
石湖园林植物LES性状对水分环境响应的研究
2021-07-25 10:38曲彦婷张鹏远张姗姗李英姿李雨桐
东北农业大学学报 2021年6期
陈 珊,张 兴*,曲彦婷,张鹏远,张姗姗,李英姿,李雨桐
(1.苏州科技大学建筑与城市规划学院,江苏 苏州 215129;2.黑龙江省科学院自然与生态研究所,哈尔滨 150040;3.西交利物浦大学理学院,江苏 苏州 215127)
近年,随城市化进程不断加快,加剧甚至改变气候系统变化进程,造成以温度升高、极端事件频发等为主要特征环境变化,严重影响生态系统构成和功能,给城市生态健康带来严峻挑战[1-2]。定量检测城市生态系统健康状况并及时采取应对措施是实现城市可持续发展有效途径。园林植物作为有效环境监测生物学工具[3],在固碳释氧、净化空气、调节小气候等方面均具有重要作用。植物叶片对环境因素变化非常敏感,环境因子影响叶片性状,改变不同植物生长状况,进而导致植物种间关系,群落中物种组成或生态系统能量和物质循环流动[4-5]。
水分是限制植物地理位置分布最重要因素之一,也是决定群落循环的重要环境因子。本文探索小尺度下植物叶经济谱性状对不同水分环境因子响应机制,有针对性提出面向区域环境下园林植物选择框架方法,以期为日益遭到自然及人为破坏的区域生态环境保护与修复提供科学依据。
1 材料与方法
1.1 研究区域
石湖景区(120°36′6.87028″E,31°15′49.64921″N)位于江苏省苏州市吴中区,总面积34.62 km2,是苏州著名山水型自然风景名胜区,其人为扰动较严重,植被种类丰富且分布较为密集。根据土壤烘干法并兼顾取样点土壤成分及大气条件相对一致的设点原则,选择3个具有代表性的不同水分条件样点,即石湖滨湖区(高水分条件区)、植物园区(中水分条件区)与上方山区(低水分条件区)。
1.2 试验材料
通过实地踏查及分析,在3个样地分别采集生长状况相对一致且具有良好长势9种典型园林植物当年生新鲜叶片为研究材料,分别为桂花[Osmanthus fragrans(Thunb.)Lour.]、香 樟(Cinnamomum septentrionaleHand.-Mazz.)、银 杏(Ginkgo bilobaL.)、红 枫[Acer palmatum'Atropurpureum'(Van Houtte) Schwerim]、榔榆(Ulmus parvifoliaJacq.)、紫薇(Lagerstroemia indicaL.)、石楠[Photinia serratifolia(Desf.)Kalkman]、南天竹(Nandina domesticaThunb.)、毛鹃(Rhododendron×pulchrumSweet)。选取每个树种长势良好当年生枝条10枝,佩戴聚乙烯塑料手套,在每个向阳面光照强度一致枝条上选择20片完全展开并无病虫害成熟新鲜叶片作为样本,采样后立即放入自封袋内并编号,然后放入干冰泡沫箱内带回实验室,从中随机抽样30片叶片于冰箱保鲜层中储藏待用。
1.3 测定项目与方法
1.3.1 叶经济谱性状测定
从采集带回的叶片中随机抽取20份样本,使用精确值0.0001电子天平称取叶饱和鲜质量(LFW),游标卡尺(精确至0.01 mm)测量叶厚度(LT),将样本叶片置于温度控制在55℃烘干箱内48 h至恒重,称取叶干质量(LDW),计算干物质含量(LDMC)。

使用叶面积扫描仪MICROTEK Scan Maker i800plus测定20份样本叶片叶面积(LA)、叶片长度(LL)、叶片宽度(LW)、叶周长(LP)等指标,并计算比叶面积(SLA)和比叶重(LMA)。
