
南渡江上游海南半鲿生物学参数估算
2021-07-17 03:25:48梁智策李世启蔡杏伟申志新郭志强
水生生物学报 2021年4期
梁智策 刘 飞 李世启 常 通 蔡杏伟 申志新 郭志强*
(1. 南海海洋资源利用国家重点实验室, 海南大学海洋学院, 海南大学生命科学与药学院, 海口 570228;2. 海南省海洋与渔业科学院, 海口 571126)
海南半鲿(Hemibagrus hainanensisTchang 1935)为海南岛珍稀鱼类, 隶属于鲇形目(Siluriformes)、鲿科(Bagridae)、半鲿属(Hemibagrus)。其濒危等级为数据缺乏(DD)[1]。20世纪中期, 海南半鲿在海南岛南渡江水系、昌化江水系及万泉河水系均有分布[2]。近几十年来, 随着海南省社会和经济的不断发展, 人们活动对岛内南渡江、昌化江和万泉河等河流生态系统的干扰也在益日剧增, 如拦河筑坝、水域污染、过度捕捞、挖沙采石和外来物种放生等[3—5]。人类活动导致河流生态环境的一系列变化将会对鱼类产生一定的影响。
目前, 有关于海南半鲿的研究仍停留在形态学分类和分布记录等方面[2,6,7], 而对其基础生物学及资源动态方面的研究属于空白状态。有关海南半鲿的基础研究严重滞后, 相关野外实地调查数据缺乏, 导致无法对其受濒危程度进行评估[1]。因此, 开展海南半鲿个体生物学和种群生态学等方面研究,积累相关方面数据已迫在眉睫。鱼类生长和死亡参数是阐述鱼类生活史策略的主要指标[8], 它不仅是鱼类生物学以及生态学特征的研究基础, 同时也是鱼类种群结构变动趋势评估的重要依据[9,10], 因此开展海南半鲿生长特性和资源现状研究对于其种群资源保护和合理开发利用具有重要的理论和现实意义。
FISAT Ⅱ软件中的体长股分析法(LCA)具有无需鉴定研究对象年龄、数据结构简单和采样工作量小等优点, 被广泛应用于评估鱼类生长参数与资源量[11]。本研究以南渡江上游海南半鲿为研究对象, 通过评估海南半鲿的体长组成、体长-体质量关系及种群参数, 以期掌握目前南渡江上游海南半鲿生长特性及其资源利用现状, 旨在为海南岛珍稀海南半鲿的种群资源恢复和保护提供理论依据, 从而实现南渡江上游海南半鲿资源的可持续发展。
1 材料与方法
1.1 研究区域与数据获取
南渡江为海南岛最大的河流, 发源于白沙黎族自治县南峰山, 干流斜贯海南岛中北部, 于海口市美兰区流入琼中海峡。南渡江上游山高坡陡, 支流众多, 生境多样性较高, 具有丰富的鱼类种类, 是鱼类良好的栖息地[12]。根据历史资料记载[2,13,14],20世纪80年代南渡江共记录有鱼类152种。2018年余梵冬等[12]调查南渡江鱼类种类组成及分布, 其结果表明南渡江鱼类共计136种, 保存相对完整。
