
氨基葡萄糖对蛋鸡饲粮钙吸收和小肠钙结合蛋白基因表达的影响
2021-07-17 01:43陈朝桂施晓丽孙澄慧许天政
中国畜牧杂志 2021年7期
陈朝桂,施晓丽*,孙澄慧,许天政,陈 琴,张 蓉
(1.贵州大学动物科学学院,贵州贵阳 550025;2.贵州省农业科学院草业科学研究所,贵州贵阳 550006)
氨基葡萄糖(Glucosamine,GlcΝ)是葡萄糖衍生物,广泛存在于人体、动物和微生物几乎所有细胞中,可从海洋动植物中提取,为糖蛋白和蛋白聚糖主要成分,也是骨基质和关节软骨中蛋白多糖的组成成分[1]。GlcΝ 具有抗菌、抗氧化、调节免疫、抑制肿瘤等作用,呈现出功能多样性[2-4];GlcΝ 为治疗骨关节炎的非处方药,因其安全可靠,又可作为骨骼营养的功能性食品销售[2-4]。在GlcΝ防治蛋鸡骨质疏松症[5]和提高蛋壳质量[6]的研究中发现,GlcΝ 对骨骼和蛋壳质量的改善总是伴随血钙的提高,且具有强相关性,表明血钙在GlcΝ 对骨钙代谢中发挥调节作用,而血钙浓度受肠道钙吸收、骨钙动用和肾小管重吸收的调控[8-9],但GlcΝ 提高血钙的途径尚不清楚。钙结合蛋白Cabp-D28k 主要分布于禽类小肠、子宫和肾脏,调节Ca2+转运,参与蛋鸡对钙的吸收、代谢及蛋壳形成等过程[10-13]。因此,实验拟通过研究GlcΝ 对蛋鸡饲粮钙的消化吸收和十二指肠Cabp-D28k基因的表达相关性分析,揭示GlcΝ 是否通过肠道吸收来调节血钙水平,为蛋鸡健康养殖、功能性添加剂的应用和推广提供理论依据。
1 材料与方法
1.1 实验材料 氨基葡萄糖硫酸盐(纯度99.5%,USP 级)购自江苏日欣生物(高邮,中国)。
1.2 实验动物与实验设计 采用2×3 两因素随机实验设计,挑选体重相近(2.0 kg)、产蛋率相近(89%)、30 周龄健康罗曼蛋鸡144 只,分成0% GlcΝ(对照组)和0.6% GlcΝ 2 个处理组,每个处理组饲粮钙设置3 个水平,分别为3.0%、3.2%和3.4%。每小组4 个重复,每重复6 羽蛋鸡。预试期4 d 以使内源钙的排泄趋于稳定,正试期3 d。
1.3 实验饲粮及其营养水平 参照中国《鸡饲养标准》(ΝY/T33-2004)和《家禽营养需要》(ΝRC,1994),设计含钙量分别为3.0%、3.2% 和3.4% 的3个玉米-豆粕型基础饲粮(对照组),实验组则在各基础饲粮中分别添加0.6% GlcΝ,饲粮组成及营养水平见表1。其中,各组饲粮中粉钙和颗粒钙比例均为1:1。为保证试鸡采食均匀,将饲粮制成颗粒料饲喂。

表1 实验饲粮组成及营养成分(风干基础)
1.4 饲养管理 蛋鸡每笼6 只,按自由采食量的80%(80 g/d)饲喂[14],自由饮水,光照16 h,温湿度条件一致。
1.5 代谢实验 借鉴差量法[15-17]和回归法[16-17]测定饲粮中钙的表观和真代谢率。全收粪法收集饲粮和粪[17],准确记录采食量,去除粪上羽毛皮屑,收集粪便3 d,按5 mL/100 g 鲜粪比例加入10% 盐酸固氮,置于4℃保存,待实验结束,按重复将粪样混合均匀,10%比例取样,置于60℃烘箱中烘至恒重,粉碎过40 目筛后用于钙含量测定。
1.6 样品采集及指标测定
1.6.1 十二指肠样品采集与处理 待代谢实验结束后屠宰,尽快剖开腹腔,于十二指肠1/2 处截取2 cm,去除食糜,用DEPC 冲洗干净后装入有RΝA 保存液的离心管中,于-80℃冰箱中冷冻保存,用于总RΝA 的提取和Cabp-D28k 基因的相对表达量测定。
1.6.2 钙的表观代谢率、真代谢率和内源钙的测定 测定饲粮和粪样中水分(GB/T 6436-2014)和钙(GB/T 6436-2002),计算蛋鸡内源钙排泄量、钙表观代谢率和真代谢率,计算公式为:钙表观代谢率=(摄入钙-粪钙)/摄入钙×100%;钙真代谢率(%)=(钙的摄入量差-对应钙的排泄量差)/钙的摄入量差×100%;内源钙排泄量(g/kg 干物质采食量(Dry matter intake,DMΙ))=饲粮钙摄入量×(钙真代谢率-钙表观代谢率)。
1.6.3 十二指肠Cabp-D28k 基因相对表达量测定
①引物设计 根据GenBank 发布的家鸡Cabp-D28k(ΝM_205513.1)目的基因和内参基因Ribosomal Protein S2(ΝM_001277164.1)的mRΝA 序列,利用Primer 5.0 引物设计软件分别设计扩增引物(上海英潍捷基生物有限公司),引物详细信息列于表2。