此外,作品以含蓄的文体风格凸显出“冰山”的八分之七。作品中无论是景色的描写、人物动作以及人物语言都是简而概之。如“当一大股暗黑色的血沉在一英里深的海里然后又散开的时候,它就从下面水深的地方蹿上来。它游得那么快,什么也不放在眼里,一冲出蓝色的水面就涌现在太阳光下。”这段描写并没有通过比喻或形容词来渲染,但鲨鱼的迅猛、凶狠以及当时紧迫的局势却生动直观地呈现在读者脑海中,使人有身临其境之感。

将烘干叶片研磨至粉末状,凯氏定氮仪PEIOU SKD-800测定叶氮,计算单位质量叶氮含量。

式中,C为盐酸溶液浓度(mol·L-1);V为滴定样品使用盐酸标准溶液消耗体积(mL);m为烘干样品粉末质量(g)。
1.3.2 水分生理指标测定
从采集回来叶片中各随机选取10片,0.0001天平称重得鲜质量W1,然后将新鲜叶片放入蒸馏水中饱和吸水24 h后取出叶片并将叶片表面水分吸干,称重得饱和鲜质量W2,最后把叶片放入80℃烘箱内烘干至恒重获得叶片干质量W3,计算叶片相对含水量(RWC)和相对水分亏缺(RWD)。
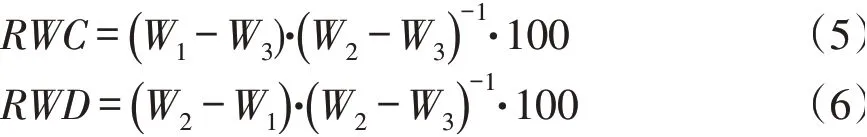
同时,在样本叶片中随机选取10片,0.0001天平称重得鲜质量W1,然后将叶片置于实验室内让其自然失水12 h后称重得W2,计算叶片离体失水速率(RWL)。

1.3.3 数据处理与分析
通过SPSS 22.0软件对叶经济谱性状作单因素(one-way ANOVA)方差分析和Duncan法多重比较(α=0.05)分析,并用Pearson法分析叶经济谱性状在不同物种及不同水分区间相互关系。最后,运用主成分分析法综合分析石湖风景区不同园林植物各叶片性状,筛选主要性状指标,通过模糊隶属函数分析园林植物在不同水分生境环境适应性。
平均隶属函数值计算公式如下,植物叶片性状与环境适应性呈正相关关系,则用公式(8)计算。
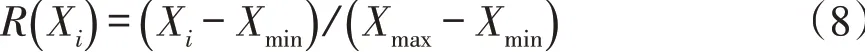
植物叶片性状与环境适应性呈负相关关系,则用公式(9)计算。
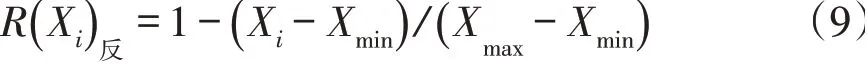
式中,Xi为第i个指标测定值,Xmax为所测指标最大值,Xmin为所测指标最小值。
2 结果与分析
2.1 叶经济谱性状之间差异性分析
从物种水平看,石湖风景区9种典型园林植物LP、LA等与植物叶片外在形态指标相关叶经济谱性状均存在极显著差异(P<0.01)。叶片大小直接影响植物光合能力和获取碳资源能力[4],群落平均叶片大小与环境中降水量直接相关,与温度相关性次之。由表1可知,9种园林植物中,银杏LT最大,红枫LT最小;香樟LA最大,南天竹LA、LP最小。植物常通过增加叶厚度以抵抗物理胁迫,且当水分供给不足时,通过减小叶面积减少水分消耗及降低高蒸腾带的迫害,因此,群落中南天竹较其余植物更适合在水分较少生境中种植,银杏次之。SLA、LDMC、LMA、Nmass是反映植物对资源开展投资、利用、回报的关键叶经济谱性状。石湖风景区中除LDMC外,其他9种园林植物SLA、LMA与Nmass同样存在极显著差异(P<0.01)。不同园林植物SLA排序为红枫>香樟>紫薇>南天竹>榔榆>银杏>毛鹃>石楠>桂花,除桂花、石楠、毛鹃、银杏之间,毛鹃、榔榆、南天竹、紫薇之间,银杏、榔榆、南天竹、紫薇、香樟之间,石楠、榔榆之间和紫薇、红枫之间无显著差异外,其他植物之间均差异显著。石湖风景区9种园林植物LMA排序为桂花>石楠>毛鹃>紫薇>南天竹>银杏>榔榆>香樟>红枫,除红枫、香樟、榔榆、银杏、南天竹之间,香樟、紫薇之间及榔榆、紫薇、毛鹃、石楠之间无显著差异外,其他植物间均差异显著。Nmass排序为南天竹>香樟>毛鹃>桂花>红枫>石楠>紫薇>银杏>榔榆,除榔榆、银杏之间,银杏、紫薇之间,紫薇、石楠之间及红枫、桂花、毛鹃之间无显著差异外,其他植物间均差异显著。因此,在石湖风景区中,香樟、红枫和南天竹位于叶经济谱中具有比叶重小、叶氮含量高、光合能力强、叶寿命短等特点,即快速投资-收益型物种一端,石楠、银杏和桂花在经济谱中偏向处于具有比叶重大、叶氮含量低、光合能力弱、叶寿命长等特点,即缓慢投资-收益型物种一端,而其余植物性状在叶经济谱位置中变化表征不明显,说明榔榆、毛鹃和紫薇对石湖风景区外界环境胁迫响应不明显。