本研究于2019年1—12月对南渡江上游渔获物按月度进行调查(图1)。每月15—17号在日落之前放置3条多网目复合刺网(长: 30 m; 高: 1.5 m; 网目(2a/cm)分别为: 1.0、6.0和12.5)和地笼(长: 4 m; 网目: 2a=0.4 cm), 次日6:00收网, 收集网内所有的渔获物, 并及时放入带有冰块的鱼箱内保存, 带回实验室进行体长(精确到1 mm)和体质量(精确到0.01 g)等生物学参数的测定。
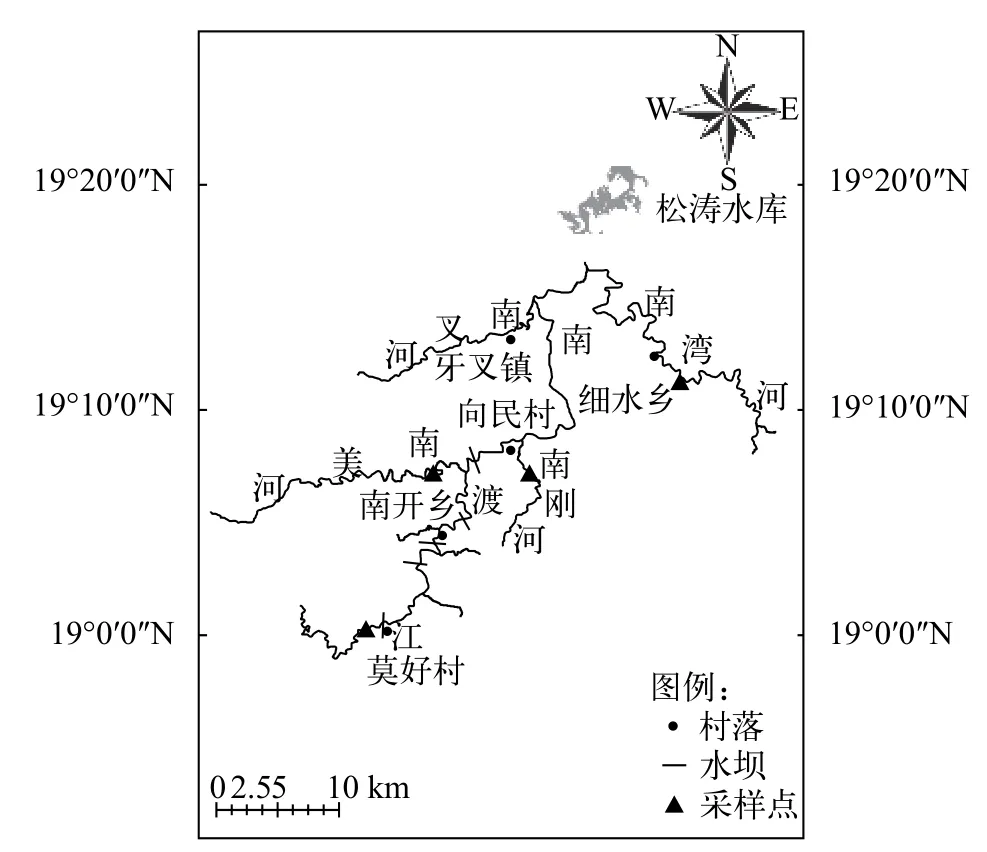
图1 南渡江上游海南半鲿采样点布置图Fig. 1 Sampling stations of H. hainanensis in the upper Nandu River
1.2 研究方法
体长与体质量关系其体长与体质量关系式W=aLb由幂函数拟合, 式中W为体质量(g);a为肥满度;L为体长(mm);b为幂指数系数。
生物学参数海南半鲿的体长以10 mm为组距[15], 分为25组, 依次是40—49 mm、50—59 mm、······、260—269 mm和270—279 mm。输入海南半鲿体长频率数据, 利用FiSAT Ⅱ软件中的ELEFANⅠ法估算其渐近体长(L∞)和生长系数(K)[16]。并利用Pauly[17]经验公式推算其理论生长起点年龄t0:
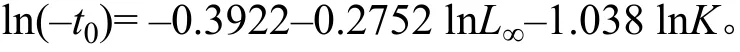
另外, 采用体长变换渔获曲线法估算总死亡系数(Z), 自然死亡系数(M)通过其自然死亡系数估算模块进行估算[15,18]。
生长特性指标运用von Bertalanffy生长方程[19], 并利用Lt(t龄时的预测体长)、t0(理论生长起点年龄)、Wt(t龄时的预测体质量)、W∞(渐进体质量)等参数, 拟合南渡江上游海南半鲿的体长、体质量生长方程:

对上述生长方程中t进行一阶、二阶求导, 可获得其体长、体质量的生长速度及加速度曲线方程, 分别为:

分析单位补充量渔获物量(1)估算平均选择体长。线性回归中未被利用各点的相对期望值ln (N/Δt)通过由体长变换曲线拟合而成的直线方程进行估算, 进而得到未被利用各点观察与期望值两者的比值及其累积率, 开捕体长(Lc)为累积率达到50.0%时所对应的体长。(2)分析相对单位补充量渔获量。Beverton和Holt在1957年提出了动态综合模式(Dynamic pooled model)来解释鱼类种群参数和渔获量的关系[20]。当公式中参数发生变化时, 便可得到渔获量(Y)及单位补充量渔获量(Y′/R)的相应变化情况, 也称Beverton-Holt (B-H)稳定状态模型[20]。Pauly和Soriano[21]利用动态综合模式来预测Y′/R和B′/R(相对单位补充量生物量)。Y′/R和B′/R曲线通过刀刃式选择假设模型(Knife-edge model)建立, 数学公式如下:

FiSAT Ⅱ软件中开发率(E)一般有下述几种水平: (1)Emax为获得最大渔获量时的开发率; (2)E0.1为Y′/R边际增长减少10.0%时的开发率; (3)E0.5为资源量下降到原始水平50.0%时的开发率。
2 结果
2.1 体长分布