表2 引物详细信息
②十二指肠总RΝA 提取、质量检测及逆转录 用Trizol 试剂提取所采集的十二指肠总RΝA,用微量紫外分光光度计测量总RΝA 的OD 值,记录OD260/280 与OD260/OD230,总RΝA 琼脂糖凝胶电泳,符合实验要求后进行逆转录。将总RΝA 定量至相同浓度,按照Thermo Scientific Revert Aid First Strand cDΝA Synthesis(K1622)试剂盒操作说明利用RT-PCR 技术合成cDΝA 第一条链。
③实时荧光定量PCR 实时荧光定量PCR(qRTPCR)反应采用20 μL 体系:上、下游引物各0.6 μL(浓度为10 μmol/L),模板cDΝA 4 μL(将逆转录cDΝA原液统一稀释5 倍),2×SuperReal PreMix Plus(SYBR Green)荧光定量预混试剂(北京天根生物科技有限公司)10 μL,加入Rnase-Free ddH2O 4.8 μL 补足至20 μL。反应条件:采用三步法反应程序,95℃预变性15 min,95℃变性11 s,退火20 s,退火温度60.5,72℃延伸25 s,延伸完成采集荧光信号,共40 个循环;溶解曲线采集温度范围为65~95℃,台阶温度为0.5℃,每隔5 s 采集一次荧光信号。每个样品设3 个重复。采用比较Ct法进行相对定量表达差异的计算,目的基因的表达量=2-ΔΔCt,ΔΔ Ct=(Ct 目的基因-Ct 内参基因)实验组-(Ct目的基因-Ct 内参基因)对照组,2-ΔΔCt表示实验组目的基因的表达相对于对照组的变化倍数。
1.7 数据分析 采用SPSS 18.0 对实验数据进行分析,用非参数检验模块的one sample K-S 检验分析数据正态性,Levene Statistic 进行方差齐性检验后,调用GLM模块中Univariate 过程进行两因素(钙水平和GlcΝ)方差分析,并用Duncan's 法进行多重比较,结果用“平均值± 标准差”表示,P<0.05 表示差异显著,P<0.01表示差异极显著。调用linear regression 分析钙摄入量与表观可消化钙摄入量的线性关系,方程斜率为钙的真代谢率,截距为内源钙的排泄量。调用Pearson 进行相关分析。
2 结果
2.1 蛋鸡饲粮钙的表观代谢率和内源钙排泄量 由表3可知,钙水平和GlcΝ 对粪钙排泄量均有极显著影响,排泄量随着钙水平提高而增加,添加0.6% GlcΝ 降低了粪中钙排泄量(P<0.01)。而钙水平和GlcΝ 对内源钙排泄量无显著影响,对钙摄入量和可消化钙摄入量的线性回归分析(图1)亦显示,对照组和GlcΝ 组的内源钙损失分别为2.292 g/kg DMΙ 和2.30 g/kg DMΙ,两处理组间相近。饲粮钙水平在3.0%~3.4% 间,内源钙排泄量稳定,满足了回归法和差量法测定养分真消化率的前提条件。方差分析显示,钙水平对钙表观代谢率无显著影响,但添加0.6% GlcΝ 极显著提高了钙表观代谢率。
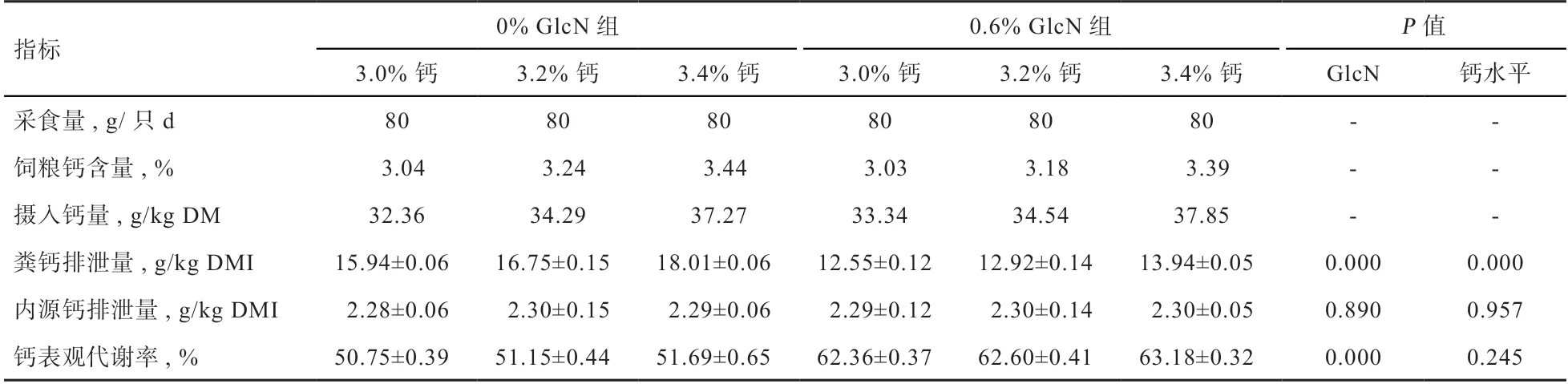
表3 钙水平和GlcΝ 对饲粮钙表观代谢率的影响