1状性谱济经叶物植林园同不区景风湖石 表aerAcinecSuhihSnistnalpnedragtnereffidfomurtcepscimonocefaelfoscitsiretcarahC1elbaT N AMLALSALCMDLTLPL ssam )%()mc·g()g·mc()mc()g·g()mm()mc(2-1-2 21-花桂c3953.0±5374.1a3800.0±3230.0c2190.01±6382.33a3504.31±3512.51a9640.0±9355.0ba6130.0±0092.0b0583.61±0978.74.ruoL).bnuhT(snargarfsuhtnamsO樟香b3232.0±3087.1c7200.0±2900.0ba1840.43±3319.711a6994.6±1159.71ba1701.0±7774.0c0310.0±2271.0c5746.02±8000.33 .zzaM-.dnaHelanoirtnetpesmumomanniC杏银e2650.0±4988.0cb7300.0±1410.0cb1255.22±3679.57b2145.6±1685.11b7160.0±2633.0a2920.0±3323.0c8391.31±6319.13.LabolibogkniG枫红c6051.0±3183.1c0200.0±1700.0a9708.26±8796.651b6668.4±8576.11ba9011.0±9034.0d0510.0±3390.0a5174.91±0582.55mirewhcS)ettuoHnaV('muerupruportA'mutamlaprecA榆榔e0180.0±6107.0cb9400.0±5310.0cb3315.92±2492.38c4862.3±8460.6ba0230.0±8324.0cb3560.0±9812.0d9695.81±7427.42 .qcaJailofivrapsumlU薇紫ed4280.0±199.0cb3310.0±6510.0ba0646.66±5619.011a0715.61±5681.71a7305.0±5355.0b5470.0±0062.0dc9158.51±9575.72 .LacidniaimeortsregaL楠石d6751.0±8921.1b3900.0±3120.0c7469.22±8874.55a5642.6±3356.61ba4380.0±6854.0ba8950.0±0082.0dc0129.7±3541.03namklaK).fseD(ailofitarresainitohP竹天南a2333.0±2282.2cb9800.0±9410.0b4050.58±8741.701c1629.1±3180.3ba7950.0±1084.0c0420.0±6551.0e4293.6±7076.11 .bnuhTacitsemodanidnaN鹃毛c1212.0±0935.1b0800.0±8810.0cb0349.62±8464.26c9057.1±8024.4ba5670.0±5434.0ba5790.0±1182.0e3238.3±7370.31 teewSmurhclup×nordnedodohR=N=AML=ALS=AL=CMDL=TL=PL50.0<P量含氮叶量质位单 ;重叶比 ;积面叶比 ;积面叶 ;量含质物干 ;度厚叶 ;长周叶 