
Distribution and environmental impact factors of picophytoplankton in the East China Sea during spring*
2021-07-29 09:11FengWANGYuqiuWEIJiaqiYUECongcongGUOJunSUN2
Feng WANG, Yuqiu WEI, Jiaqi YUE , Congcong GUO , , Jun SUN2,,**
1 Institute of Marine Science and Technology, Shandong University, Qingdao 266237, China
2 College of Marine Science and Technology, China University of Geosciences(Wuhan), Wuhan 430074, China
3 Research Centre for Indian Ocean Ecosystem, Tianjin University of Science and Technology, Tianjin 300457, China
Abstract Marine picophytoplankton (Pico) as a major contributor to primary productivity in oligotrophic waters, play a very important role in marine material cycle and energy conversion, and their key role in the carbon cycle and global climate change is increasingly emphasized. To study the factors aff ecting the dynamic distribution of Synechococcus, Prochlorococcus, and picoeukaryotes in the East China Sea(ECS), a marginal sea of the Northwest Pacific, we investigated 27 stations in May 2017, and collected 148 samples of Pico and nutrients from the surface to the bottom. By means of flow cytometry, the abundance of Pico was measured, and then we estimated the carbon biomass and analyzed the distribution of Pico.Finally, combined with the ECS unique geographical situation and hydrological regime, we evaluated various factors aff ecting the Pico in the ECS. In Pico community, the picoeukaryotes cell abundance was between 0.49×10 2-1.44×10 4 cells/mL. Prochlorococcus ranged from 1.36×10 3-3.47×10 4 cells/mL and Synechococcus ranged from 0.69×10 3-1.15×10 5 cells/mL. Synechococcus was the most, both in abundance and in carbon biomass. Picoeukaryotes were the least in abundance, but has larger contribution to carbon biomass than Prochlorococcus. Water temperature, salinity, and stability of water column influenced Pico distribution. Picoeukaryotes were abundant in the shelf sea, whereas Synechococcus and Prochlorococcus were detected in the northeast of Taiwan, China. This study provided basic information for the study of Pico communities in the ECS and its adjacent marine ecosystem.
Keyword: picophytoplankton; distribution; East China Sea; physicochemical condition
1 INTRODUCTION
Phytoplankton is the dominant contributor to both primary production and biomass in the marine ecosystems (Metz et al., 2019). It is widely distributed in the epipelagic and photic zone in open oceanic regions (Li, 1994). In terms of the particle size,phytoplankton can be divided into three groups:microphytoplankton (20-200 μm), nanophytoplankton(2-20 μm) and picophytoplankton (Pico, 0.2-2 μm)(Takahashi and Bienfang, 1983; Hallegraeff and Jeff rey, 1984). Cells in picosized are greatly adapted to phytoplankton life, because they sink extremely slowly and are more eff ective at taking up nutrients and absorbing radiant energy than larger cells (Fogg,1986). In addition, the small size makes them have higher nutrient utilization and carbon fixation ability(Guo et al., 2014). On the whole, owing to their high concentrations, wide distribution, and major contribution to nutrients and carbon cycling, Pico have been considered to play a critical role in marine ecosystems (Jiao et al., 2002) and have large impact on the global biogeochemical circulation (Wei et al.,2019b). Moreover, they have great significance for the formalization of the concept of the microbial loop(Azam et al., 1983). At least three types of Pico can be identified in the collection samples:Synechococcus(Syn),Prochlorococcus(Pro), and picoeukaryotes(PEuks) (Marie et al., 1997). At present, there are three common ways to quantify and identify the taxa of Pico, including flow cytometry (FCM) (Marie et al., 1997), electron microscopy (SEM) (Guillou et al.,1999), and molecular approaches (Wood, 1988). FCM is an instrument that used to determine the properties of single cells or other particles in suspension(Bohmer, 1987). It can reflect the cells size, shape structure, pigment types, and internal structure shown in the Pico autofluorescence. It can quickly complete quantitative and qualitative research on single-cell Pico, and even sort the cells. FCM analysis has provided significant new methodological capabilities in oceanography.
SynandProare the most important primary producers in many oceanic environments among these autotrophic Pico (Chiang et al., 2002).Syngroup is known to make a relatively large contribution to photosynthetic phytoplankton in nearshore or coastal waters (Jochem, 1988). In addition,Synis one of the most important food sources for microzooplankton because ofits rapid biomass cycle and high-energy conversion effi ciency in the ocean microbial food web.Prois the smallest cyanobacterial cells in oceans, it is usually dominant in stratified tropical oceanic environments, single and widely distributed, it has many adaptation mechanisms, for example, reducing cellular nutrient requirements and facilitating growth in the nutrient-poor environment(Read et al., 2017).Procontains a unique photosynthetic pigment-divinyl chlorophyll, which can capture extremely weak light (Chisholm et al.,1992). The discovery ofProhas displacedSyn, andProbecomes the most abundant marine autotrophs(Jiao et al., 2005). Although concentrations of the PEuks are much smaller than that ofSynandPro, it is widely distributed in the Atlantic Ocean (Dı́ez et al.,2001), Pactific Ocean (Short and Suttle, 2003) and the Mediterranean Sea (Dı́ez et al., 2001). Furthermore,PEuks has turned out to be very diverse with Pico taxa distributed widely across several branches of the eukaryotic tree oflife (Vaulot et al., 2008; Kuwata et al., 2018). The PEuks is more suited for effi cient photosynthesis in the dim light at the bottom of the euphotic zone (Wood, 1985).
