
高寒植物功能性状对种子萌发率与出苗率的影响
2021-07-26 08:56:22武学霞陈冠光李兰平
青海大学学报 2021年4期
武学霞,陈冠光,李兰平*
(1.省部共建三江源生态与高原农牧业国家重点实验室,青海大学,青海 西宁 810016; 2.兰州大学生命科学学院,甘肃 兰州 730000; 3.兰州大学资源环境学院,甘肃 兰州 730000)
种子是被子植物有性繁殖的载体,对于物种的延续与种群的扩大具有至关重要的作用,因此引起了众多学者的关注[1-4]。目前国内对于种子的研究主要集中在种子的大小及其变异[4-8]、不同物种种子萌发特性[9-14]、温度对植物种子萌发的影响[15-16]、光照对不同物种种子萌发与幼苗建植的影响等[17-18]。
功能性状是指在形态上、生理上或物候上能够反映植物的生态学策略与决定植物如何响应环境因素的一些特征[19]。如高度、比叶面积分别与植物的光截获能力和光合作用能力息息相关[20]。目前已有学者将植物功能性状用于预测特定区域物种的多度[20]、热带雨林移植幼苗的存活率[21]及对高寒植物与旱区植物萌发率、幼苗存活率的影响[17,22],尽管这些研究结果有所差异,但也说明通过功能性状来预测植物种子的萌发率与出苗率是具备一定可行性的。以上相关研究功能性状数据来源于植株幼苗,本研究采用成熟植株的功能性状而并非是幼苗的功能性状,这对于通过植物性状来预测其种子萌发率及在建植过程中的表现有一定意义,同时也可为人工草地与半人工草地建植草种的选择提供一定的指导。通过对甘肃省玛曲县兰州大学高寒草甸与湿地生态系统定位研究站周边常见的30种典型高寒草甸植物功能性状及其萌发率与盆栽出苗率进行研究,以期阐明植物性状与其种子萌发率及出苗率之间的关系。
