxins,and reactive oxygen species (ROS) are inevitably produced during the oxidation of toxins in sea urchins.Echtay et al. (2002) found that peroxides promotedUCP2gene expression, while a high expression ofUCP2eff ectively inhibited overproduction. The pharynx of the sea urchin is contained in Aristotles’lantern. The pharynx, stomach, and intestine constitute the digestive tract of the sea urchin. It is an important place for digestion and absorption in the body. The digestive tract has a certain fat content and immune functions, which prevent pathogenic and poison invasion. Algae food with certain toxins enter the gastrointestinal tract through the mouth to be digested,and the coelomic fluid of sea urchins is not only involved in transport, secretion, and buff ering but,more importantly, is involved in immune defense.Some studies have confirmed that amebocyte in sea urchin coelomic fluid play a role in phagocytosis to kill non-self-substances and mediate cytotoxic activity via the production of ROS (Ito et al., 1992).Therefore, low levels ofSiUCP2expression in the coelomocytes, Aristotles’ lantern and stomach may be related to ROS.
Lipids, including fat, phospholipids, and sterols,play a crucial role in individual life activities. The accumulation of total lipids, unsaturated FAs, and phospholipids in marine animals is essential for gonadal and early development (Palacios et al., 2007;Farhoudi et al., 2011). Previous studies have shown that the use of FAs in embryos varied with embryonic development stages (Yao and Zhao, 2006). During the early development of stingray embryos for example,high PUFA levels occur in the blastula stage, the gastrula stage and organogenesis, indicating that more PUFAs are needed during these periods to support the energy required for embryonic development (Yao et al., 2009). To understand the expression of theSiUCP2gene and its role in the development of embryos inS.intermedius, the relative expression levels ofSiUCP2gene at diff erent development stages ofS.intermediuswere determined using qRTPCR. The results show thatSiUCP2was expressed at all development stages in sea urchins. DecreasedSiUCP2expression in early embryonic development may be related to fat utilization as an energy source,as developing embryos use their lipid reserves to meet their energy needs (Coulibaly et al., 2006). During the gastrula stage in sea urchins, the intestine is diff erentiated and they begin to take in external nutrients and FAs are synthesized. Meanwhile, lipid metabolism is relatively enhanced, soSiUCP2gene