。) (著显异差示表母字同不列同:注ssam cificepS=AML;aerafaelcificepS=ALS;aerafaeL=AL;tnetnocrettamyrD=CMDL;ssenkcihtfaeL=TL;ecnerefmucricfaeL=PL.)50.0<P(ecnereffidtnacifingisetacidninmulocemasehtnisretteltnereffiD:etoN ssamtinureptnetnocnegortinfaeL=N;thgiewfael ssam
由图1可知,同种植物在不同水分生境之间变化较大。研究发现,石湖风景区9种园林植物叶经济谱性状仅有LP、Nmass在3个不同水分生境中均存在显著差异(P<0.05),表明LP和Nmass是反映该区域植物对不同水分生境响应良好指标。桂花、银杏、红枫和榔榆LP在3个不同水分生境中均表现为植物园区>上方山区>滨湖区,紫薇和南天竹表现为滨湖区>植物园区>上方山区,石楠和毛鹃表现为上方山区>植物园区>滨湖区,仅有香樟LP大小表现为上方山区>滨湖区>植物园区。银杏、榔榆、紫薇、石楠和毛鹃Nmass在3个水分梯度环境中均呈滨湖区>上方山区>植物园区趋势,银杏和南天竹则表现为上方山区>植物园区>滨湖区,而香樟和银杏Nmass均在植物园区中最小,红枫反之。
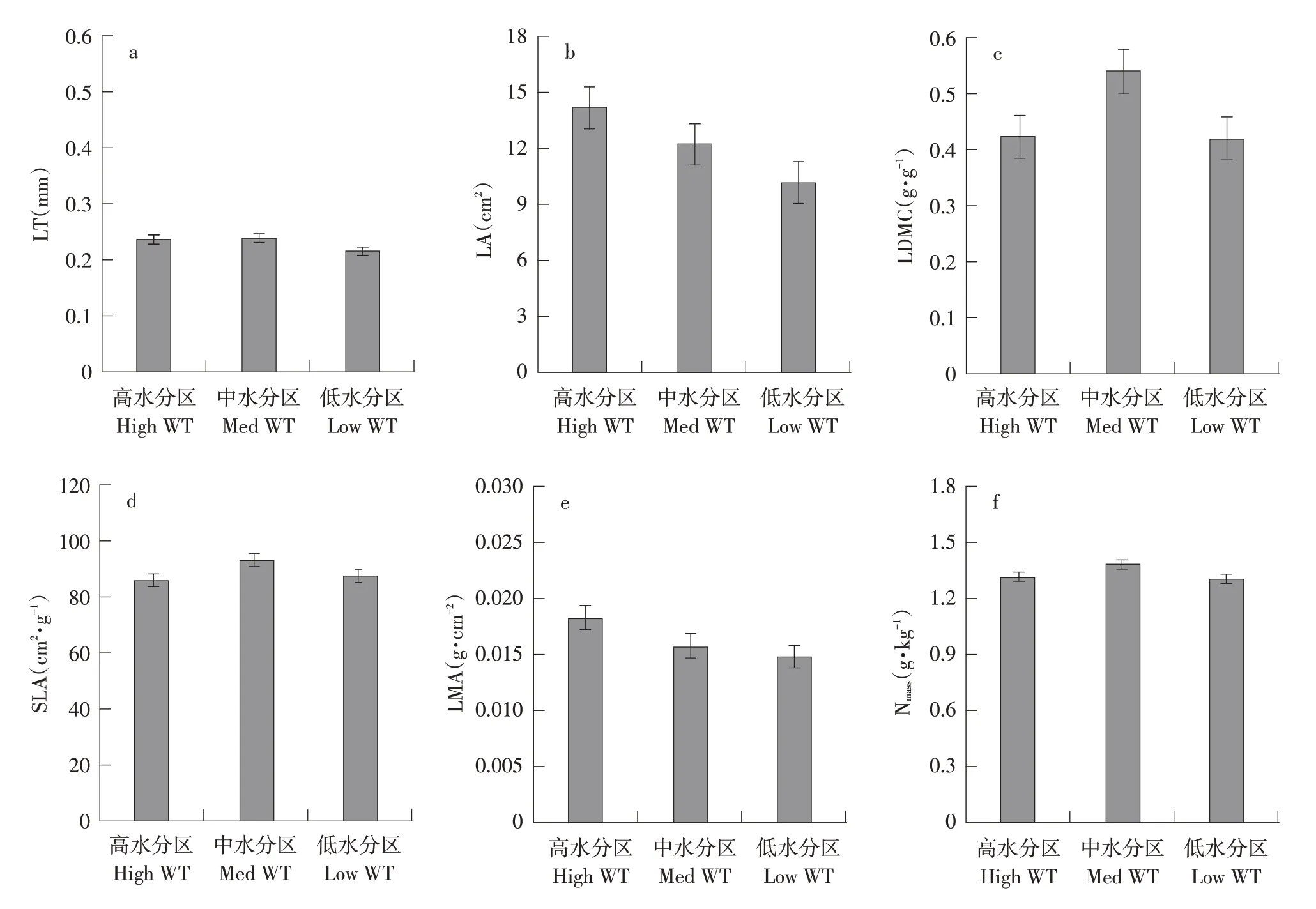
图1 主要叶经济谱性状特征对水分梯度响应Fig.1 Responses of main leaf economic spectrum traits to water gradient
另外,除桂花、红枫外,其余植物LA在石湖风景区中3个不同水分生境中均存在显著差异(P<0.05),表现相对一致响应,且LA指示叶片能量和水分平衡,植物叶片外在结构对其资源获取能力相关性较大,因此,LA也是该区域中植物的良好指示指标。香樟和石楠LA在3个不同水分生境中均表现为上方山区>滨湖区>植物园区,桂花和毛鹃LA则表现为上方山区>植物园区>滨湖区,表明这4种园林植物对石湖风景区不同水分生境响应一致。另外,LDMC也可指示植物对环境资源保持能力,较高LDMC通常具有较强抵抗环境胁迫能力[14]。银杏和南天竹LDMC在3个水分生境中表现为上方山区>植物园区>滨湖区,桂花和红枫表现为上方山区>滨湖区>植物园区,香樟和毛鹃表现为滨湖区>植物园区>上方山区,榔榆、紫薇和石楠则呈植物园区>滨湖区>上方山区趋势。SLA、LMA是反映植物对资源投资、利用、回报的叶经济谱性状[15]。桂花、香樟和紫薇SLA在石湖风景区3个水分梯度中表现为植物园区>滨湖区>上方山区,榔榆、石楠和南天竹则表现为上方山区>植物园区>滨湖区,红枫表现为滨湖区>植物园区>上方山区,毛鹃则表现为上方山区>滨湖区>植物园区,LMA均反之。
2.2 叶经济谱性状与水分环境相关性分析
在石湖风景区中,9种园林植物叶经济谱性状之间均表现出显著相关性(见表2)。其中,LT与LL之间存在极显著正相关(P<0.01);LDMC与LL、LW之间均存在显著负相关(P<0.05)。LA与LL、LW、LP间呈极显著正相关(P<0.01),与LT呈显著正相关(P<0.05),与前人研究表现出不一致响应。比叶面积和干物质含量是植物叶片最重要性状指标,以往研究表明,SLA与LDMC通常呈显著负相关,本研究中石湖风景区园林植物SLA与LDMC之间存在极显著负相关(P<0.01),再次论证前人研究,植物通过增加干物质含量,降低比叶面积降低植物叶片水分散失;LMA与LT、LDMC之间存在极显著正相关(P<0.01),与LW、SLA之间存在极显著负相关(P<0.01);Nmass与LW间呈显著负相关(P<0.05),与LT呈极显著负相关,与SLA呈显著正相关(P<0.05);植物叶片相对含水量和失水速率又是衡量植物保持水分能力常用性状指标,研究表明植物表现出LDMC值越大,植物叶片保水力越强的趋势。