1 材料与方法
1.1 研究区域概况
试验地点位于甘肃省玛曲县兰州大学高寒草甸与湿地生态系统定位研究站。地理位置为33°40′N,101°52′E,海拔3 550 m;年平均气温1.2 ℃,月平均气温为-10 ℃(1月)~11.7 ℃(7月);年均降水量约为600 mm,主要集中在6—8月;年平均日照2 583.9 h,年冷季长达314 d,暖季约51 d[23]。研究区草地类型属于高寒草甸,土壤类型主要为亚高山草甸土。
1.2 功能性状测定
测定的性状特征主要包括植物种子百粒重、种子数、高度、个体生物量、比叶面积与繁殖分配。供试种子收集于2013年,研究站高寒草甸试验样地内。采集的种子在室内自然风干、去除杂质后保存在干燥通风处。随机选取100粒所采物种种子进行称量,重复6次,精确到0.000 1 g,其平均值为该物种种子百粒重。种子数为各物种植株个体所产生的种子数量,具体测定方法为采集各物种种子开始掉落前植物个体10~30株,统计个体种子数量,取平均值为该物种种子数。当所选物种处于开花以及之后物候期的植株比率超过50%时,在试验样地随机选取12~30株处于花期的个体,且所取样本仅限于植物的地上部分。对于克隆植物,将一个无性系分株作为一个个体[24]。每株植物在采集后立即测其高度,并随机取下2片(部分物种1片或多片叶片)成熟健康的叶片测量其叶面积,之后将其在80 ℃下烘干48 h后进行称重,用于计算比叶面积。植株剩余部分(除测量叶面积叶片外)同样在80 ℃下48 h烘干后称重,其中植株繁殖部分单独称量,最后计算生物量与繁殖分配。