expression was slightly increased (Zuo et al., 2016).When sea urchins develop into prism larvae, they begin to float. When they develop into two-arm larvae, the range of the sea urchin movement is expanded, and the digestive tract is formed in the larvae, so their food intake is increased, thus lipid and energy metabolism are enhanced (Chang et al., 2004).Therefore, the expression level ofSiUCP2gene was increased. When sea urchins undergo metamorphosis to the juvenile period, they became benthic, and their food changes from planktonic single-celled algae to benthic diatoms. Their food intake tends to be stable,so does the lipid metabolism and balanced. Therefore,the expression level of theSiUCP2gene was relatively decreased.
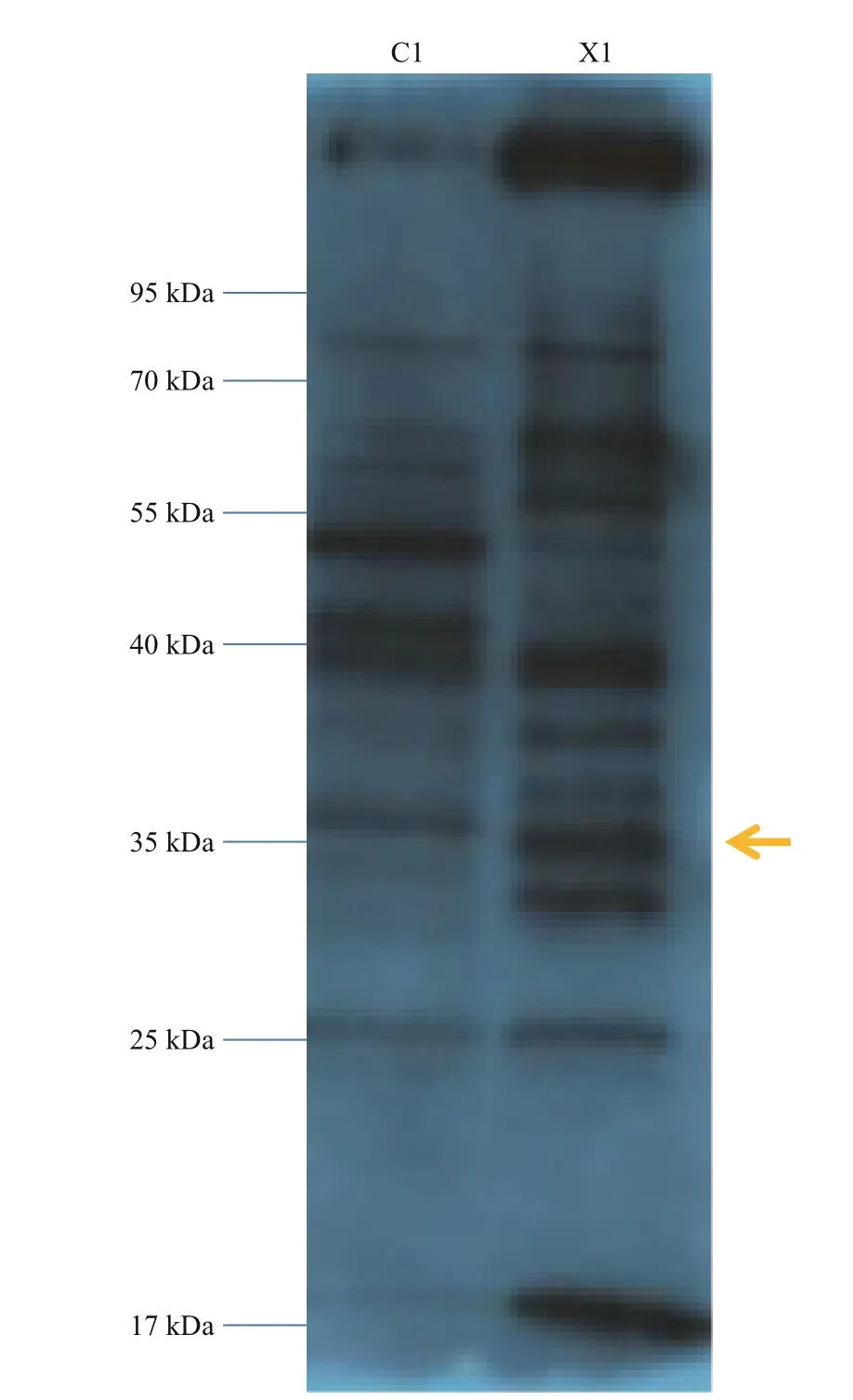
Fig.7 Pre-experiment of expression of the SiUCP2 protein in the intestine and gonad of S. intermedius
The intestines of sea urchins may be the site for FA synthesis (Han et al., 2019) and the gonads of sea urchins are the place where lipids are stored (Zuo et al., 2016). To understand clearly the role ofSiUCP2in lipid metabolism, the diff erence in expression ofSiUCP2in the intestines and gonads during starvation was analyzed using qRT-PCR. A previous study showed thatUCP2expression in the lungs and stomach of starved or polysaccharide-treated mice were increased (Pecqueur et al., 2001). In this study,as the starvation period was prolonged,SiUCP2expression showed a pattern of first decreasing, then increasing, and decreasing again in the intestine and gonads. The sea urchins used food storage as energy after starvation of 0-7 d, and their metabolism was slowed down. Therefore, the expression level ofSiUCP2was decreased after starvation for 0-7 d.When starvation was extended to 7-14 d, the expression levels ofSiUCP2were gradually increased in the intestine and gonads, and the expression level in the gonads was greatly increased, suggesting that sea urchins may maintain their life activities by consuming lipids and proteins in the gonads and intestine after 7-14 d of starvation, which further indicated thatSiUCP2played a role in lipid metabolism (Solanes et al., 2003; Tang et al., 2013),
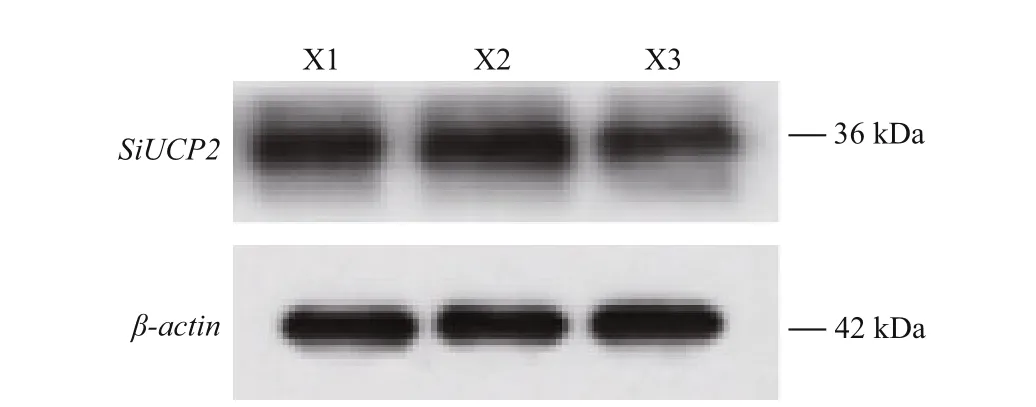
Fig.8 Expression of the SiUCP2 protein in gonads of S.intermedius ( n=3)
X1, X2, and X3 represent the gonads of the three sea urchins.andUCP2reduced ROS production that mediated oxidative damage in mitochondria (Echtay et al.,2002). After 14-21 d, the nutrients in sea urchins are exhausted thereforeSiUCP2expression gradually decreases.