The East China Sea (ECS) is not only the largest marginal sea of the western North Pacific but also the most productive sea in China (Chiang et al., 2002;Gong and Liu, 2003; Gong et al., 2003). The entire sea area is on a continental shelf, and the average depth is about 349 m. The ECS has formed a huge low-salt water mass, which has become a eutrophic area in China off shore waters, with the salinity above 34. There are two major sources of nutrients: one is persistent upwelling located in the southern ECS, the other is Changjiang (Yangtze) River runoff (Chiang et al., 2002). In addition, the ECS was aff ected by oligotrophic water from the Kuroshio (Jiao et al.,2002). It thus ideally suits for ecological studies.Compared with the Bohai and the Yellow Sea, the ECS has higher temperature and salinity. Typically,the ECS has a tropical and subtropical climate. Such physicochemical and hydrographical conditions are conducive to the growth of Pico and complicate their distribution (Chen, 2000). The intrusion of the coastal current, Kuroshio into the continental shelf obviously caused variations in the distribution of temperature,salinity, nutrients and so on, and then consequently resulted in the variation of Pico community structure(Jiao et al., 2005).
Previous studies about Pico groups were obtained by epifluorescence microscopy (Vaulot and Ning,1988), which was not as effi cient, sensitive, and precise as FCM. Therefore, more eff orts are needed for a better understanding of the dynamics and regulation mechanisms of Pico in such ecosystems. The purpose of this study was to investigate the structure and dynamics of these three groups as well as relevant diverse hydrographical and physicochemical conditions factors, in an attempt to understand the factors that aff ecting the distribution of these Pico in the ECS.
2 MATERIAL AND METHOD
2.1 Study area
These data were obtained on the R/VDongfanghong 2cruise in May 2017. Our study area extended from 25°N to 30°N and from 121°E to 126°E in the ECS,and 27 stations were investigated. All stations were selected from the Changjiang River estuary to the north of Taiwan, China, and were assigned to transect A, B, and C (Fig.1). The Changjiang River estuary was one of the important estuaries in China. The surrounding area had developed economy and prosperous industry. At the same time, it also had important fishing grounds and aquaculture areas. This water area was aff ected by water masses such as the Changjiang River diluted water, coastal currents,Taiwan warm current, and Kuroshio. The ecological environment was complex and changeable.

Fig.1 The East China Sea sampling stations (a) and the schematic diagram of the circulation system (b)
2.2 Sample collection
At each station, seawater samples were collected from 4 to 7 diff erent depths within the upper 100-m water column using 12-L Niskin bottles equipped with a SeaBird CTD (SBE 9/11 plus). Physiochemical parameters such as salinity, temperature, and depth were also recorded in situ at the same time by the CTD sensors. For nutrients analysis, 250-mL seawater was collected and filtered through 0.45-μm cellulose acetate membrane filters and then immediately refrigerated at -20 °C for further analysis. Samples for FCM analysis were fixed with paraformaldehyde(final concentration: 1%), and then left to stand for 10-15 min in the dark at room temperature, so as not to loss of resolution and changes in cell counting due to fixation or freezing, then immediately freezetrapped in liquid nitrogen until received at the laboratory (Marie et al., 1997).
2.3 Sample analysis
Analyzed for the nutrients concentrations including ammonium, phosphate, silicate, nitrate, and nitrite were performed by a Technicon AA3 Auto-Analyzer(Bran+Luebbe) according to the classical colorimetric methods. The nitrate and nitrite were measured using cadmium-copper column reduction and naphthylethylenediamine photometric method,respectively. The determination of silicate and phosphate was by silicon molybdenum blue spectrophotometry and phosphor molybdenum blue spectrophotometry, respectively (Dai et al., 2008).
FCM (Becton-Dickinson Accuri C6) was used to quantify the abundance of three Pico groups. FCM was considered very successful in the study of Pico(Troussellier et al., 1995). According to the spontaneous fluorescence characteristics of Pico,FCM can eff ectively detect and enumerate the Pico groups without stain fluorescein (Yentsch et al.,1983). FCM equipped with a laser emitting at 488 nm,the phycoerythrin and chlorophyll contained inSyncould be excited orange fluorescence (FL2) and red fluorescence (FL3) at 560-590 and 685 nm,respectively.Proand PEuks contained chlorophyll only, which can emit red fluorescence only, thusSynthat can emit orange fluorescence could be distinguished out (Phinney and Cucci, 1989). At the same time, forward scatter light (FSC) was used to characterize cell size to distinguishProand PEuks.Prowas a small cell with weakly fluorescent, while PEuks was the group with larger cell size and stronger fluorescent (Wei et al., 2019a). We set the trigger threshold (~600) below the lowest scatter signals from surfaceProcells. It should be noted that in order to prevent the channel of FCM from being blocked by cells or impurities with larger particle size, we need to unclog and backflush the instrument when analyzing samples (Wei et al., 2019b). We collected 1.5 mL of seawater, and added 0.5 mL of paraformaldehyde,then only 196 μL of samples was used for analysis at a rate of 66 μL/min running for 3 min. Sample injection into the flow chamber is volumetric,allowing direct cell counts per μL to be obtained(Troussellier et al., 1995).