1.3 萌发试验
于2014年4月底,随机选取健康、成熟的种子,在直径9 cm的玻璃培养皿中进行萌发试验,培养基为蒸馏水润湿的2层滤纸。萌发试验在兰州大学高寒草甸与湿地生态系统定位研究站内人工气候箱中进行,人工气候箱设置参数为20 ℃/5 ℃、12 h光照/12 h黑暗(近似于研究区域植物萌发季节的平均最高和最低气温)。每个培养皿均匀放置50粒种子,每个物种设置3次重复。以胚根突破种皮作为萌发的标准,每天记录萌发种子数,并将已萌发的种子移出培养皿,萌发周期持续50 d。
1.4 盆栽试验
盆栽试验开展时间为2014年4月底至2014年9月中旬。基质为试验站内草甸土,经过筛除去杂质后置于直径为17 cm花盆内,基质用量约为花盆容量的3/4。而后将50粒饱满的供试种子均匀撒在基质上,并覆盖细沙一层,每个物种重复3次。定期浇水,防止基质干燥。每月记录幼苗出土情况。
1.5 数据统计和分析
对原始数据进行Spearman相关性分析,之后对种子萌发率、出苗率进行反正弦转换,对百粒重、种子数、比叶面积、生物量及植株高度进行对数转换,并删除萌发率与出苗率同时为0的5种物种,用Pearson相关性分析与线性回归分析种子萌发率、盆栽出苗率与百粒重、种子数、比叶面积、生物量、繁殖分配及植株高度之间的关系。采用SPSS 17.0与R 3.3.5进行所有的数据转换与分析。
2 结果与分析
本研究中,供试种子平均萌发率为36%,平均出苗率为5%,平均百粒重为0.113 g(表1)。成熟植株种子数、繁殖分配、比叶面积、高度与生物量分别为232.39粒、20%、201.99 cm2/g、28.65 cm与0.633 g(表 1)。