南渡江上游共采集海南半鲿358尾, 其体长为40—290 mm, 平均体长为143.7 mm, 其中优势体长集中在80—180 mm, 占总个体数的77.1%, 各体长组分布见图2。
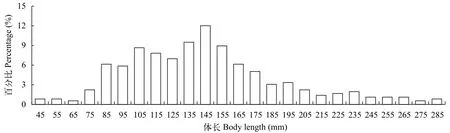
图2 南渡江上游海南半鲿体长分布Fig. 2 The distributions of the body length of H. hainanensis in the upper Nandu River
2.2 体长与体质量关系
通过拟合南渡江上游海南半鲿体长(L, mm)和体质量(W, g)的幂指数曲线, 得到其体长和体质量关系回归方程为:
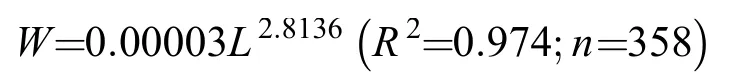
即其幂指数值为2.8136, 接近3 (P>0.05), 为等速生长(图3)。
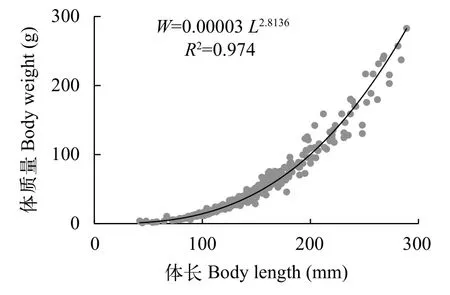
图3 南渡江上游海南半鲿体长与体质量关系式Fig. 3 Length-weight relationships of H. hainanensis in the upper Nandu River
2.3 生长参数与生长方程
海南半鲿为等速生长鱼类, 故本研究运用von Bertalantly生长方程描述其生长规律。运行FISATⅡ软件中ELEFAN Ⅰ法的K扫描模块可获得渐近体长(L∞)为304.5 mm, 生长参数(K)为0.49。再根据Pauly[17]提出的计算理论生长起点年龄的经验公式求出t0为–0.29。因此其von Bertalantly体长和体质量生长方程分别为:

南渡江上游海南半鲿年龄与体长、体质量2条相关曲线均为不具拐点的上升曲线(图4)。
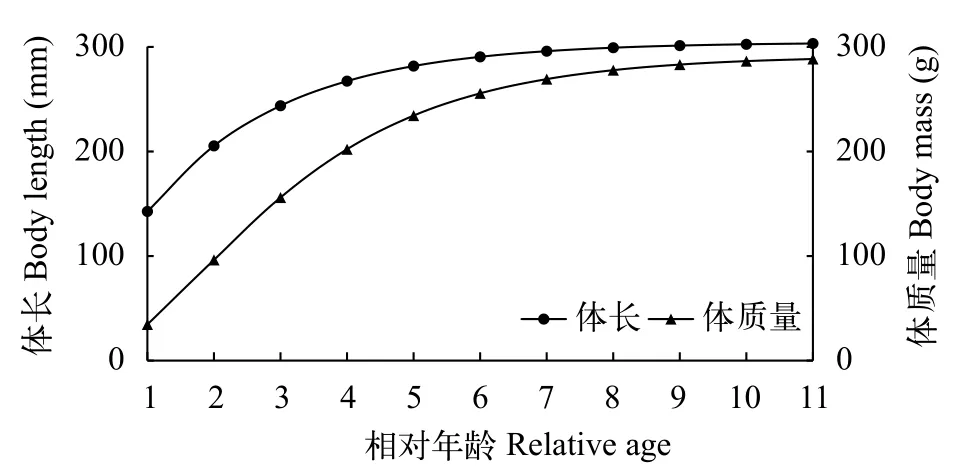
图4 南渡江上游海南半鲿von Bertalantly生长曲线Fig. 4 Von Bertalantly growth curve of H. hainanensis in the upper Nandu River
运用体长变换渔获物曲线法对南渡江上游海南半鲿总死亡系数(Z)进行估算, 其拟合的直线方程为(图5):
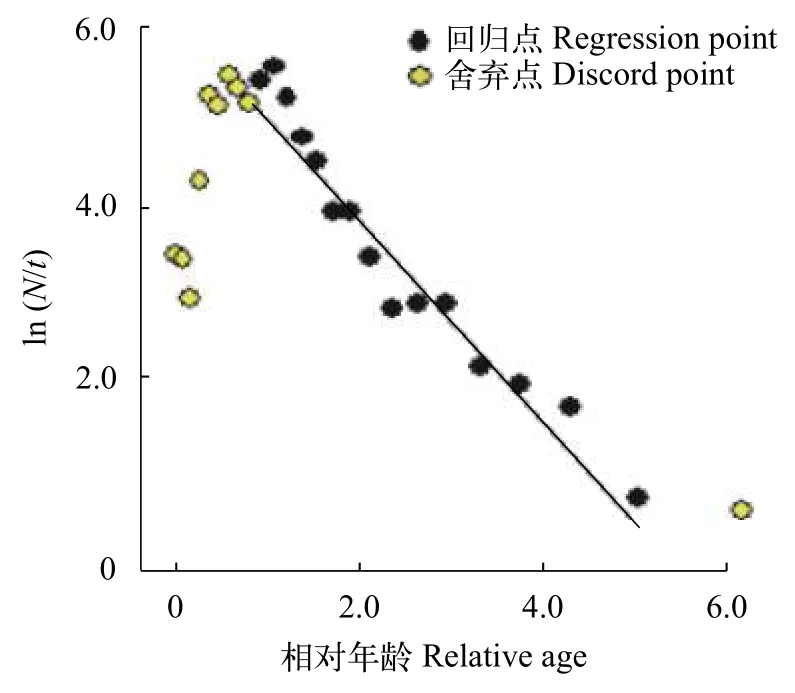
图5 体长变换渔获曲线估算南渡江上游海南半鲿总死亡系数(Z)Fig. 5 Estimation of total mortality parameter (Z) from lengthconverted catch curve for H. hainanensis in the upper Nandu River

式中,N为数量,t为相对年龄。
该拟合的直线方程斜系数为–1.22, 其95%置信区间为–1.40 — –1.05。南渡江上游海南半鲿的总死亡系数(Z)为1.22, 其95%置信区间为1.05—1.40。