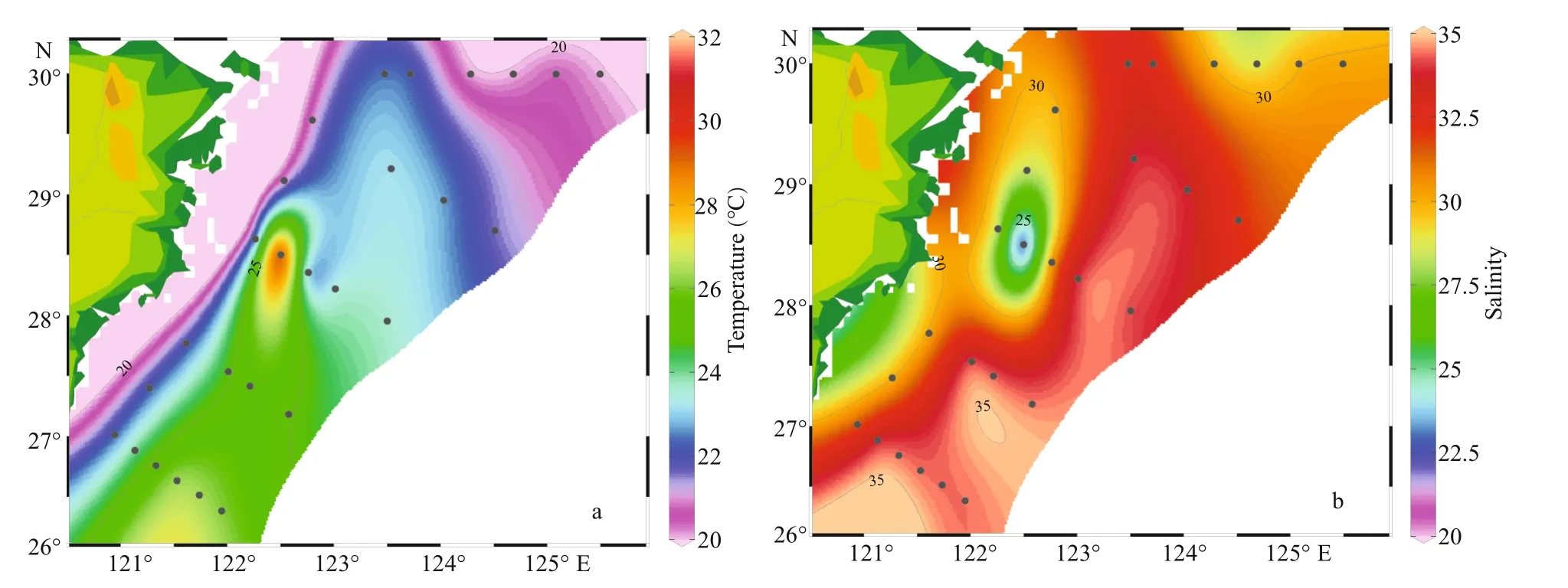
Fig.2 Surface distributions of temperature (°C) (a) and salinity (b)

2.4 Estimation of Pico carbon biomass
At present, Pico biomass was usually estimated according to the carbon conversion coeffi cient of various groups. The carbon conversion coeffi cient commonly used in the international was adopted in this study. The carbon conversion coeffi cient ofSynwas 250 fg C/cell, Pro was 53 fg C/cell, and that of PEuks was 2 100 fg C/cell. We had the equivalence(Crosbie and Furnas, 2001):
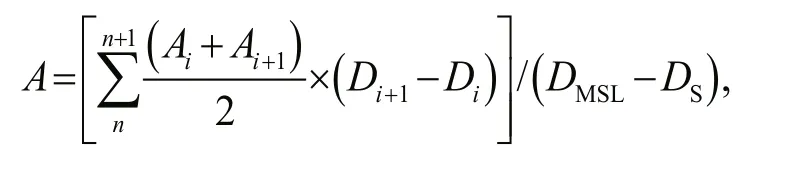
whereAis the carbon biomass of Pico,nis the number of sampling layer;Aiis the Pico biomass(μg C/L) at sampling layer;Diis the depth at sampling layeri(m);DMSLis the maximum sampling layer (m),DSis the surface sampling layer (0 m)(Wei et al., 2020).