表1 所选物种种子萌发率、出苗率及其功能性状
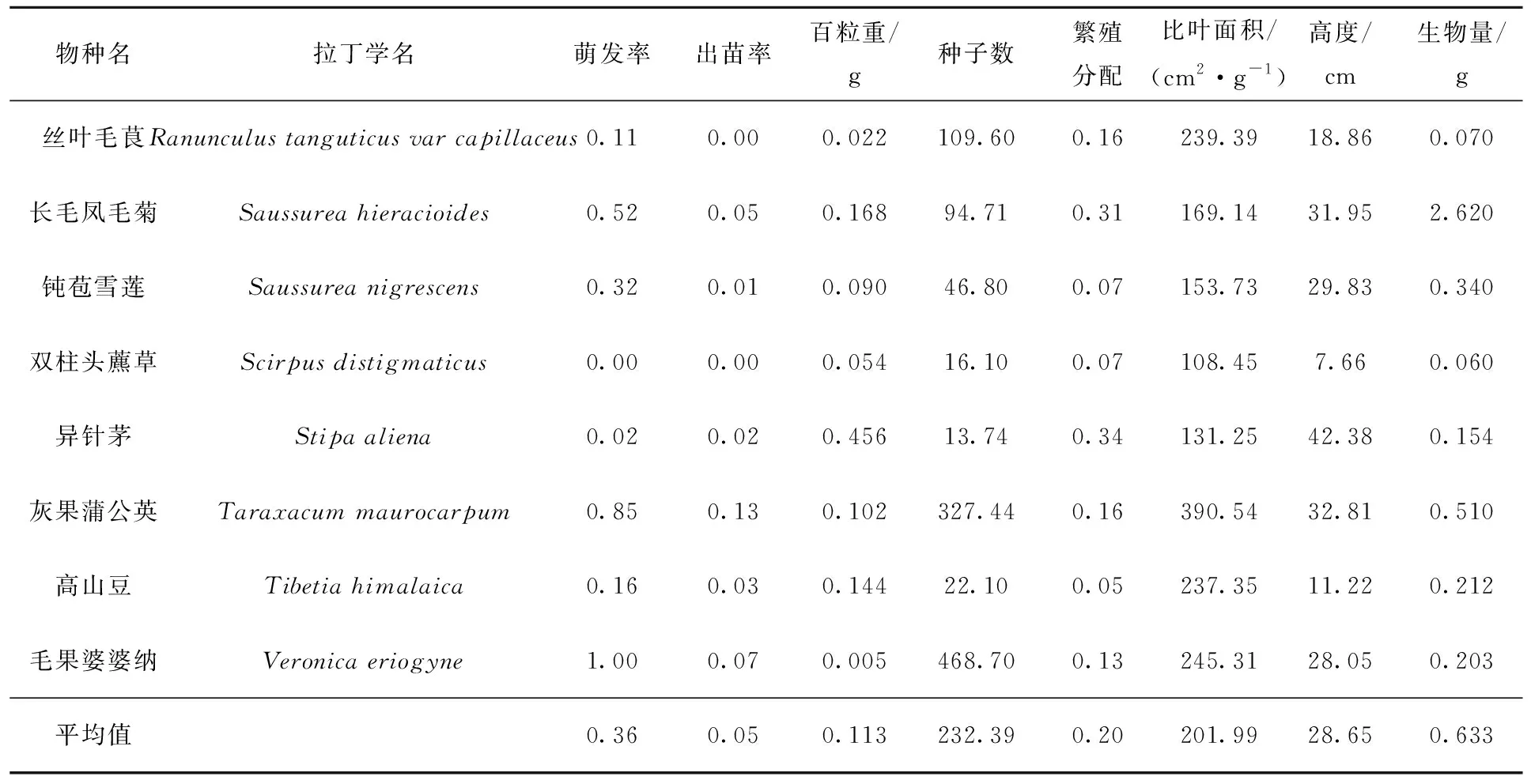
表1(续)
Spearman相关性分析表明,萌发率与种子数呈极显著正相关(rho=0.463,P<0.01),与其余性状无显著相关性(表 2);出苗率与萌发率(rho=0.704,P<0.01)、种子数(rho=0.500,P<0.01)、生物量(rho=0.597,P<0.01)及高度(rho=0.468,P<0.01)均表现出极显著正相关(表 2)。

表2 萌发率、出苗率与功能性状Spearman相关性分析结果
经过数据转换与物种筛选后,Pearson相关性分析结果与Spearman相关性分析结果一致:萌发率与种子数呈极显著正相关(r=0.613,P<0.01);出苗率与种子数(r=0.486,P<0.05)呈显著正相关,出苗率与萌发率(r=0.549,P<0.01)、生物量(r=0.625,P<0.01)及高度(r=0.588,P<0.01)均呈极显著正相关(表 3)。

表3 萌发率、出苗率与功能性状Pearson相关性分析结果
回归分析中,仅种子数能较好地线性拟合萌发率(R2=0.38,P<0.01,图 1),说明在玛曲县常见高寒植物中,种子量较大的植物往往具有较高的种子萌发率。而种子数(R2=0.24,P<0.05)与出苗率有显著线性关系,生物量(R2=0.39,P<0.01)、高度(R2=0.35,P<0.01)、萌发率(R2=0.30,P<0.01)与出苗率有极显著线性关系(图 2和图 3),说明植物部分功能性状可较好地预测出苗率。