将南渡江上游海南半鲿的渐近体长(L∞)为304.5 mm、生长参数(K)为0.49及T为24.8℃(本研究采用南渡江上游年均水温), 代入Pauly[18]经验公式可得: 南渡江上游海南半鲿的自然死亡系数(M)为1.05, 总死亡系数(Z)为1.22, 捕捞死亡系数(F)为0.17; 由E=F/Z得出南渡江上游海南半鲿的开发系数E=0.14。
2.4 生长特性与拐点年龄
鱼类体长和体质量的生长速度和生长加速度用于描述其生长过程变化特征。南渡江上游海南半鲿体长生长速度和加速度曲线方程(图6)分别为:
体长生长速度: dLt/dt=149.21e–0.49(t+0.29)
体长生长加速度: d2Lt/dt2= –73.11e–0.49(t+0.29)
体质量生长速度和加速度方程(图7)分别为:
体质量生长速度: dWt/dt=428.80 e–0.49(t+0.29)·[1–e–0.49(t+0.29)]2
体质量生长加速度: d2Wt/dt2=210.11 e–0.49(t+0.29)·(1–e–0.49(t+0.29))·(3e–0.49(t+0.29)–1)
图6表明, 海南半鲿体长生长速度随年龄的增加而递减, 成负相关关系, 至高龄时逐渐趋近于0。图7表明, 海南半鲿体质量的生长速度先增大后减小, 体质量的生长速度在2龄之前随年龄增加而增大, 最大值出现在1—2龄, 拐点之后, 体质量的生长速度随年龄的增加而减小, 至高龄时则逐渐趋近于0。海南半鲿的生长拐点年龄为1.95, 对应拐点体长和体质量分别为202.9 mm、93.08 g。
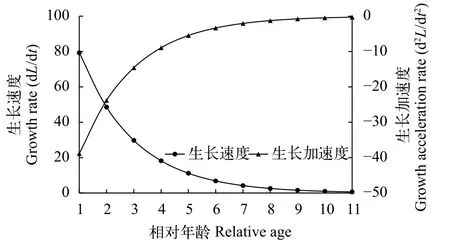
图6 南渡江上游海南半鲿体长生长速度和加速度变化曲线Fig. 6 Growth rate and acceleration rate in body length of H.hainanensis in the upper Nandu River
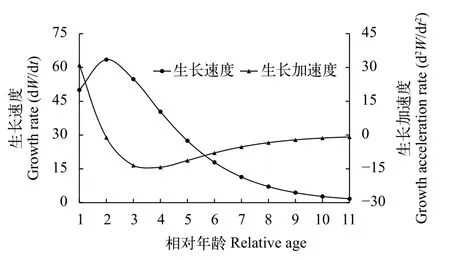
图7 南渡江上游海南半鲿体质量生长速度和加速度变化曲线Fig. 7 Growth rate and acceleration rate of body mass of H.hainanensis in the upper Nandu River
2.5 分析单位补充量渔获量
估算平均选择体长当累积率达到50%时对应的体长为(Lc)为146.5 mm, 与开捕体长(Lc)相对应的年龄(tc)为1.05。
分析相对单位补充量渔获物量采用FiSATⅡ软件分析南渡江上游海南半鲿的开发率(E)与相对单位补充量渔获量(Y′/R)的关系(图8)。其中,M/K=2.14,Lc/L∞=0.48, 估算出Emax=0.78,E0.1=0.67,E0.5=0.36。
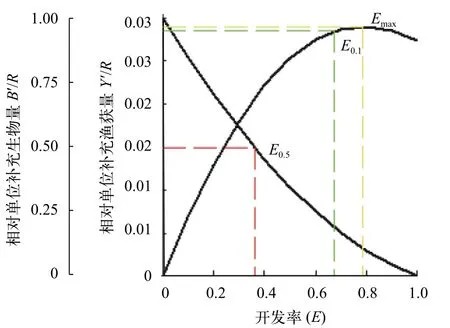