3 RESULT
3.1 Temperature and salinity
Distribution of temperature and salinity in the surface water was shown in Fig.2. The surface temperature varied from 20.03 ℃ to 31.26 ℃ and the average temperature was 23.22 ℃. The highest temperature appeared at station S03-02, and the lowest was at station S01-06. The surface salinity varied from 21.29 to 34.68,and the average salinity was 33.31. The highest salinity appeared at station S04-01, and the lowest was at station S03-02. The overall trends were that temperature was higher in the south and lower in the north, and the salinity was higher in the east than in the west. Waters of relatively high temperature (>25 °C) and salinity(>32.5) occupied the northeast of Taiwan, China.
3.2 Nutrient
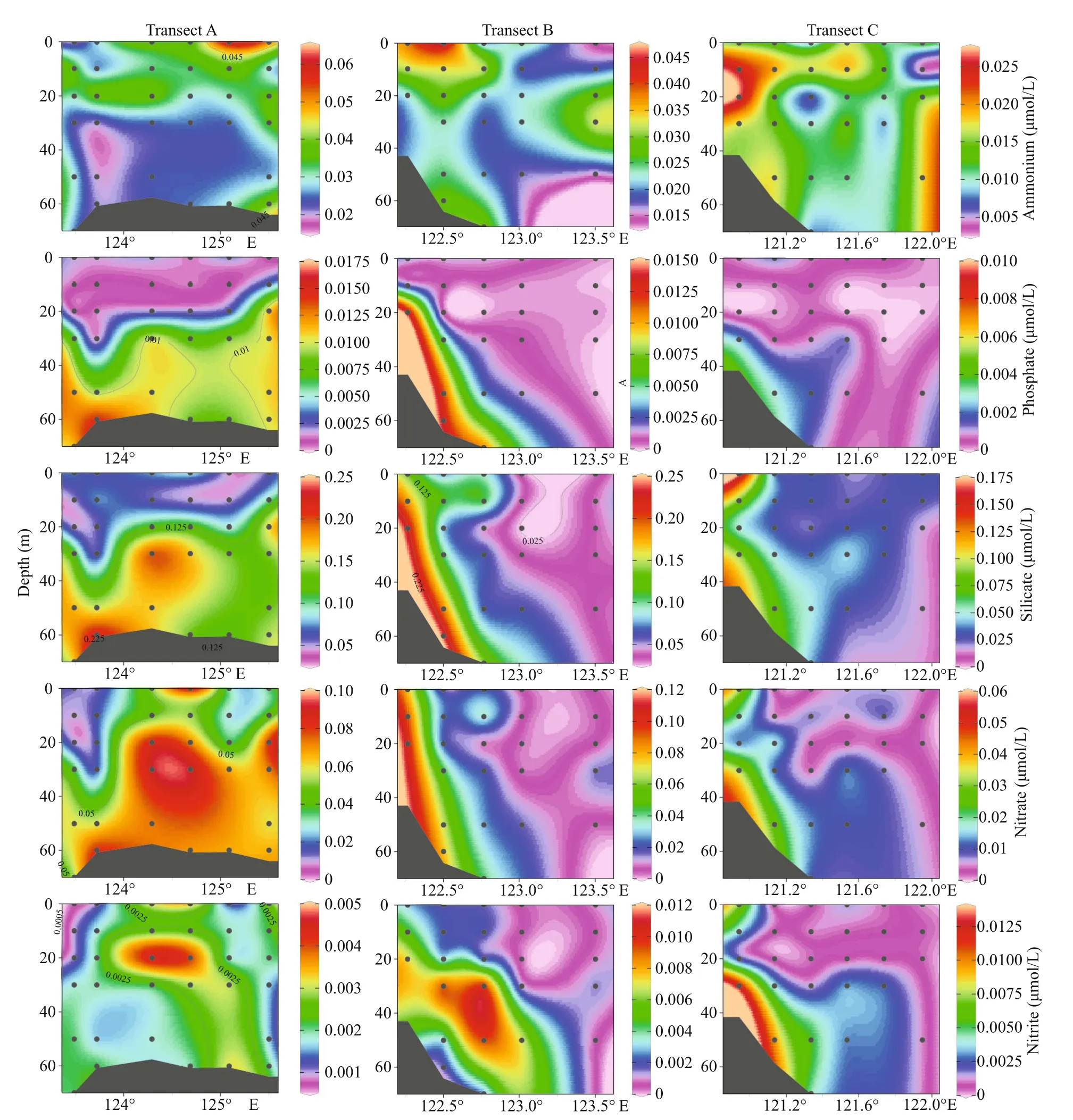
Fig.4 Section distribution of nutrients along transects A, B, and C
In the surveyed seawater (Fig.3), the surface concentration of ammonium ranged from 0.008 6 to 0.062 μmol/L, and the average concentration was 0.022 μmol/L. Station S04-03 had the highest concentration. The concentration of phosphate was between 0.000 2 and 0.002 4 μmol/L, and the average concentration was 0.001 2 μmol/L. Higher phosphate concentrations were mainly concentrated in section A.Station S01-07 had the highest concentration. The silicate concentration was between 0.021 and 0.286 μmol/L, with an average concentration of 0.096 μmol/L. Inshore silicate concentration was higher.Station L23-02 had the highest concentration. The nitrate concentration was between 0.003 and 0.139 μmol/L, and the average concentration was 0.035 μmol/L. Station L23-02 had the highest concentration. The concentration of nitrite was between 0.000 3 and 0.007 μmol/L, and the average concentration was 0.003 μmol/L. The distribution of nitrate and nitrite was approximately the same, with higher concentrations inshore. Station L23-02 had the highest concentration.Then we selected three sections from the study area to show the distribution of nutrients in more detail (Fig.4),we found that nutrients were higher in the bottom, and gradually decreased from bottom to the surface layer.