本研究中,石湖风景区园林植物RWC与LDMC、LA、LMA之间存在显著正相关(P<0.05),与LW呈极显著负相关(P<0.01);RWL与SLA呈显著负相关(P<0.05),而与LDMC之间正相关关系不显著;RWD与LDMC、LA、LMA之间存在显著负相关(P<0.05),与LW呈极显著正相关(P<0.01),与RWC呈极显著负相关(P<0.01)。

表2 石湖风景区植物叶经济谱性状间相关性分析Table 2 Correlation analysis among traits of plant leaf economic spectrum in Shihu Scenic Area
研究发现,在低水分区中,SLA与LDMC呈极显著负相关(P<0.01),与前人研究结果一致,植物通过增加干物质含量,减小比叶面积适应低水分生境(见图2)。另外,SLA还与Nmass呈极显著正相关(P<0.01),LW与RWC呈显著负相关(P<0.05),与RWD呈显著正相关(P<0.05),但在其余两个水分生境中相关性均不显著。庞志强等喀斯特研究表明,植物通过减小叶厚度,增大叶面积增加水分利用效率,但本研究中低水分区内LT和LA之间相关性不显著,且呈正相关关系,可能因人为干扰等综合因素导致的结果[16]。RWC与RWL是衡量植物叶片保水力常用指标,而植物叶片保持水分能力又是反映植物抵御干旱环境能力的一个重要性状[17]。本研究中低水分区园林植物RWC与LW呈显著负相关,RWL与SLA呈显著负相关,表明RWC与RWL增大同时,LW与SLA减小,这说明园林植物通过减小叶片大小提高植物叶片保水力,以此来适应低水分条件生活环境。中水分区中,RWC与LDMC之间呈显著正相关(P<0.05),叶片相对含水量是衡量植物叶片保水力常用指标,呈LDMC值越大,植物叶片保水力越强趋势。高水分区中,Nmass与LT之间呈极显著负相关(P<0.01),RWL与Nmass之间呈显著负相关(P<0.05),LMA、LDMC与LL、LW、LP之间均存在显著(P<0.05)或极显著负相关(P<0.01)。

图2 不同水分环境叶经济谱性状与生理性状间关系Fig.2 Relationship between leaf economic spectrum traits and physiological traits in different water environments
为确定不同园林植物主要叶经济谱性状对环境变化响应是否一致,本文计算不同水分生境园林植物平均性状值之间Pearson相关系数(见表3)。研究发现,低水分下LT与中水分下LT呈极显著正相关(P<0.01);中水分下LDMC与高水分下LDMC呈极显著正相关(P<0.01);中水分下SLA、LMA与高水分下SLA、LMA呈显著正相关(P<0.05);中水分下Nmass与高水分下Nmass呈显著正相关(P<0.05),低水分下Nmass与高水分下Nmass呈极显著正相关(P<0.01),低水分下Nmass也与中水分下Nmass呈显著正相关(P<0.05);而LA在高、中、低水分之间相关性均不显著。因此,在石湖风景区植物区系中,叶经济谱预测性状之间关系确实受到环境变化影响,但较弱。

表3 主要性状物种均值在不同水分区之间Pearson相关系数Table 3 Pearson correlation coefficients between species mean values of traits under different water conditions
2.3 叶经济谱性状与水分环境之间关联