图8 南渡江上游海南半鲿相对单位补充量渔获量、生物量与开发率的关系Fig. 8 Relationship between relative yield-per-recruit (Y′/R),biomass-per-recruit (B′/R) and exploitation ratio (E) of H.hainanensis in the upper Nandu River
通过分析相对单位补充量等值曲线, 阐明Y′/R与Lc和E之间的动态变化趋势(图9)。
在图9中, 颜色由蓝色到红色表示数值逐渐由低到高依次增加。通常将渔获量等值曲线以等值线平面圆点分四个象限, 分别为左上区域(A)、左下区域(B)、右上区域(C)和右下区域(D)[22]。在图9中, 当前南渡江上游海南半鲿渔业点为A点, 且开发率(E)为0.14,Lc/L∞为0.42[即开捕年龄(tc)为1.05、开捕体长(Lc)为146.5 mm]位于等值曲线的B象限,表明南渡江上游海南半鲿的渔业资源未处于过度开发状态。
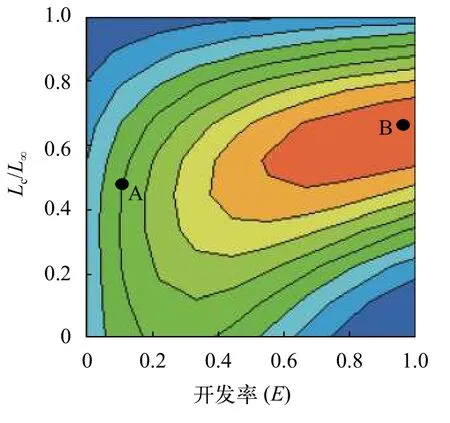
图9 南渡江上游海南半鲿相对单位补充量与开发率和开捕体长的关系(A. 渔业点; B. 最佳开发状态)Fig. 9 Relationships between relative yield-per-recruit (Y′/R) and exploitation ratio (E), standard length at first catch (Lc) of H.hainanensis in the upper Nandu River
在图9中, 南渡江上游海南半鲿逻辑上的最佳开发状态为B点。在M/K为2.14不变的情况下,Y′/R达到最大值0.03, 与其对应的E约为1.00,Lc/L∞约为0.70。若从A点调到B点,Y′/R将达到最大值, 其对应的开发率E增幅为614.3%及Lc/L∞增幅为67.5%, 而开发率的增加则对应着捕捞强度的增大。
当E为0.14,M/K为2.14时, 提高开捕体长Lc(在Lc/L∞>0.42的前提下)时,Y′/R与Lc/L∞呈正相关, 当Lc/L∞提升至0.50时(Lc为152.3 mm,tc为1.13),Y′/R则会达到相应的最大值0.01, 之后Lc/L∞与Y′/R呈负相关。与采捕时的开捕体长(Lc=146.5 mm)相比, 当开捕体长提升至152.3 mm时, 其对应的Y′/R并无变化,均为0.01。另外, 当E<0.14时,Y′/R随着开发率E的增加而增加; 当Lc=146.5 mm,E>0.14时, 随着开发率E的增加,Y′/R呈先增大后减小的趋势。当开发率E增大为E0.5(0.36)时, 对应的Y′/R≈0.02; 当开发率E继续增大为E0.1(0.67)时, 对应的Y′/R值增加, 即Y′/R≈0.02; 当开发率E继续增大为Emax(0.78)时, 对应的Y′/R值减小, 即Y′/R≈0.02。
从维持合理开发利用生物资源量来说, 当Emax为0.78时, 仅约10.0%的生物量资源得到保持;而当E0.5为0.36时, 则有约50.0%的生物量资源可得到保持。