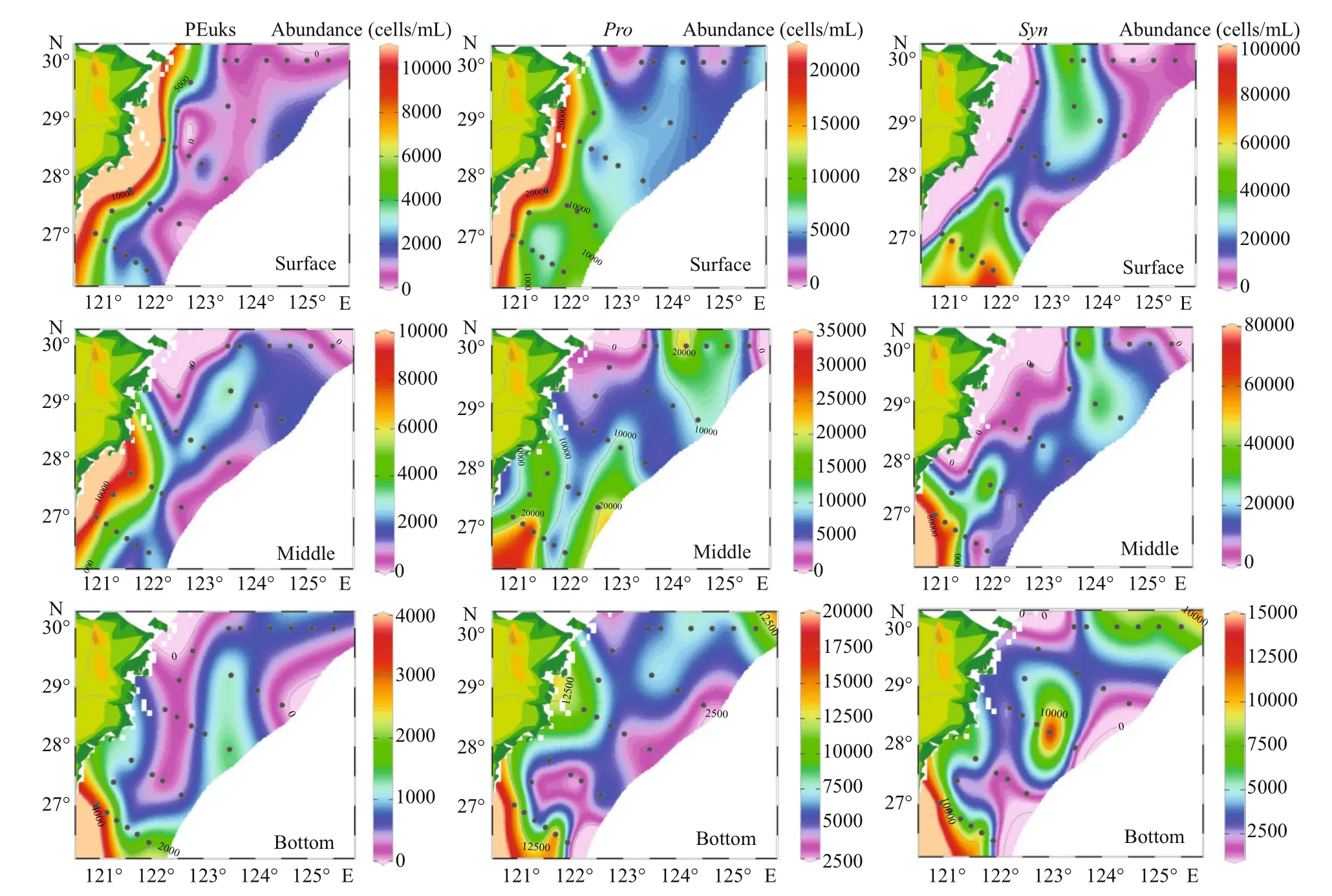
Fig.5 Horizontal distribution of diff erent Pico taxonomic abundance (cells/mL) in the surface, middle, and bottom layers
3.3 Distributions and compositions of Pico abundance
In our study, the cell abundance of diff erent Pico groups we obtained were roughly of the same order as other studies conducted in the ECS (Pan et al., 2005).The horizontal distributions of Pico are shown in Fig.5.Synwas the predominant group of Pico in the ECS, with a percentage of 65.32%. The PEuks andSynwere concentrated on the surface layer, and then the abundance was gradually decreased. However,Prowas concentrated in the subsurface layer. The abundance of PEuks was between 0.49×102-1.44×104cells/mL. In the investigated sea area, PEuks group had the highest abundance in the coastal region,and then decreased in off shore. The maximum PEuks concentration was recorded at station S03-01 (10 m),and the minimum was at station S02-01 (bottom layer,86 m). The cell abundance ofProranged from 1.36×103-3.47×104cells/mL. The maximumProabundance was recorded at station S05-03 (30 m),and the minimum was at station S01-03 (30 m).Syncell abundance ranged from 0.69×103-1.15×105cells/mL. The maximumSynconcentration was recorded at station S05-06 (20 m), and the minimum was at station S01-03 (30 m). BothProandSyngroups had the lowest abundance in the Transect A, and then the abundance increased inshore, with the highest value appearing in the northeast of Taiwan,China. The vertical distribution of Pico abundance presented an apparent stratification phenomenon(Fig.6). The distribution trend of Pico was first up and then down. The distribution of PEuks andProabundance increased gradually from 0 to 40 m depth and reached the maximum at 40 m. Thereafter, the abundance dropped to a minimum from 60 m to the bottom layer. Nevertheless, theSynabundance reached the maximum at 20 m, and then dropped to minimum from 40 m to the bottom layer.Synabundance was higher than PEuks andPro.