由PCA分析(KMO=0.591,P<0.01),取特征值>1成分得到4个成分,解释总方差为80.4%。从图3A可知,前4个主成分包含所测叶经济谱性状大量信息,说明前4个主成分指标解释9种园林植物对环境适应能力。

图3 石湖风景区不同园林植物叶经济谱性状主成分分析Fig.3 Principal component analysis of leaf economic spectrum traits of different garden plants in Shihu Scenic Area
选取每个主成分中特征向量值绝对值>0.8为该成分主要指标,第1主成分包含主要性状指标为LL、LW、LP、LA,多是与叶片外在形态有关性状;第2主成分主要性状指标为RWC和RWD;第3主成分中主要性状指标为LT;第4主成分主要性状指标为LDMC。说明由主成分筛选出LL、LW、LP、LA、RWC、RWD、LT、LDMC这8个性状指标是影响石湖风景区园林植物环境适应能力主要指标,可用于评估园林植物对环境变化的适应能力。
2.4 植物环境适应性模糊隶属函数分析
植物对环境适应性综合评价隶属函数值常用来评价植物耐盐性、耐湿性等,从而比较各植物环境适应能力[6]。利用模糊隶属函数模型分析由主成分分析筛选出的8个主要性状指标,得到各性状指标环境适应性系数,最后根据平均隶属函数值判断园林植物环境适应性。
通过模糊隶属函数分析可知,在高水分区内9种园林植物8个主要指标综合平均隶属函数值分别为
0.4837、0.4151、0.4705、0.2946、0.4724、0.4397、0.5157、0.7526、0.2606,根据平均隶属函数值得知,9种园林植物在高水分区中对环境适应能力强弱依次为紫薇、石楠、桂花、香樟、红枫、银杏、榔榆、南天竹、毛鹃,说明在高水分区内紫薇对环境适应能力最强,毛鹃则最弱;9种园林植物8个主要指标综合平均隶属函数值在中水分区中分别为0.6903、0.5071、0.6594、0.2658、0.3826、0.6379、0.5654、0.3089、0.3406,根据平均隶属函数值可知,9种园林植物在中水分区中对环境适应能力强弱依次为桂花、红枫、银杏、石楠、榔榆、香樟、毛鹃、紫薇、南天竹,据此可知,桂花对该环境适应能力最强,南天竹则最弱;在低水分区内各指标综合平均隶属函数值分别为0.6467、0.2813、0.4856、0.1615、0.6498、0.6090、0.5589、0.4673、0.3584,根据平均隶属函数值可知,9种园林植物在低水分区中对环境适应能力强弱依次为香樟、桂花、银杏、石楠、红枫、紫薇、毛鹃、榔榆、南天竹,说明在低水分区内香樟对环境适应能力最强,而南天竹则最弱。
由图4可知,桂花和石楠在不同水分生境中环境适应能力均处于相对较强地位,而紫薇面对不同水分条件环境适应性差异较大。另外,由以上结果可知,在高水分区适应性较强的园林植物呈现LL、LT大,Nmass小的一致性特征,而其余性状差异较大;在中水分区适应性较强园林植物表现出LL、LW大,LDMC较小性状特征,其余性状对不同水分梯度响应则差异较大;而低水分区中LL、LW、LA较大的园林植物在环境中适应性较强。这些结果表明不同物种在同一环境变化中反应并不一致。因此,仅通过单个物种性状特征对环境变化的反应并不能准确预测环境中整体物种叶经济谱性状趋势。
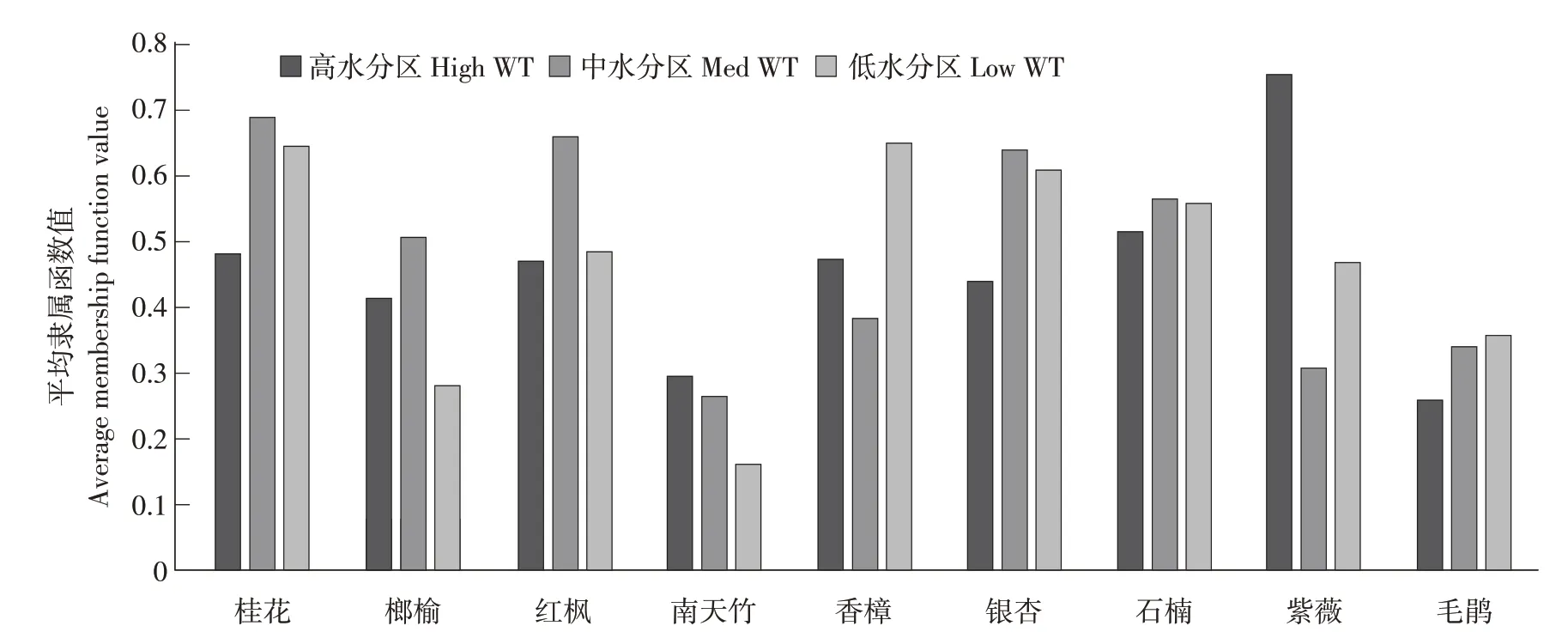
图4 园林植物不同水分条件下隶属函数值Fig.4 Membership function values of garden plants under different water conditions
3 讨论与结论
3.1 园林植物在不同水分生境中经济性状变化