3 讨论
3.1 生物学参数
利用体长频率数据对鱼类生长和死亡参数进行估算, 避免了年龄鉴定的主观误差及其繁琐工作,该方法已在渔业资源评估中得到了广泛应用并取得了良好的应用效果[22—24]。海南半鲿体表无鳞片,若以耳石作为年龄鉴定的材料, 其操作过程繁琐,技术性要求高, 且周期性较长[24]。因此, 本研究采用体长股分析法(LCA)评估南渡江上游海南半鲿的生长特性和死亡参数。
研究中模型本身适用性和数据误差会影响模型分析结果的准确度。当样本量(大个体样本)和采样月份足够多时, 即可保证模型分析结果的准确度。本研究使用地笼和多网目复合刺网的采样方式, 所捕获样本体长分布合理, 确保了模型分析结果的准确度。一般认为, 自然死亡系数(M)受鱼类生长状况、栖息环境等因素的影响[25]。在本研究中, 南渡江上游海南半鲿M为1.05, 与其他鲿科鱼类相比, 处于较高水平[26,27]。这表明海南半鲿的死亡多是由被捕食和栖息环境变化等自然因素所造成的, 受人为捕捞等因素干扰较小。K值表示鱼类的生长系数, 规定了曲线接近渐进值的速率[28]。von Bertalanffy生长方程表明,K值和鱼类渐近体长L∞、渐进体质量W∞呈负相关, 与M呈正相关。当K满足e–k<1时, 说明von Bertalanffy生长方程能较好地拟合该鱼类的生长[29]。在本研究中K=0.49, e–k=0.61<1, 符合理论要求。评估模型及数据的误差会影响研究结果的准确性, 当M/K的值处于1.50—2.50时, 说明所求得的M值是合理的[30];Z/K≤3时,表明自然死亡为种群死亡的主要原因, 而当Z/K>3时, 则表明捕捞死亡为其主要原因[15]。在本研究中M/K=2.14,Z/K=2.49, 表明估算的M值是合理的, 且海南半鲿的种群死亡主要原因为自然因素所造成的死亡。本研究分析当前南渡江上游海南半鲿的资源开发率(E)为0.14。根据Peter[31]提出资源最适开发率水平为0.50, 表明海南半鲿的资源未遭过度开发; Mehanna[32]认为Y′/R与E关系曲线中,Emax的左侧区域代表渔业资源能得以可持续发展的安全区域, 本研究中南渡江上游海南半鲿的开发率为0.14位于Emax(0.78)的左侧区域, 表明海南半鲿资源尚可持续发展。结合各项指标综合分析, 海南半鲿的资源未处在过度开发状态。
3.2 资源利用与物种保护
渔业资源利用量与多种因素有关, 例如: 鱼类种群大小、开捕年龄及捕捞强度等, 同时, 这些因素也影响着渔业资源状态[33]。可通过调节渔业管理的2个重要途径(捕捞强度(捕捞死亡系数)和初次捕捞年龄(开捕体长))来改变渔业资源的利用量, 以期获得最大可持续渔产量[15]。本研究通过Beverton-Holt动态综合模型分析, 当前南渡江上游海南半鲿渔业利用状态较好, 未遭过度捕捞。若维持当前开发率, 增大开捕体长(Lc)至152.3 mm时(tc为1.13),Y′/R达到最大值为0.01,Y′/R值并无改变; 由图9可见, 增加当前开发率至E0.1, 对应的Y′/R可增大64.3%。两者相比较, 若为优化资源利用状态, 增大开捕体长对捕捞产量的影响较小, 合理适当地增加捕捞强度对提高捕捞产量影响较明显。因此, 建议有关渔业部门合理地控制好捕捞网具的网目尺寸以及捕捞强度, 以期维持南渡江上游海南半鲿的种群资源量。