3.4 Pico carbon biomass
In our study (Fig.7),Synwas the main contributor to Pico carbon biomass, with a percentage of 52.72% in the ECS. Moreover, the proportion ofSynwas more than that of PEuks (42.41%) andPro(4.87%).
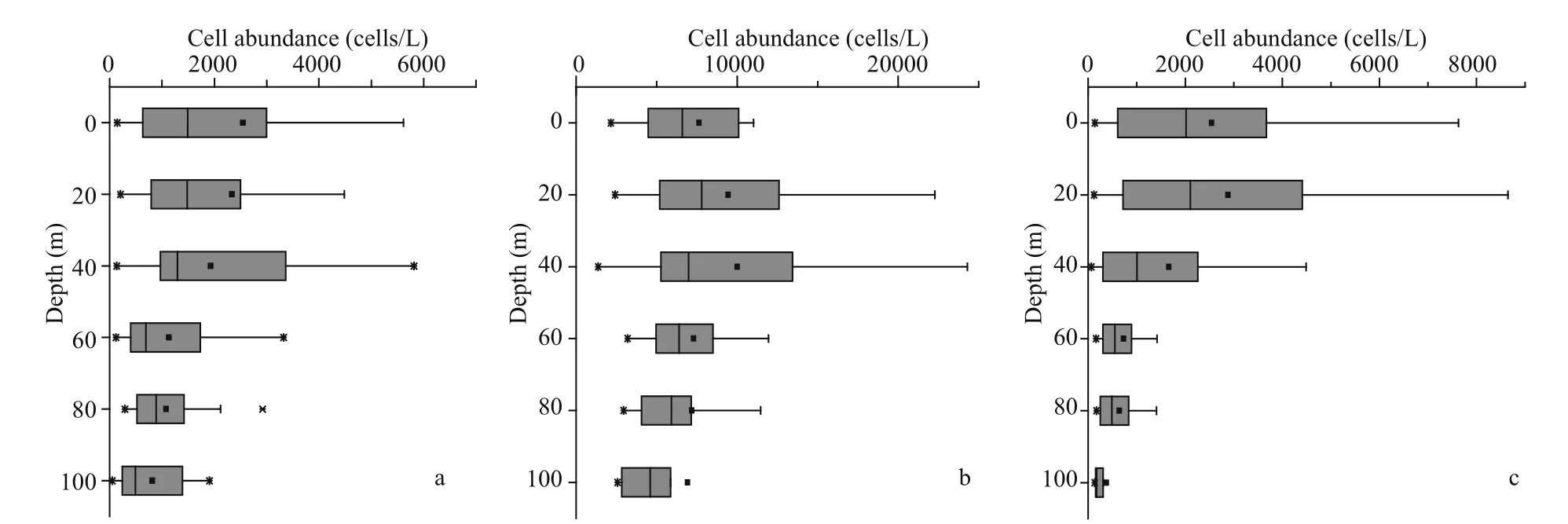
Fig.6 Vertical distribution of Pico abundance in the study area (cells/mL)
4 DISCUSSION
4.1 Relationship between the Pico community and environment
Using RStudio to analyze the correlation between Pico and environmental factors, the result was shown in the Fig.8. The environmental factors in this study included: temperature, salinity, depth, and nutrients(nitrate, nitrite, silicate, phosphate, and ammonium).Three Pico groups had diff erent distribution characteristics in the marine ecosystem (Fig.6). PEuks andSynmainly existed in the surface layer,Propreferred the subsurface layer. In general, temperature was necessary for the growth and reproduction of Pico. Indeed, a significant negative correlation existed between the three groups of Pico and depth. The light irradiation decreased with the increase of seawater depth, which leads to the decrease of Pico abundance.For taxa ofSyn, strains showed a significant positive correlation with temperature. A recently published study found that as the temperature rose to 27 °C, the growth rate of theSynincreased all the time without reaching a plateau (Mackey et al., 2013). Mackey et al. (2013) concluded thatSynwas able to regulate photochemistry in a certain temperature range by transforming state and changing the abundance of photosynthetic proteins.Synhas developed a mechanistic of temperature acclimation strategies to be in favor of the larger geographic range of this group. According to the above foundation, the relation betweenSynand temperature can be well explained.There was a significant positive correlation between PEuks and temperature, which indicated that temperature was an important control factor for PEuks distribution. Besides, PEuks group had a negative correlation with phosphate. Furthermore, temperature was a more important regulating factor than nutrients,salinity, and depth for PEuks. Meanwhile,Syngroup showed a positive correlation between temperature and salinity. In addition, depth and nutrients had a strong negative impact on them, which is in agreement with previous studies conducted in the ECS that Pico abundance would increase with the increasing of temperature and salinity (Chiang et al., 2002; Jiao et al., 2005). In common,Progroup had a positive correlation between temperature and salinity, but a negative correlation with depth and nutrients.According to the research report,Prolacked the functional genes related to the absorption and transformation of nutrients (Rocap et al., 2003).Indeed, high concentrations of nutrients in the coastal region had a toxic eff ect onPro(Paytan et al., 2009).These results explain thatProprone to oligotrophic seawater, and this was in accordance with previous studies in other tropical (Arabian Sea) and subtropical regions (North Pacific Ocean) (Liu et al., 1998;Shalapyonok et al., 2001). Compared withProand PEuks,Syngroup had higher growth potential than the other two Pico. This was an important advantage forSyngroup to be the dominator when conditions were favorable in the oligotrophic seawater. However,PEuks had high species diversity, which can keep stable in the mixing ocean (Guo et al., 2014).
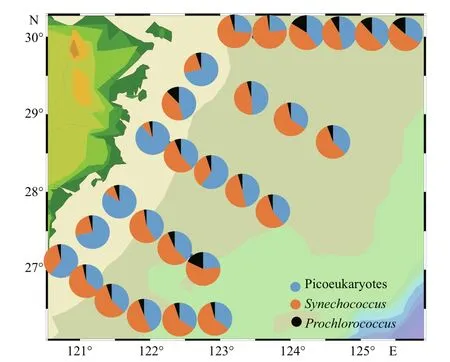