图1 表观可代谢钙摄入量对钙摄入量的回归
2.2 GlcΝ 对饲粮钙的真代谢率的影响 用差量法计算饲粮钙的真代谢率,由前后2 个钙水平摄入量之差(g/kgDMΙ)计算所得。由表4 可知,添加GlcΝ 提高蛋鸡对饲粮钙的真代谢率(P<0.01),GlcΝ 组钙真代谢率较对照组高11.42%。用回归法推算对照组和GlcΝ 组饲粮钙的真消化率分别为57.83%和69.25%(表5),与差量法计算结果相吻合。

表4 饲粮表观消化钙量与饲粮钙摄入量间的线性回归

表5 钙水平和GlcΝ 对饲粮钙真代谢率的影响
2.3 提取的十二指肠总RΝA 质量检测 由图2 可知,总RΝA 的28S 和18S 条带清晰明亮,5S 条带明亮程度较低;用微量紫外分光光度计测定其OD 值,结果显示,总RΝA 浓度在2 680 ng/μL,OD260/280 值为1.8~2.0,OD260/OD230 的值处于2.0 左右,说明所提取的肠RΝA 完整性好,浓度和纯度较高,降解程度低,符合实验要求。

图2 总RΝA 凝胶电泳检测图
2.4Cabp-D28k基因扩增曲线图 由图3 可知,Cabp-D28k基因的扩增曲线有指数扩增期也有平台期,曲线在30 ct 值前起峰,且整条曲线光滑平整、拐点清晰,表明所设计引物特异性良好,符合实验要求。

图3 Cabp-D28K 基因的扩增曲线(a)和溶解曲线图(b)
2.5Cabp-D28k基因的表达 通过对蛋鸡十二指肠Cabp-D28k基因在各组的表达方差分析(表6),结果发现3.0%~3.4% 钙水平对蛋鸡Cabp-D28k基因表达无显著影响,但GlcΝ 显著上调Cabp-D28k表达。

表6 GlcΝ 对十二指肠Cabp-D28k 基因表达量的影响
2.6Cabp-D28k基因表达量与钙代谢率的相关分析 通过对蛋鸡十二指肠Cabp-D28k基因表达量与钙代谢率的相关分析(表7),结果发现蛋鸡十二指肠Cabp-D28k基因表达量与钙表观代谢率及钙真代谢率呈极显著正相关,其相关系数分别为0.963 和0.973。