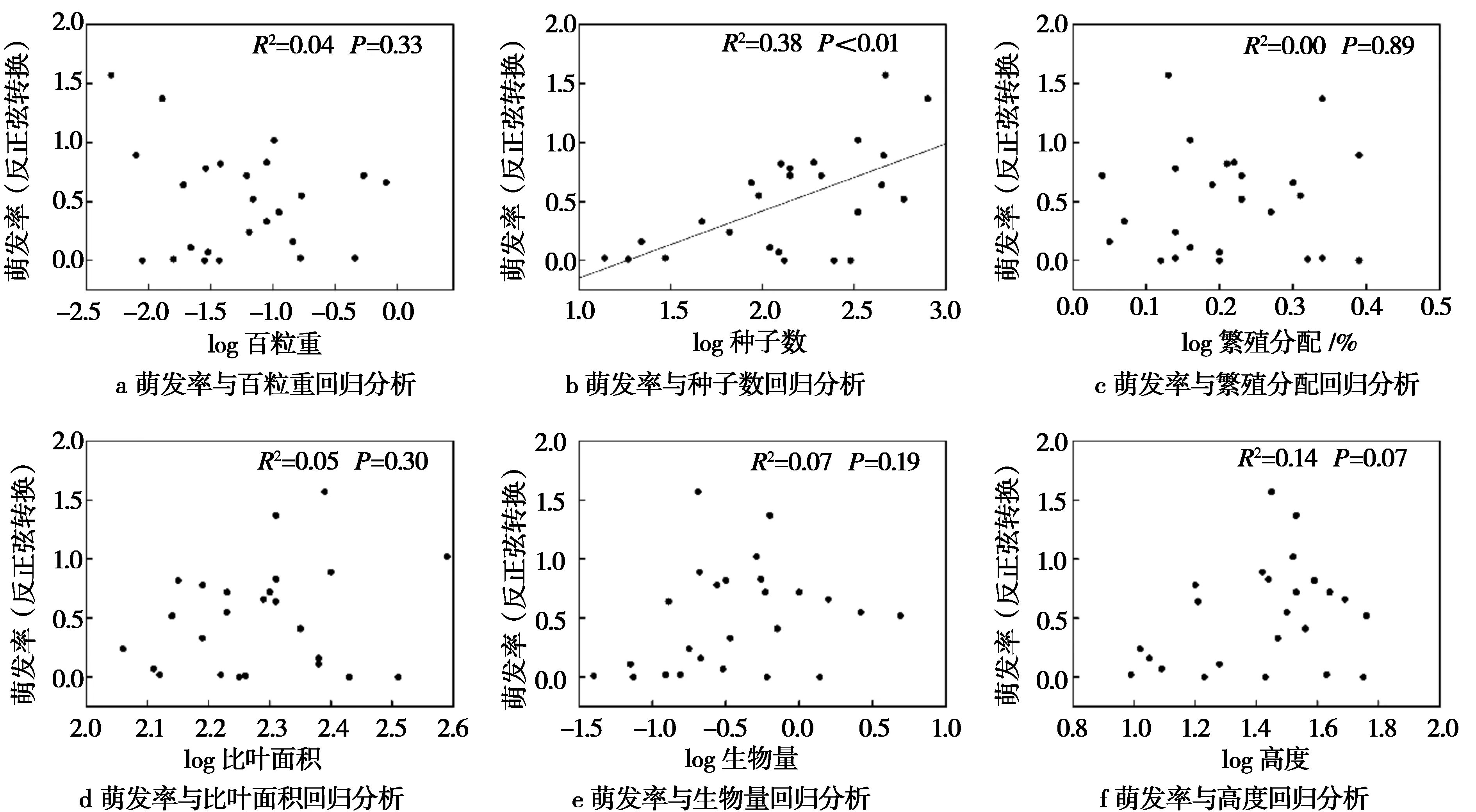
图1 萌发率与功能性状回归分析结果Fig.1 Regression analysis of germination rate and functional traits
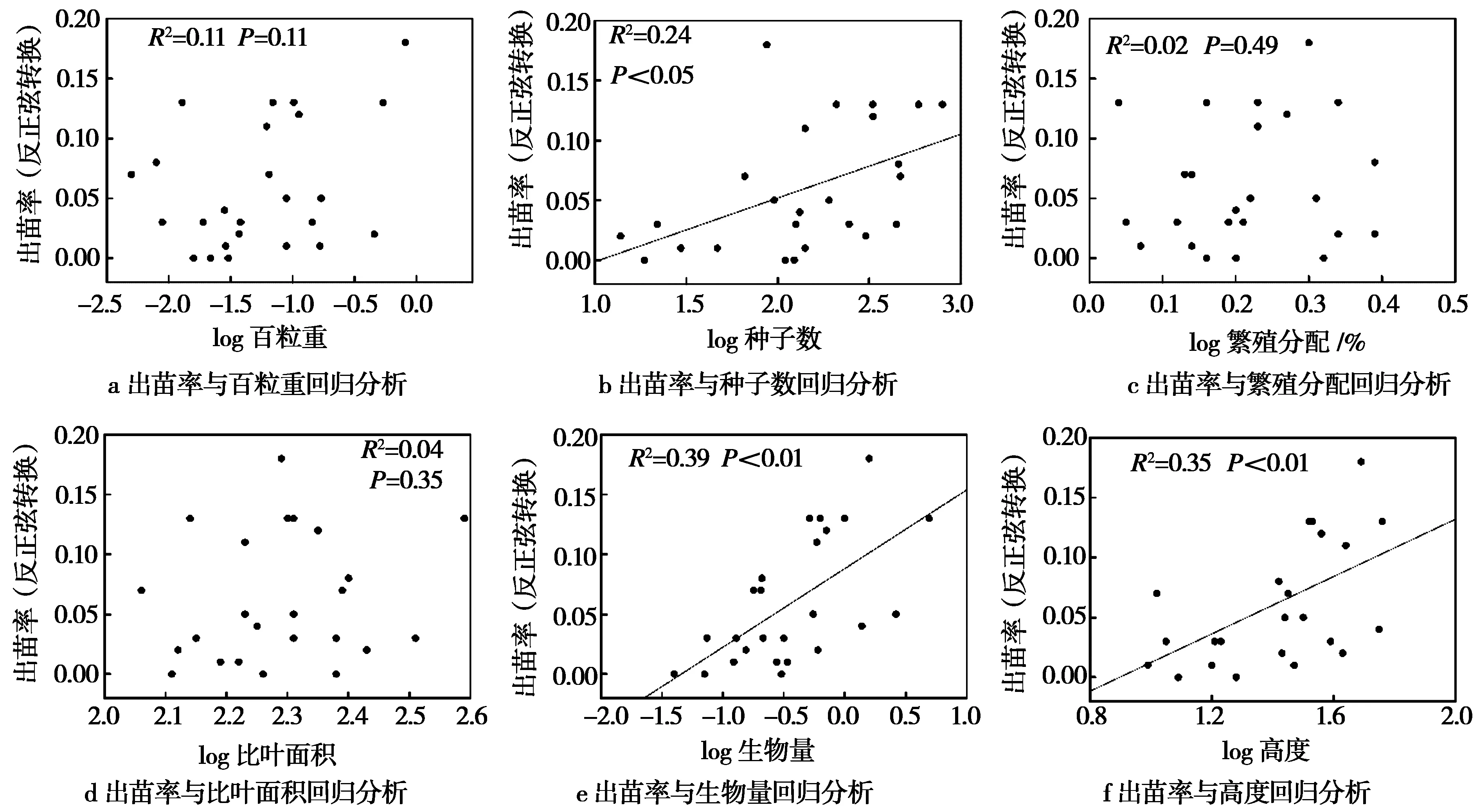
图2 出苗率与功能性状回归分析结果Fig.2 Regression analysis of emergence rate and functional traits
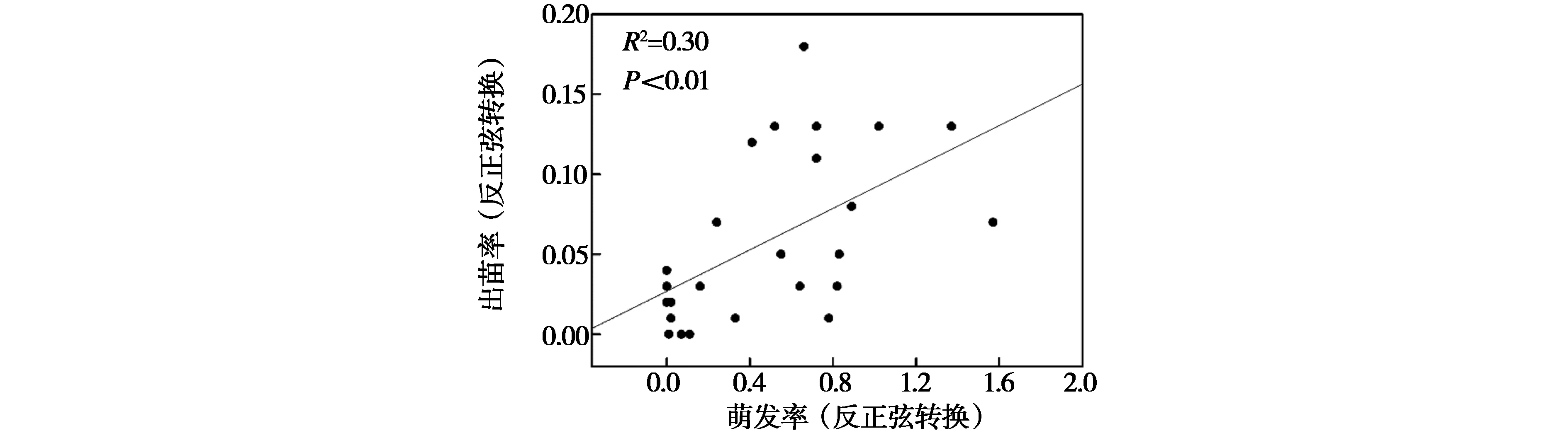
图3 出苗率与萌发率回归分析Fig.3 Regression analysis of emergence rate and germination rate
3 讨论与结论
种子的大小在植物功能性状对种子萌发和幼苗建植影响的研究中较为多见,一般认为同一物种种子萌发率、出苗率与其大小呈正相关[3]。但是在不同物种间,有的研究认为种子大小与萌发率呈显著的负相关[25-26],也有研究认为种子大小与萌发率和出苗率呈正相关[27],或与萌发率并没有显著相关性[10,28]。本试验研究结果支持玛曲县常见高寒植物种子大小与萌发率和出苗率均无显著相关性,种子大小对萌发率和出苗率的影响可能在逆境中会更容易观察到[6]。
种子数与萌发率和出苗率呈极显著正相关,这可能是因为产生种子数量较多的物种大多为冒险型物种;即在种子开始萌发时,大部分种子会在较短时间内萌发[29],因此具有相对较高的萌发率与出苗率。与Liu等[30]发现不一致的是在本研究中植物高度和生物量与出苗率呈极显著正相关,但与萌发率没有显著相关性。