Song (2002) used in-situ hybridization to detect the expression ofUCP2in mouse liver. The results show thatUCP2was not expressed in mouse hepatocytes but expressed in macrophages under normal healthy conditions. When treated with lipopolysaccharides(LPS),UCP2was expressed in mouse hepatocytes. In this study, DIG-labeled specificSiUCP2probes were designed to perform fluorescence in-situ hybridization in the intestines and gonads ofS.intermedius. The results show thatSiUCP2was expressed in the intestine and gonads, which is consistent with our qRT-PCR results. Owing to the scattered positive signals in the gonads, it was presumed thatSiUCP2was expressed in gonadal nutritive phagocytes, which is consistent with the findings of Larrouy et al. (1997).In the future, study of role ofSiUCP2in the gonads will be conducted.
In this study, the antibodies for SiUCP2 ofS.intermediuswere prepared successfully. Subsequently,we extracted the UCP2 protein and used Western blot to detect its expression in the intestines and gonads ofS.intermedius. The results of SDS-PAGE show that the SiUCP2 protein with a molecular weight of 36.11 kDa was successfully obtained. This size was very similar to the molecular weight of human UCP2(Pecqueur et al., 1999) and Chinese perch (Wen et al.,2002). Furthermore, the SiUCP2 protein was expressed in the gonads. The above-mentioned results are consistent with our in-situ hybridization results,which further demonstrated that UCP2 was closely related to lipid and energy metabolism.
Based on the results of this study, we speculated that theSiUCP2gene might be related to the fatty acids synthesis ofS.intermedius. In the future, we will continue to pay attention toSiUCP2, study which fatty acids are produced bySiUCP2, clarify the mechanism ofSiUCP2in the synthesis of fatty acids,improve the content of fatty acids in sea urchins, and further improve the gonadal quality of sea urchins.We hope to combine molecular technology with aquaculture in the future to select and breed more nutritious and economic value ofS.intermedius.
5 DATA AVAILABILITY STATEMENT
The data used to support the findings of this study are available from the corresponding author upon request.
6 AUTHOR CONTRIBUTION
Jun DING, Bing HAN, and Lingshu HAN contributed to the conception of the study. Lingshu HAN and Zijiao QUAN performed the experiments.Lingshu HAN, Zijiao QUAN, Xiaofang HUANG,and Beichen DING performed the data analyses.Lingshu HAN drafted the manuscript. Lingshu HAN,Jun DING, Bing HAN, and Heng WANG revised the manuscript. Jun DING and Yaqing CHANG approved the final version.
 Journal of Oceanology and Limnology2021年4期
Journal of Oceanology and Limnology2021年4期
- Journal of Oceanology and Limnology的其它文章
- Numerical study of the seasonal salinity budget of the upper ocean in the Bay of Bengal in 2014*
- Study on evaluation standard of uncertainty of design wave height calculation model*
- A fast, edge-preserving, distance-regularized model with bilateral filtering for oil spill segmentation of SAR images*
- A Gaussian process regression-based sea surface temperature interpolation algorithm*
- Climatology and seasonal variability of satellite-derived chlorophyll a around the Shandong Peninsula*
- Sources of sediment in tidal flats off Zhejiang coast, southeast China*