不同园林植物在有限资源环境中,通过调整叶片或其他器官生理性状间权衡策略适应所处生境,最终达到和平衡“生存、生长、繁殖目的”[18-19]。因此,不同植物可通过各叶经济性状间权衡形成各自最佳组合模式,从而形成针对各个植物所处环境的生态适应策略,更好适应外界环境变化。
研究发现,当植物处于不同水分生境中,不同园林植物LP、LA及LL均存在显著差异,表明叶片大小在该区域中是反映水分环境响应的良好指标。低水分生境内LT均值较高水分生境高,植物通过增加其叶片厚度或密度降低植物内部水分损耗,印证Reich和Witkowski等研究结论[20-21]。植物在特定环境中通过保持其SLA相对稳定以适应环境变化[22]。在不同水分生境下,桂花、榔榆、南天竹和紫薇SLA存在显著差异,而其余植物SLA差异则不显著,对水分环境表现出不同响应。SLA低的植物一般是资源贫瘠环境中权衡的表现[23-24],本研究中银杏和红枫2种耐旱植物随水分胁迫增加,通过增加其SLA权衡策略来响应高水分环境,与李永华等干旱地区植物研究结论一致[25],但榔榆、石楠、南天竹3种耐旱植物SLA对水分环境变化权衡作用与之相反,因此不同耐旱植物SLA随水分环境变化规律还有待进一步论证。LDMC也与植物截取光资源和抗旱等能力密切相关,喻阳华等通过研究黔西北地区优势树种叶片功能性状与经济谱分析发现,干旱生境中植物通常具有较高干物质含量[26],与本研究结果一致。在低水分区内植物存在SLA降低、LDMC增加现象,表明当植物受制于恶劣水环境时,为降低叶片水分散失并最大化提升能量和物质使用效率,其叶经济谱性状形成这种权衡作用,用于提高保卫组织构造的筑建、增强抗逆能力。研究发现,桂花、南天竹、银杏、石楠和香樟Nmass在低水分区中更高,榔榆、紫薇和毛鹃Nmass则在高水分区中更高,与Wright[27]等和Pavón[28]等研究结论一致。另外,不同水分生境中红枫、紫薇、桂花和银杏等耐旱植物叶经济谱性状具有相似或不同相关关系,反映植物环境适应策略异同性。低水分区9种园林植物使LMA较低、Nmass较高,快速获取养分以适应环境;而LMA较高,Nmass较低时,表明植物在所处环境中会慢速获取养分。植物通过将有限氮资源转移至可抵抗环境胁迫的组织或结构中,应对外界环境胁迫,因此Nmass增加说明植物受到的环境压力加剧,此为环境适应的结果,也可能是响应优越环境的结果。
3.2 同一环境下不同园林植物调节方式
叶经济谱可将植物叶片性状间权衡作用数量化表现出来,不同园林植物在叶经济谱中所处位置均不同。叶经济谱一端是一类具有薄叶、比叶重小、叶氮含量高、光合速率大、呼吸速率快、叶寿命短的“快速投资-收益”型(Quick investmentreturn species)物种,而叶经济谱另一端是具有厚叶、比叶重大,寿命长,含氮量、光合速率和呼吸速率均偏低的“缓慢投资-收益”型(Slow investment-return species)物种。本研究中,香樟、南天竹、红枫LT、LMA显著低于其他植物,而Nmass显著高于其他植物,表明香樟、南天竹、红枫位于“快速投资-收益”型物种一端,紫薇、石楠、银杏、桂花位于“缓慢投资-收益”型物种一端,而榔榆偏向处于“快速投资-收益”型物种一端,毛鹃偏向处于“缓慢投资-收益”型物种一端。
在PCA分析中,第一个主要成分与叶片外部结构(如LL,LW,LP,LA等)具有高度相关性;第二主要成分与RWC和RWD具有高度相关性,且主要概括叶片水分状态,反映水分环境对植物的影响;第三主要成分与LT相关性较高,主要表明叶片厚度大小。第四主要成分与LDMC具有较高相关性,主要反映植物叶片结构和植物抗旱能力。因此,仅靠单一性状判断植物对环境响应机制不可靠,植物叶片结构性状、化学性状、生理性状等均与环境变化密切相关,说明植物可通过叶经济谱性状间权衡作用响应所处生境的环境因子变化。