Fig.7 Spatial distributions of depth-weighted average carbon biomass of picoeukaryote, Synechococcus,and Prochlorococcus

Fig.8 Correlation analysis of three Pico groups with associated environmental variables
4.2 Coupling of hydrography and the Pico community
As we all know, the unique geographical location of the ECS made it more obviously aff ected by the currents. Four water systems (Fig.1b), the Taiwan warm current, Kuroshio current, Coastal current and Changjiang diluted water, played a crucial role in the growth and distribution of Pico in the ECS (Pan et al.,2005). The oligotrophic Kuroshio originated from the North Equatorial Current at about 10°N-15°N, and flowed northward along the east coast of the Philippines. The Kuroshio is famous for its strong velocity, and it has high temperature and salinity. The intrusion of the Kuroshio has a huge impact on the northeastern of Taiwan, China. It is the most considerable feature of the ECS current circulation(Le et al., 2012). The Taiwan warm current has high temperature and salinity throughout the year. The Coastal current is mainly distributed in the south of the Changjiang River estuary and along the Zhejiang and Fujian coasts. One of the main characteristics of this current is low salinity. The Changjiang diluted water is one of the largest rivers in the world. A large number of nutrients and other substances are continuously imported into the river through fresh water runoff , which greatly aff ects the ecosystem of Pico communities in the estuary (Jiao et al., 2002).Moreover, the Changjiang diluted water extends from the estuary to the northeast, and it has low salinity.
From the temperature and salinity distribution diagrams (Fig.2), both temperature and salinity were low in coastal areas and high in oceanic areas.Generally, the temperature and salinity parameters of the surveyed sea area were mainly aff ected by the current such as the Changjiang diluted water, Taiwan warm current, and Kuroshio (Chung et al., 2014). And the temperature was greatly aff ected by the Kuroshio and Taiwan warm current, the coastal and the Transect A had low temperature. On the other hand, the salinity was diluted by the Coastal current and Changjiang diluted water that contributed to low salinity inshore.The salinity gradually increased from the inshore to off shore. In the off shore area, the temperature and salinity were significantly higher, there was a thermohaline gradient between the coastal water and ocean. The above phenomenon indicated that the off shore area was less aff ected by the Changjiang diluted water, but more influenced by the Taiwan warm current and Kuroshio. In addition, the concentrations of nutrients increased with the increased of the total runoff when the Changjiang River entered the flood season in May (Li et al., 2007).There were some diff erences in nutrients concentration among diff erent water layers (Fig.4). Specifically, the concentrations of nutrients were highest in the bottom layer, followed by the middle layer and were lowest in the surface layer. The main reason for this phenomenon was that there were high concentrations of plankton in the surface and middle layers, and their growth played an important role in the consumption of nutrients. The higher nutrient concentration in the bottom seawater was due to the release of nutrients between the sediment interfaces on the sea floor, as well as the invasion of high-salinity and high-nutrient seawater, such as Taiwan warm current, upwelling,etc.