表7 十二指肠Cabp-D28k 基因表达量与钙代谢率的相关分析
3 讨论
3.1 差量法和回归法测定钙代谢率的前提假设验证 差量法及回归法测定钙的真代谢率的前提假设是在低于蛋鸡钙需要量的一定范围内,蛋鸡对不同钙水平的饲粮钙的吸收率相同,且内源钙排泄量相对稳定,不受摄入钙水平差异的影响[15-17],因此实验设置了3.0%、3.2%和3.4% 3 个钙水平,以验证条件是否满足。差量法计算的对照组内源钙排泄量为2.28~2.30g/kg DMΙ,而GlcΝ组的内源钙排泄量为2.29~2.30 g/kg DMΙ。方差分析显示:钙水平和GlcΝ 对内源钙排泄量均无显著影响。用回归法得到的对照各组和GlcΝ 各组内源钙排泄量分别为2.29 g/kg DMΙ 和2.30 g/kg DMΙ,与差量法计算得到的值相吻合,亦与《家禽营养》中蛋鸡每天内源钙排泄量为0.1~0.28 g(按每只鸡每天采食100 g 可换算为0.83~2.33 g/kg DMΙ)[18]及胡海波等研究肉鸡内源钙排泄量为1.94~2.3 g/kg DMΙ[17,19]的结果一致。本研究结果表明钙水平在3.0%~3.4%时,蛋鸡内源钙排泄稳定,满足差量法和回归法测定饲粮钙真代谢率的前提条件。
3.2 GlcΝ 对蛋鸡钙吸收的调节 饲粮中钙的消化吸收受粒径大小[19-21]和钙源[21-22]的影响。肉鸡对石灰石粉的表观回肠消化率在54%~61%之间[23]。粒径小于0.5 mm的石粉钙的回肠真消化率为35%~54%,而粒径1~2 mm的大颗粒石灰石中钙的真消化率则可达63%~77%[19-21]。牡蛎壳中钙的真消化率则为34%~56%[17],磷酸氢钙中钙的真消化率为30%~34%。故在实验中各组饲粮的颗粒钙与粉钙提供钙的比例保持1:1,磷酸氢钙添加比例亦在各组间一致,以排除钙源和颗粒大小对钙代谢率测定结果的干扰。本实验蛋鸡对饲粮中钙的表观代谢率为50.75%~51.69%,真代谢率约为57.80%,与上述报道[23]相近。在饲粮中添加0.6%GlcΝ 钙的表观和真代谢率分别为62.77% 和真代谢率为69.3%,表明添加0.6%GlcΝ 可显著提高蛋鸡对饲粮钙的吸收。但GlcΝ 是如何促进蛋鸡肠道对钙的吸收,鲜有报道。本实验的RTPCR 分析结果显示添加0.6% GlcΝ 显著上调了蛋鸡十二指肠Cabp-D28k基因表达,且Cabp-D28k基因表达与饲粮钙的表观代谢率和真代谢率呈极显著正相关,和报道的Cabp-D28k表达量与肠钙吸收呈正相关相吻合[10-13],提示GlcΝ 可通过促进十二指肠Cabp-D28k基因表达来增强蛋鸡对饲粮钙的吸收。但GlcΝ 是通过何种途径上调Cabp-D28k表达,则有待借助分子生物学技术阐释其分子机制。此外,有研究表明GlcΝ 的ΝH2+与OH-使其具有与多价金属螯合的能力[24-27],而螯合作用可促进机体对金属元素的吸收,提高其利用率[28-30],GlcΝ 提高钙吸收是否与其螯合特性有关,亦有待于进一步探讨。
4 结论
1)在饲粮中添加0.6% GlcΝ 可分别提高饲粮钙的表观代谢率(11.54%)和真代谢率(11.42%)。
2)GlcΝ 可通过上调Cabp-D28k基因表达促进蛋鸡对饲粮钙的吸收。
猜你喜欢
农业工程学报(2022年8期)2022-08-08
作物学报(2022年9期)2022-07-18
知识窗(2022年6期)2022-07-08
中老年保健(2021年10期)2021-11-30
晚晴(2021年1期)2021-05-17
福建基础教育研究(2019年10期)2019-05-28
科学之谜(2018年10期)2018-01-02
安徽理工大学学报·社会科学版(2014年6期)2015-02-06
银行家(2014年1期)2014-02-14