3.3 园林植物对环境变化响应一致性
研究发现,不同物种和生境下植物叶经济谱性状之间差异均显著,且表现出相似或不同权衡关系,此结果与既往研究结论一致[29-30],表明植物叶经济谱性状指标差异是受其所处环境因子及自身遗传特性影响,反映植物在不同生境条件下对环境适应性。然而,尽管存在这些重要的相互作用,但各个性状特征之间反应通常在同一方向,仅变化幅度不同。因此,不同水分环境个体性状之间相关性普遍较显著。本研究中桂花在高水分下具有相对较低SLA,在低水分下仍具有相对较低SLA,表明物种水平对环境条件变化反应差异可能不足以改变物种在叶经济谱性状轴上相对位置。此外,本研究中主要叶经济谱性状值在高水分区与中水分区或中水分与低水分之间相关性相对较显著,高水分与低水分间相关性始终较低,说明水分条件微小变化可引起性状微小变化,但环境的较大变化在性状中产生较大且难以预测。虽然园林植物在物种水平对环境变化响应程度不同,但基于对植物群落有限抽样,特别是在小尺度范围群落中,预测广泛植物群落在全部环境梯度上性状特征值也许可行。另外,尽管跨广泛的环境梯度预测性状不准确,但与物种内性状变化相比,这些剧烈的环境变化可能更能引起群落物种组成变化。
当分析单个性状响应时,物种间响应一致性显著,而物种在多元性状间响应却不一致。即使植物群落性状表现出明显一致总体趋势情况下,也会出现个别植物与平均物种明显不同的性状变化情况,表明物种对环境变化总体特征反应存在特殊性。叶经济谱预测性状之间协调变化,本研究发现,石湖风景区中不同园林植物各自改变其叶经济谱能力差异较大,这种缺乏性状协调的表现可能还存在于其他植物群落中,需进一步研究确定这种模式的普遍性。本研究中叶经济谱性状之间关系在生境内部和生境之间表现出不同响应模式。通常共同生存于同一环境的物种应表现相似性状特征,但研究表明,在栖息地内,共存物种表现显著性状特征分散[31],这可能是因本地竞争也有助于选择具有更多差异、多元策略的植物,从而降低叶经济谱预测该区域群落内性状之间关系的能力,这些结果表明在利用植物叶经济谱性状预测群落和生态系统对环境驱动因素变化响应同时,也应重视种内性状变化过程。总而言之,不同研究尺度、环境因子、遗传作用等因素影响下形成的叶经济谱不同,因此,应用叶经济谱理论于研究中时,需充分考虑群落类型、自身发育、植物功能型与生活型、环境条件、干扰因子等因素。
综上,通过研究石湖风景区主要园林植物种间和种内差异发现,同一植物在不同生境中其叶经济谱性状间的权衡策略不完全相同。这可能是多种影响,如植物自身遗传变异、树木年龄和环境等因素,表明未来在研究叶经济谱性状间权衡策略时,应适当结合植物解剖结构特征、气孔密度等微观性状。同时,需测定土壤结构、容重、有机碳、全磷等指标,同时结合坡度、坡向、坡位等地形因子,从气象局官网等平台获取年均温、年均降雨量、年均蒸发量等数据[32],结合尺度转换方法和效应展开研究。另外,除水分条件外,不同水分生境中还存在其他环境因子,如气候因子、土壤成分、地形因子、土地使用方式等,这些环境因子及其与水分相互作用如何影响叶经济谱性状、植物如何适应生态因子间复杂的相互作用等问题需进一步研究。
猜你喜欢
现代农村科技(2022年7期)2022-11-18
东北林业大学学报(2022年1期)2022-03-28
种子(2022年2期)2022-03-21
新农业(2021年12期)2021-11-29
落叶果树(2021年6期)2021-02-12
绿色科技(2019年13期)2019-11-29
东坡赤壁诗词(2019年1期)2019-04-30
小学生优秀作文(低年级)(2018年11期)2018-11-14
农民致富之友(2017年12期)2018-01-31
体育科技(2016年2期)2016-02-28