Furthermore, the mixture of currents was one of the important factors aff ecting the distribution of Pico(Xiu and Huang, 2006; Le et al., 2012). In spring,Kuroshio water invaded the northeast region of Taiwan, China and intruded into the continental shelfin the northeast of Taiwan, China. In our study, the surface seawater was typical features oflow temperature and high salinity in the upwelling region.This can explain the phenomenon that high Pico abundance encountered in the coastal area of Fujian,the shelf sea, and the upwelling area in the northeast of Taiwan, China. As temperature increased, there was an increased in the Pico abundance, indicating that temperature was a more important regulating factor for Pico, especially when the temperature exceeded 26 ℃. The spatial diff erentiation of PEuks was mainly encountered in the continental shelf area.Related studies have shown that while cyanobacteria is very little diversified, at least at the genus level with a couple of taxa (Prochlorococcus,Synechococcus)dominating, eukaryotes turned out to be very diverse with picoplankton taxa distributed widely across several branches of eukaryotic tree oflife (Vaulot et al., 2008). In addition to PEuks had more complex genome thanSynandPro, it also had a greater demand for nutrients absorption and utilization. Pan et al.(2007) proved that PEuks are the most competitive among Pico in near-shore waters in the Changjiang River estuary with abundant river-borne nutrients.The coastal areas of Zhejiang and Fujian and the Changjiang River estuary were close to human activity areas and nutrients rich.Procontained a unique photosynthetic pigment—divinyl chlorophyll,which was light dependent. It was mainly distributed in oligotrophic waters in tropical and subtropical oceans. Moreover, a large number ofliterature data showed thatProtended to be more abundant in oligotrophic rather than eutrophic waters (Jiao et al.,2002; Pan et al., 2007; Wei et al., 2019a, b). From the distribution ofProin the surface, subsurface, and bottom layer, we would infer that the warm and oligotrophic Taiwan warm current had a strong influence on thePro. This result was consistent with previous study results (Partensky et al., 1999; Jiao et al., 2002; Le et al., 2012). In spring, there was lessProin the coastal area. The abundance ofSynwas low at latitudes above 28°N, and this phenomenon was most likely a combined consequence of the low temperature and light restrictions. In summation, Pico with high abundance and biomass was found in the upwelling area located in the northeast of Taiwan,China. On this foundation, Pico grew and reproduced rapidly under suitable physicochemical conditions(such as temperature, salinity, and nutrients), and provided abundant primary productivity for marine ecosystem, inferred from a preliminary analysis.However, the influencing factors and control mechanism of Pico still need further research.
5 CONCLUSION
Syn, PEuks, andProhave a wide geographical distribution that cover both neritic and oceanic in the ECS. In spring, according to the abundance and distribution of Pico in the marine ecosystem,Synwas predominant in Pico groups, followed byProand PEuks. However, in terms of carbon biomass,Synand PEuks were the major contributor,Proonly made up a very small part of carbon biomass. There was no uniformity in both cell size and carbon conversion factors, and this has contributed to large uncertainties in carbon biomass of Pico. In our study, the calculated carbon biomass was not compared with other seasons,or with other seas, mainly because the abundance-tocarbon conversion factors were not uniform across other studies, and the results of such comparisons were not scientifically meaningful. Temperature,salinity, and nutrients were important factors aff ecting the distribution of Pico in ocean. Secondly, the input of the four water systems in the ECS, and the mixing of water systems had a significant impact on the vertical stability of water, which aff ected the distribution of Pico and resulted in the diff erences from other oceans. However, the interaction between Pico communities and its influence mechanism needs to be further explored. Other approaches such as molecular biology combined with cell sorting are needed to identify organisms at the species level.
6 DATA AVAILABILITY STATEMENT
The data used to support the findings of this study are available from the corresponding author upon request.
7 ACKNOWLEDGMENT
We thank the Open Cruise Project in the East China Sea of National Nature Science Foundation of China(NORC2017-02) for sharing the ship time.
 Journal of Oceanology and Limnology2021年4期
Journal of Oceanology and Limnology2021年4期
- Journal of Oceanology and Limnology的其它文章
- Numerical study of the seasonal salinity budget of the upper ocean in the Bay of Bengal in 2014*
- Study on evaluation standard of uncertainty of design wave height calculation model*
- A fast, edge-preserving, distance-regularized model with bilateral filtering for oil spill segmentation of SAR images*
- A Gaussian process regression-based sea surface temperature interpolation algorithm*
- Climatology and seasonal variability of satellite-derived chlorophyll a around the Shandong Peninsula*
- Sources of sediment in tidal flats off Zhejiang coast, southeast China*