在我国内陆捕捞渔业中, 渔业生产可持续发展面临的重要威胁为捕捞强度过大, 同时也是渔业行政部门重点监管的一部分[33,34]。目前, 南渡江保存了历史大部分鱼类, 河流整体健康状况良好, 但鱼类总种数相较20世纪80年代下降了10.5%[4]。本次调查发现威胁南渡江上游海南半鲿种群生存的因素可能有以下几方面: (1)近几十年来, 海南岛槟榔种植业的快速发展, 导致种植区水土流失严重[35],同时农业生产活动中的常用农药会对鱼类产生强烈的毒害作用, 导致其行为、生化指标及血液指标异常[36], 并且对鱼类胚胎发育产生严重的毒害作用[37,38]。(2)水利工程建设是威胁海南岛淡水鱼类多样性的最大因素[39], 南渡江上游建有18个小型滚水坝, 可能造成海南半鲿生境碎片化, 影响其产卵、摄食场所, 降低种群整体的遗传多样性。(3)外来鱼类入侵是导致本土鱼类种群数量下降的最大因素[40], 南渡江上游尼罗罗非鱼(Oreochromis niloticus)和马拉瓜丽体鱼(Cichlasoma managuense)等外来入侵物种的大量出现[12], 会通过资源竞争,捕食鱼卵及子代等多种途径对海南半鲿的生存造成威胁[41]。尽管目前海南半鲿资源未遭过度捕捞,但其野外种群数量稀少, 如不及时采取有效的、科学的保护措施对其进行保护, 其将会面临灭绝的威胁。为预防海南半鲿资源进一步衰减, 建议有关部门采取以下措施: (1)建立鱼类自然保护区, 加强海南岛珍稀鱼类——海南半鲿的种群动态监测和保护。(2)及早开展海南半鲿生活史策略调查研究, 为制定种群保护策略提供充分的科学依据。(3)加强水域生态环境健康监测, 开展海南半鲿人工增养殖技术研究, 重视海南半鲿野外生境保护及放流效果评估。
猜你喜欢
今日农业(2022年15期)2022-09-20 06:54:16
天天爱科学(2022年9期)2022-09-15 01:12:28
海洋信息技术与应用(2022年1期)2022-06-05 07:38:24
科普童话·百科探秘(2022年8期)2022-05-30 10:48:04
昆明医科大学学报(2022年1期)2022-02-28 07:46:32
红土地(2018年7期)2018-09-26 03:07:38
探索科学(2017年5期)2017-06-21 21:16:16
金色少年(奇趣科普)(2016年8期)2016-09-21 02:04:58
小星星·阅读100分(低年级)(2014年5期)2014-06-24 00:42:45
中国海洋大学学报(自然科学版)(2014年9期)2014-02-28 12:21:38