可能是因为高度较高和生物量较大的高寒植物在群落中往往具有一定的竞争优势,相对较高的出苗率是该优势在其幼苗建植过程中的表现,但该优势无法在种子萌发过程中体现。同时,本研究发现萌发率可较好地预测种子出苗率,因此在使用未知野生物种种子进行人工草地或半人工草地建植之前,可通过萌发试验初步预测其出苗情况。
总之,本研究表明在本试验研究区域用植物功能性状预测其种子的萌发率与出苗率是可行的,可用于预测出苗率的功能性状包括种子数、生物量与高度,而可用于预测萌发率的功能性状仅有种子数。因此在选择用于人工草地与半人工草地建植的物种时,可以优先考虑产生种子数量较多、生物量较大或高度较高的物种,以获得相对较高的幼苗出苗率。
猜你喜欢
中国农业科学(2022年13期)2022-07-26 09:55:28
辽宁农业科学(2021年5期)2021-11-03 07:19:30
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02 01:48:06
新疆农业科学(2020年1期)2020-02-14 03:22:46
新农业(2018年6期)2018-04-18 07:30:36
青苹果·教育研究版(2016年7期)2016-12-12 05:19:52
考试周刊(2016年11期)2016-03-17 05:11:15
现代农业(2016年5期)2016-02-28 18:42:34
中国麻业科学(2015年5期)2015-12-28 06:22:11
安徽农业科学(2015年25期)2015-12-22 06:21:16
